
Imported SARS-CoV-2 Variants of Concern Drove Spread of Infections across Kenya during the Second Year of the Pandemic

 , , ,
, , ,  , ,
, ,  and add
Show full author list
and add
Show full author list
Abstract
:1. Introduction
2. Materials and Methods
2.1. COVID-19 Surveillance Data
2.2. SARS-CoV-2 Testing and Reporting
2.3. SARS-CoV-2 Testing Inequity
2.4. Collection of Fatality Data
2.5. Epidemiology Data Analysis
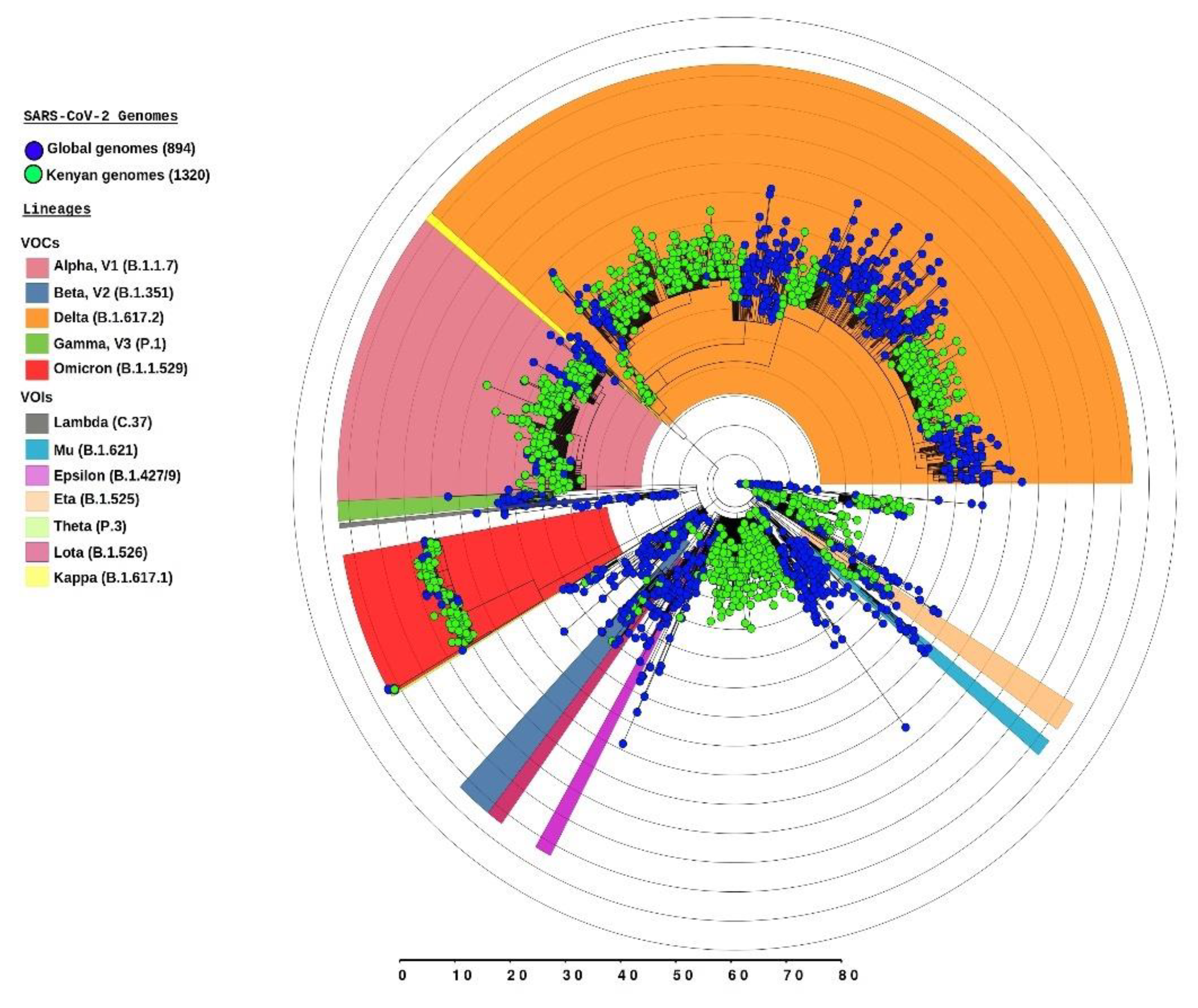
2.6. SARS-CoV-2 Genomic Surveillance
2.7. Whole Genome Sequencing and Variant Identification
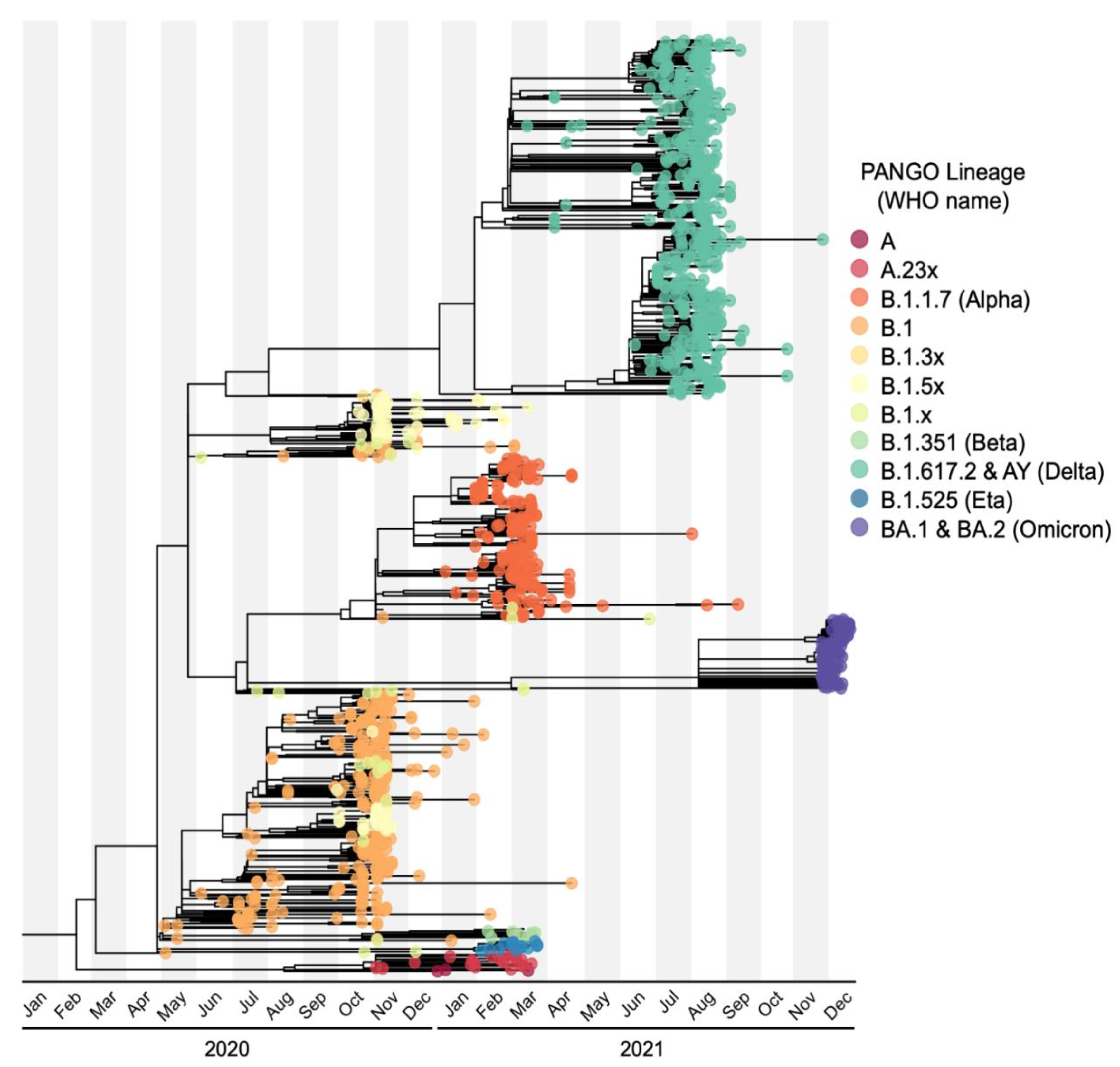
2.8. Phylogenetic Analyses
3. Results
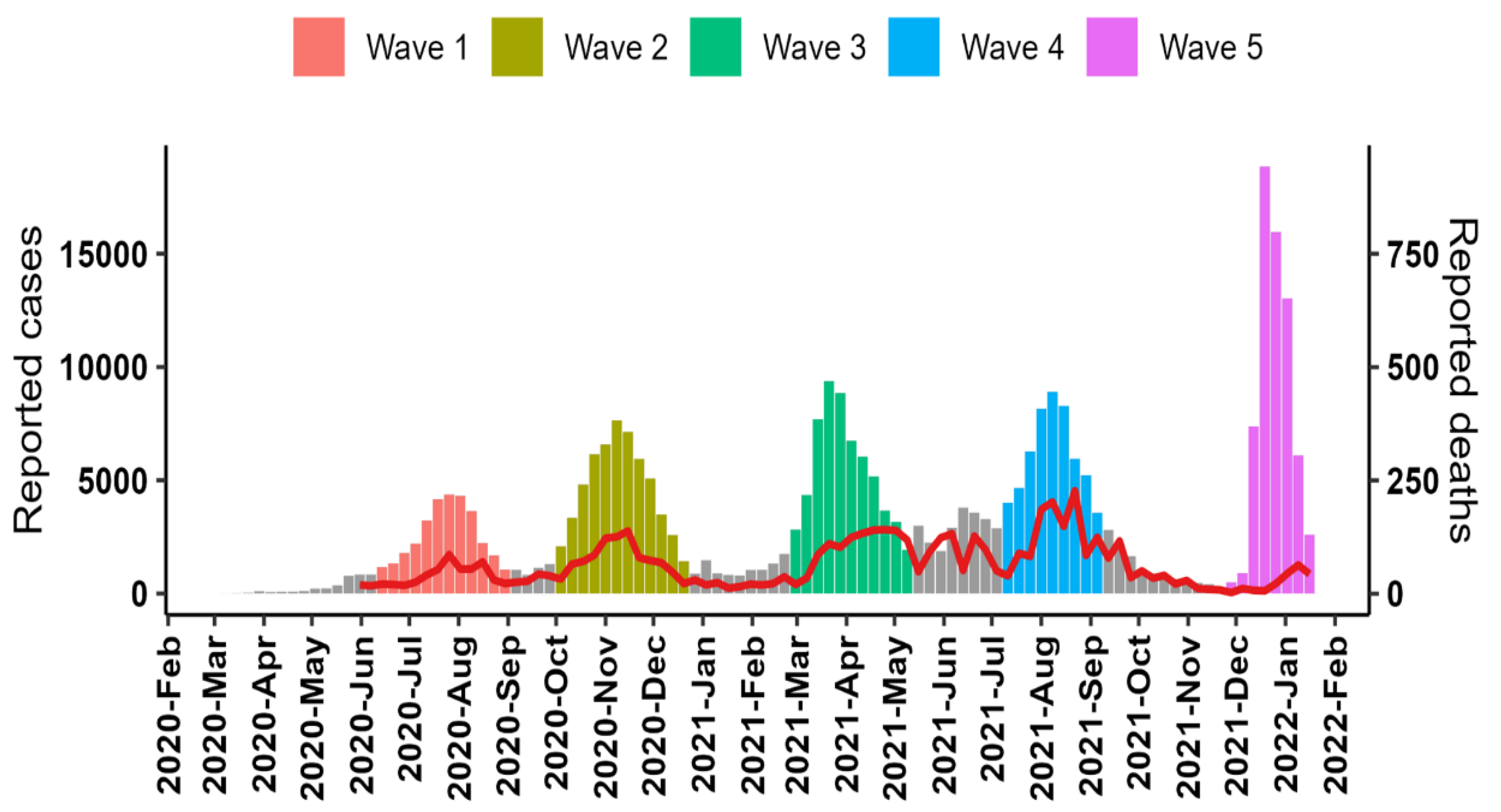
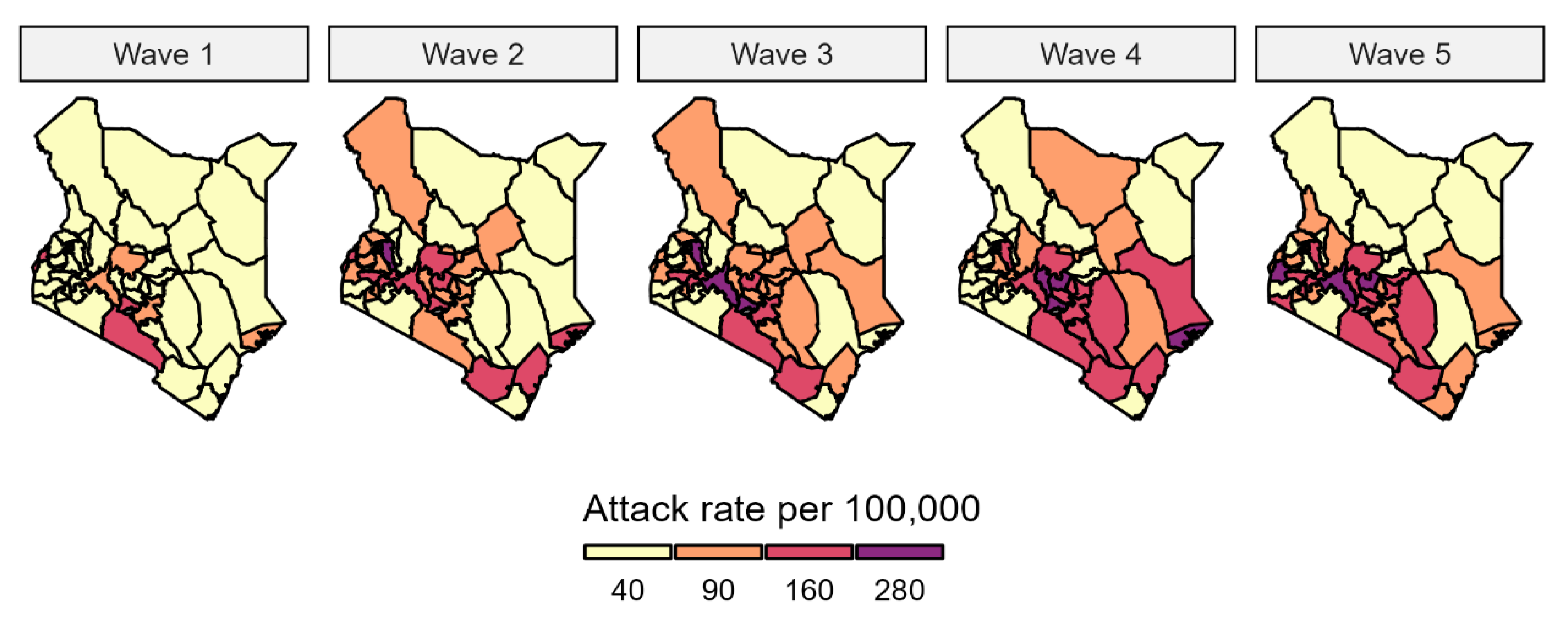
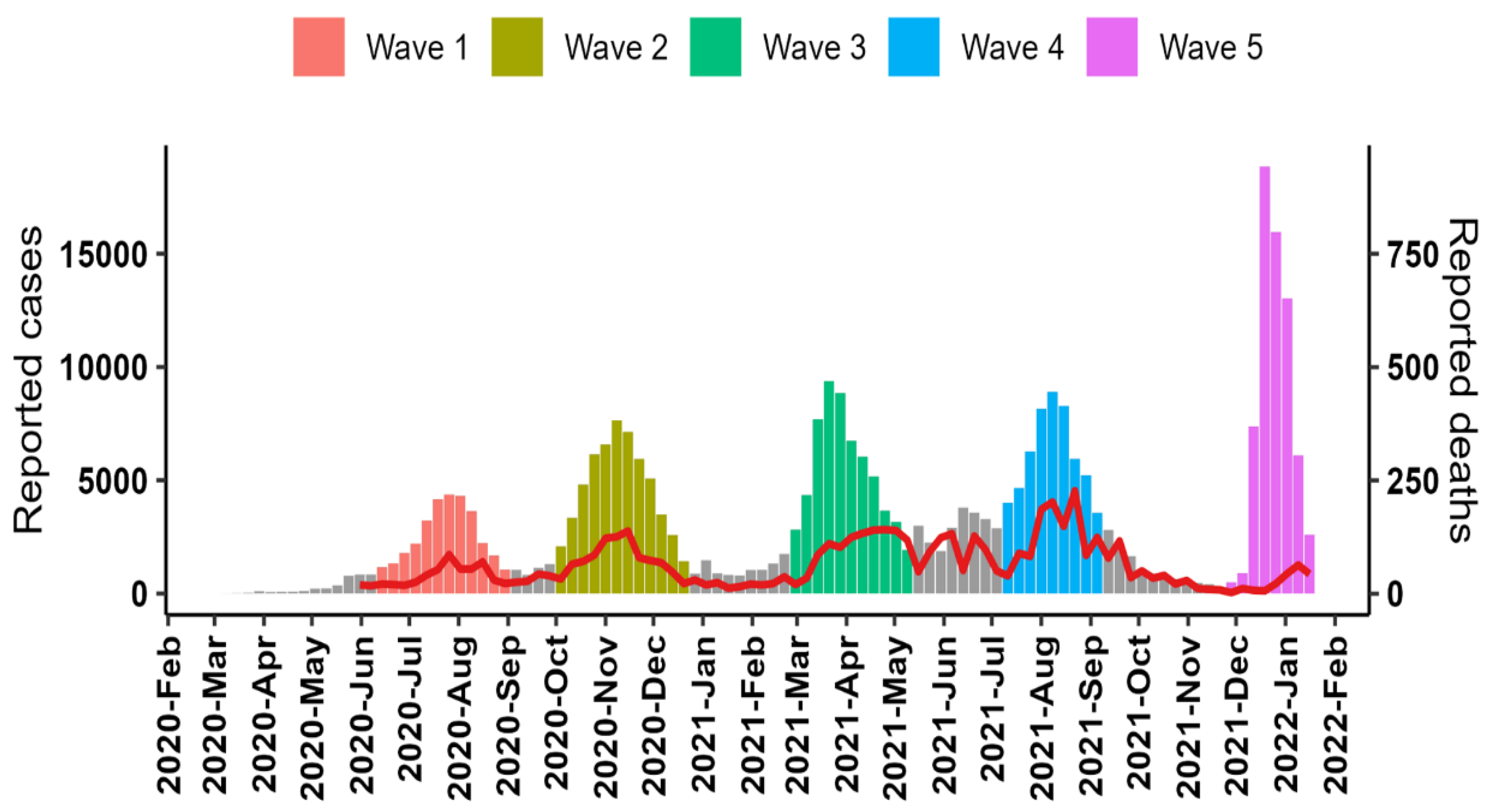
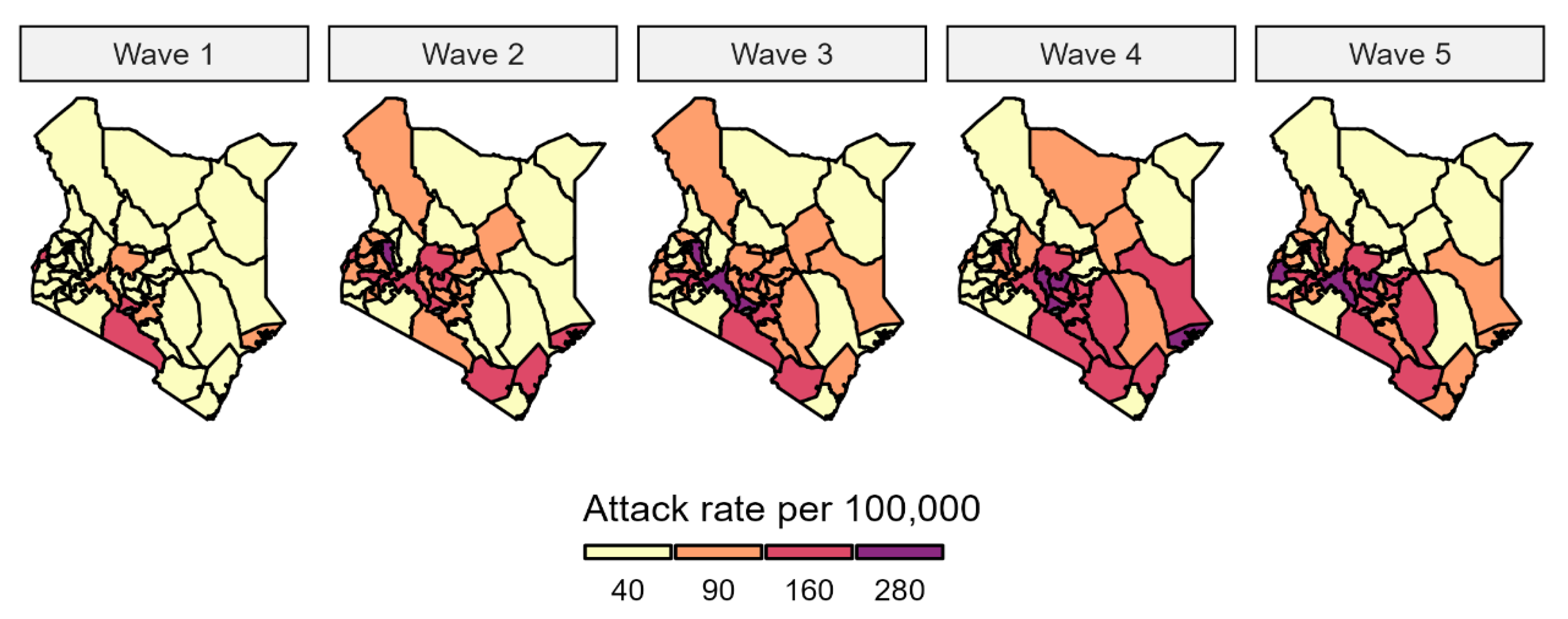
3.1. Pandemic Waves and Regional Spread of Infections
3.2. Case Fatality Ratio
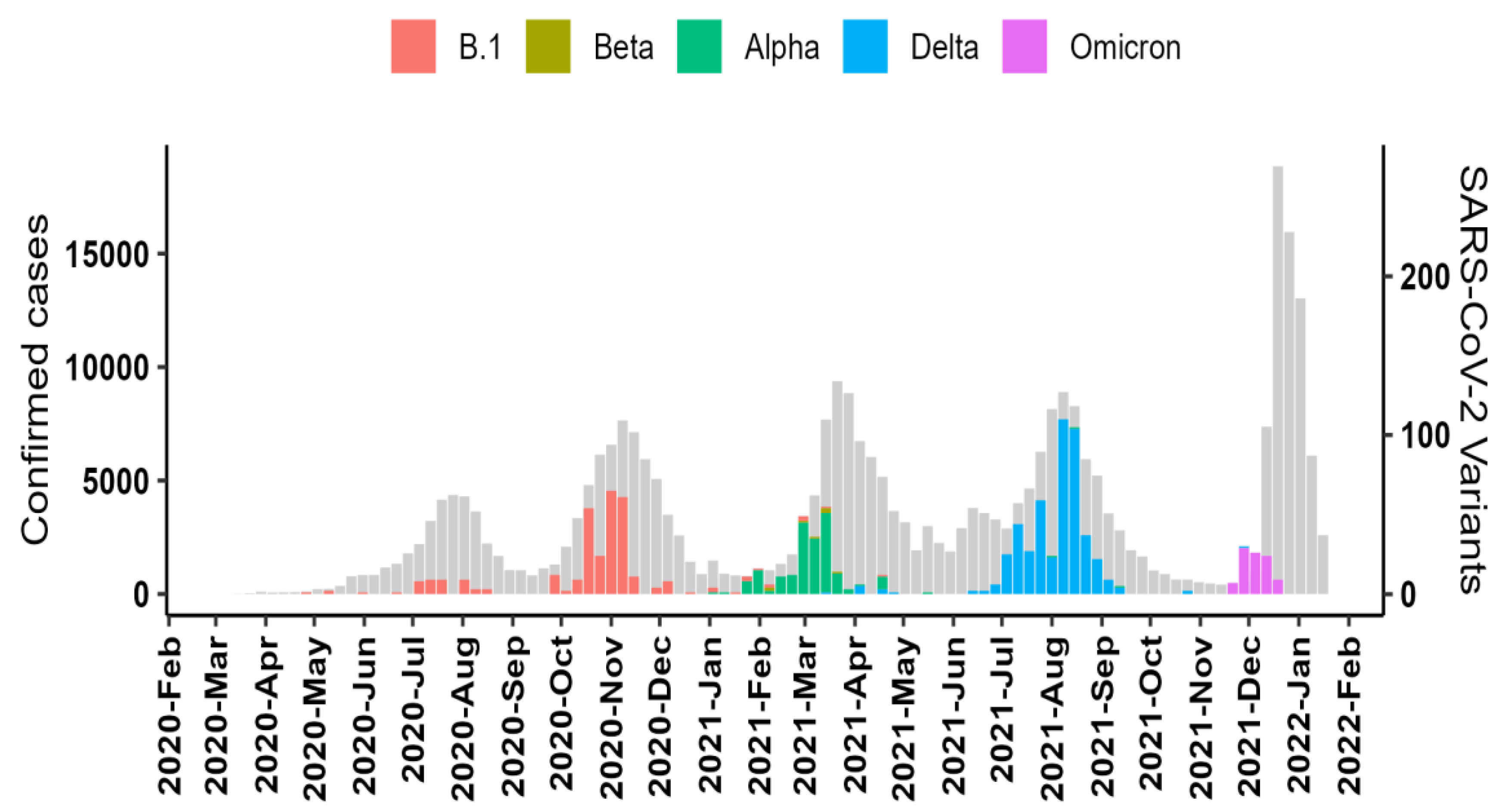
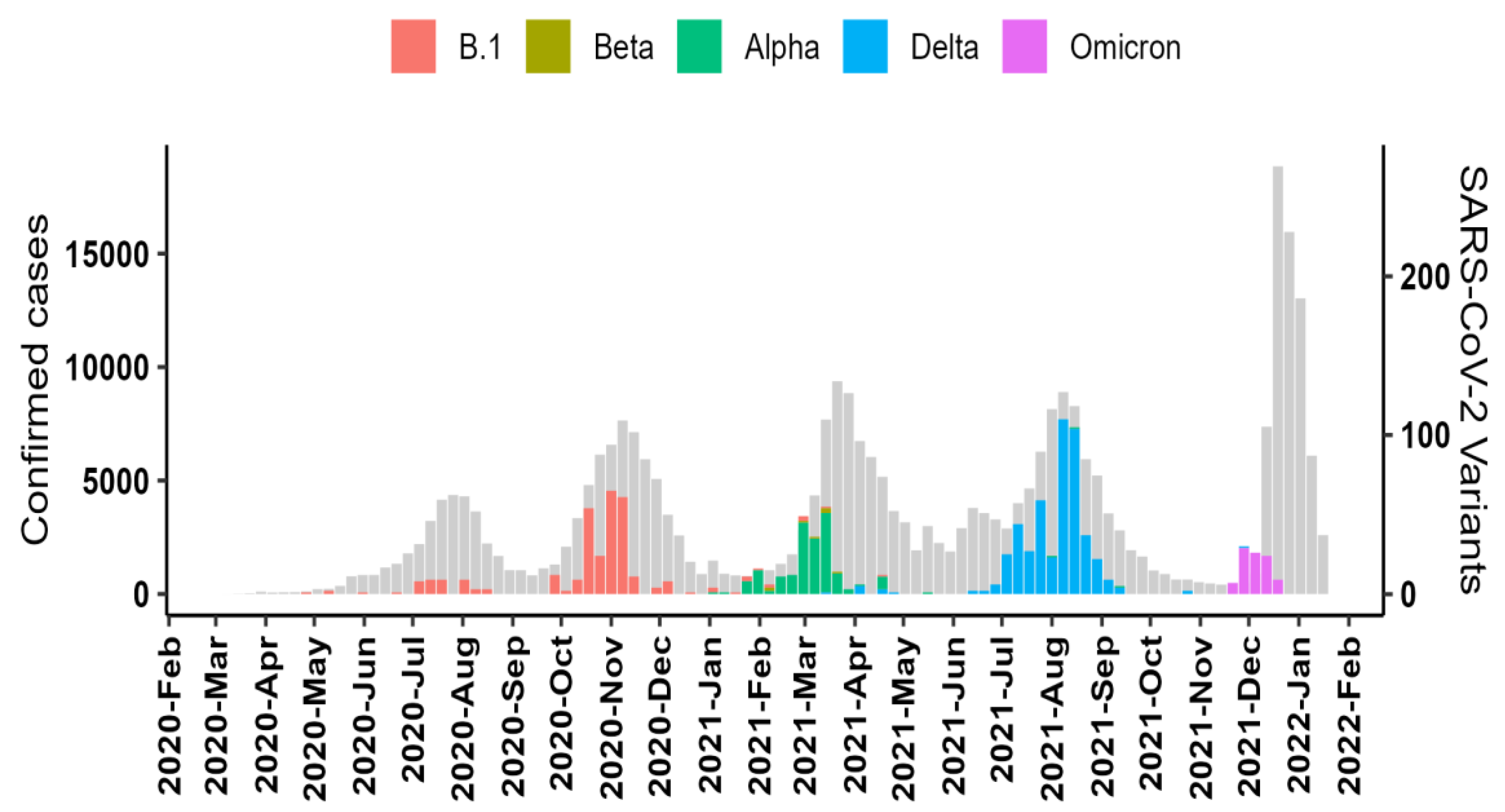
3.3. Dominant SARS-CoV-2 Lineages during Waves
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salyer, S.J.; Maeda, J.; Sembuche, S.; Kebede, Y.; Tshangela, A.; Moussif, M.; Ihekweazu, C.; Mayet, N.; Abate, E.; Ouma, A.O.; et al. The first and second waves of the COVID-19 pandemic in Africa: A cross-sectional study. Lancet 2021, 397, 1265–1275. [Google Scholar] [CrossRef]
- Hale, T.; Angrist, N.; Hale, A.J.; Kira, B.; Majumdar, S.; Petherick, A.; Phillips, T.; Sridhar, D.; Thompson, R.N.; Webster, S.; et al. Government responses and COVID-19 deaths: Global evidence across multiple pandemic waves. PLoS ONE 2021, 16, e0253116. [Google Scholar] [CrossRef]
- Jassat, W.; Mudara, C.; Ozougwu, L.; Tempia, S.; Blumberg, L.; Davies, M.-A.; Pillay, Y.; Carter, T.; Morewane, R.; Wolmarans, M.; et al. Difference in mortality among individuals admitted to hospital with COVID-19 during the first and second waves in South Africa: A cohort study. Lancet Glob. Health 2021, 9, e1216–e1225. [Google Scholar] [CrossRef]
- Aleem, A.; Akbar Samad, A.B.; Slenker, A.K. Emerging Variants of SARS-CoV-2 and Novel Therapeutics against Coronavirus (COVID-19); StatPearls Publishing: Treasure Island, FL, USA, 2022. Available online: http://www.ncbi.nlm.nih.gov/books/NBK570580/ (accessed on 27 January 2022).
- Konings, F.; Perkins, M.D.; Kuhn, J.H.; Pallen, M.J.; Alm, E.J.; Archer, B.N.; Barakat, A.; Bedford, T.; Bhiman, J.N.; Caly, L.; et al. SARS-CoV-2 Variants of Interest and Concern naming scheme conducive for global discourse. Nat. Microbiol. 2021, 6, 821–823. [Google Scholar] [CrossRef] [PubMed]
- Cantón, R.; Ramos, P.D.L.; García-Botella, A.; García-Lledó, A.; Gómez-Pavón, J.; Del Castillo, J.G.; Hernández-Sampelayo, T.; Martín-Delgado, M.C.; Martín Sánchez, F.J.; Martínez-Sellés, M.; et al. New variants of SARS-CoV-2. Rev. Esp. Quimioter. 2021, 34, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Njenga, M.K.; Dawa, J.; Nanyingi, M.; Gachohi, J.; Ngere, I.; Letko, M.; Otieno, C.F.; Gunn, B.M.; Osoro, E. Why is There Low Morbidity and Mortality of COVID-19 in Africa? Am. J. Trop. Med. Hyg. 2020, 103, 564–569. [Google Scholar] [CrossRef]
- Ngere, I.; Dawa, J.; Hunsperger, E.; Otieno, N.; Masika, M.; Amoth, P.; Makayotto, L.; Nasimiyu, C.; Gunn, B.M.; Nyawanda, B.; et al. High seroprevalence of SARS-CoV-2 but low infection fatality ratio eight months after introduction in Nairobi, Kenya. Int. J. Infect. Dis. 2021, 112, 25–34. [Google Scholar] [CrossRef]
- McKibbin, W.; Fernando, R. The Global Macroeconomic Impacts of COVID-19: Seven Scenarios. Asian Econ. Pap. 2021, 20, 1–30. [Google Scholar] [CrossRef]
- Tegally, H.; Wilkinson, E.; Giovanetti, M.; Iranzadeh, A.; Fonseca, V.; Giandhari, J.; Doolabh, D.; Pillay, S.; James San, E.; Msomi, N.; et al. Detection of a SARS-CoV-2 variant of concern in South Africa. Nature 2021, 592, 438–443. [Google Scholar] [CrossRef]
- Novelli, G.; Colona, V.L.; Pandolfi, P.P. A focus on the spread of the delta variant of SARS-CoV-2 in India. Indian J. Med. Res. 2021, 153, 537–541. [Google Scholar] [CrossRef]
- da Silva, J.F.; Esteves, R.J.; Siza, C.; Soares, E.P.; Ramos, T.C.; Campelo, E.C.; da Costa, C.F.; de Alencar, L.C.; Cavalcante, R.P.; Florêncio, C.R.; et al. Cluster of SARS-CoV-2 Gamma Variant Infections, Parintins, Brazil, March 2021. Emerg. Infect. Dis. 2022, 28, 262–264. [Google Scholar] [CrossRef] [PubMed]
- Jansen, L. Investigation of a SARS-CoV-2 B.1.1.529 (Omicron) Variant Cluster—Nebraska, November–December 2021. MMWR Morb. Mortal. Wkly. Rep. 2021, 70, 1782–1784. Available online: https://www.cdc.gov/mmwr/volumes/70/wr/mm705152e3.htm (accessed on 4 February 2022). [CrossRef] [PubMed]
- Yang, S.; Hemarajata, P.; Hilt, E.E.; Price, T.K.; Garner, O.B.; Green, N.M. Investigation of SARS-CoV-2 Epsilon Variant and Hospitalization Status by Genomic Surveillance in a Single Large Health System during the 2020–2021 Winter Surge in Southern California. Am. J. Clin. Pathol. 2021, aqab203. [Google Scholar] [CrossRef] [PubMed]
- Wangari, E.N.; Gichuki, P.; Abuor, A.A.; Wambui, J.; Okeyo, S.O.; Oyatsi, H.T.N.; Odikara, S.; Kulohoma, B.W. Kenya’s response to the COVID-19 pandemic: A balance between minimising morbidity and adverse economic impact. AAS Open Res. 2021, 4, 3. [Google Scholar] [CrossRef] [PubMed]
- Brand, S.P.C.; Ojal, J.; Aziza, R.; Were, V.; Okiro, E.A.; Kombe, I.K.; Mburu, C.; Ogero, M.; Agweyu, A.; Warimwe, G.M.; et al. COVID-19 transmission dynamics underlying epidemic waves in Kenya. Science 2021, 374, 989–994. Available online: https://www.science.org/doi/abs/10.1126/science.abk0414 (accessed on 25 December 2021). [CrossRef]
- Ministry of Health—Republic of Kenya. Available online: https://www.health.go.ke/ (accessed on 9 June 2021).
- Orangi, S.; Pinchoff, J.; Mwanga, D.; Abuya, T.; Hamaluba, M.; Warimwe, G.; Austrian, K.; Barasa, E. Assessing the level and determinants of COVID-19 Vaccine Confidence in Kenya. Vaccines 2021, 9, 936. Available online: https://www.medrxiv.org/content/10.1101/2021.06.11.21258775v1 (accessed on 4 December 2020). [CrossRef]
- COVID-19 Vaccination: Campaign Targeting Public Transport Operators Launched—Ministry of Health. Available online: https://www.health.go.ke/covid-19-vaccination-campaign-targeting-public-transport-operators-launched/ (accessed on 9 January 2022).
- Kenya Increases Uptake and Equity for COVID-19 Vaccinations. Available online: https://www.who.int/news-room/feature-stories/detail/kenya-increases-uptake-and-equity-for-covid-19-vaccinations (accessed on 7 January 2022).
- Gohl, D.M.; Garbe, J.; Grady, P.; Daniel, J.; Watson, R.H.B.; Auch, B.; Nelson, A.; Yohe, S.; Beckman, K.B. A rapid, cost-effective tailed amplicon method for sequencing SARS-CoV-2. BMC Genom. 2020, 21, 863. [Google Scholar] [CrossRef]
- Grubaugh, N.D.; Gangavarapu, K.; Quick, J.; Matteson, N.L.; De Jesus, J.G.; Main, B.J.; Tan, A.L.; Paul, L.M.; Brackney, D.E.; Grewal, S.; et al. An amplicon-based sequencing framework for accurately measuring intrahost virus diversity using PrimalSeq and iVar. Genome Biol. 2019, 20, 8. [Google Scholar] [CrossRef] [Green Version]
- Releases nf-Core/Viralrecon. GitHub. Available online: https://github.com/nf-core/viralrecon/releases (accessed on 28 January 2022).
- Hadfield, J.; Megill, C.; Bell, S.M.; Huddleston, J.; Potter, B.; Callender, C.; Sagulenko, P.; Bedford, T.; Neher, R.A. Nextstrain: Real-time tracking of pathogen evolution. Bioinformatics 2018, 34, 4121–4123. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- FigTree. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 28 January 2022).
- Didelot, X.; Wilson, D.J. ClonalFrameML: Efficient Inference of Recombination in Whole Bacterial Genomes. PLoS Comput. Biol. 2015, 11, e1004041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Toole, Á.; Scher, E.; Underwood, A.; Jackson, B.; Hill, V.; McCrone, J.T.; Colquhoun, R.; Ruis, C.; Abu-Dahab, K.; Taylor, B.; et al. Assignment of epidemiological lineages in an emerging pandemic using the pangolin tool. Virus Evol. 2021, 7, veab064. [Google Scholar] [CrossRef] [PubMed]
- Aksamentov, I.; Roemer, C.; Hodcroft, E.; Neher, R. Nextclade: Clade assignment, mutation calling and quality control for viral genomes. JOSS 2021, 6, 3773. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Haeseler, A.; von Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Hoang, D.T.; Chernomor, O.; Haeseler, A.; von Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Sagulenko, P.; Puller, V.; Neher, R.A. TreeTime: Maximum-likelihood phylodynamic analysis. Virus Evol. 2018, 4, vex042. [Google Scholar] [CrossRef]
- Chia, W.N.; Zhu, F.; Ong, S.W.X.; Young, B.E.; Fong, S.W.; Bert, N.L.; Tan, C.W.; Tiu, C.; Zhang, J.; Tan, S.Y.; et al. Dynamics of SARS-CoV-2 neutralising antibody responses and duration of immunity: A longitudinal study. Lancet Microbe. 2021, 2, e240–e249. [Google Scholar] [CrossRef]
- Harvey, W.T.; Carabelli, A.M.; Jackson, B.; Gupta, R.K.; Thomson, E.C.; Harrison, E.M.; Ludden, C.; Reeve, R.; Rambaut, A.; Robertson, D.R.; et al. SARS-CoV-2 variants, spike mutations and immune escape. Nat. Rev. Microbiol. 2021, 19, 409–424. [Google Scholar] [CrossRef]
- Xu, X.; Nie, S.; Wang, Y.; Long, Q.; Zhu, H.; Zhang, X.; Sun, J.; Zeng, Q.; Zhao, J.; Liu, L.; et al. Dynamics of neutralizing antibody responses to SARS-CoV-2 in patients with COVID-19: An observational study. Signal Transduct. Target Ther. 2021, 6, 197. [Google Scholar] [CrossRef]
- Anand, S.P.; Prévost, J.; Nayrac, M.; Beaudoin-Bussières, G.; Benlarbi, M.; Gasser, R.; Brassard, N.; Laumaea, A.; Gong, S.Y.; Bourassa, C.; et al. Longitudinal analysis of humoral immunity against SARS-CoV-2 Spike in convalescent individuals up to 8 months post-symptom onset. Cell Rep. Med. 2021, 2, 100290. [Google Scholar] [CrossRef]
- Li, B.; Deng, A.; Li, K.; Hu, Y.; Li, Z.; Shi, Y.; Xiong, Q.; Liu, Z.; Guo, Q.; Zou, L.; et al. Viral infection and transmission in a large, well-traced outbreak caused by the SARS-CoV-2 Delta variant. Nat. Commun. 2022, 13, 460. [Google Scholar] [CrossRef] [PubMed]
- Hirabara, S.M.; Serdan, T.D.A.; Gorjao, R.; Masi, L.N.; Pithon-Curi, T.C.; Covas, D.T.; Curi, R.; Durigon, E.L. SARS-CoV-2 Variants: Differences and Potential of Immune Evasion. Front. Cell. Infect. Microbiol. 2022, 11, 781429. Available online: https://www.frontiersin.org/article/10.3389/fcimb.2021.781429 (accessed on 23 April 2022). [CrossRef] [PubMed]
- Thomson, E.C.; Rosen, L.E.; Shepherd, J.G.; Spreafico, R.; da Silva Filipe, A.; Wojcechowskyj, J.A.; Davis, C.; Piccoli, L.; Pascall, D.J.; Dillen, J.; et al. Circulating SARS-CoV-2 spike N439K variants maintain fitness while evading antibody-mediated immunity. Cell 2021, 184, 1171–1187.e20. [Google Scholar] [CrossRef] [PubMed]
- López, M.; Peinado, A.; Ortiz, A. Characterizing two outbreak waves of COVID-19 in Spain using phenomenological epidemic modelling. PLoS ONE 2021, 16, e0253004. [Google Scholar] [CrossRef]
- Salfi, F.; D’Atri, A.; Tempesta, D.; Ferrara, M. Sleeping under the waves: A longitudinal study across the contagion peaks of the COVID-19 pandemic in Italy. J. Sleep Res. 2021, 30, e13313. [Google Scholar] [CrossRef]
- Kimita, G.; Nyataya, J.; Omuseni, E.; Sigei, F.; Lemtudo, A.; Muthanje, E.; Andika, B.; Liyai, R.; Githii, R.; Masakhwe, C.; et al. A genomics dissection of Kenya’s COVID-19 waves: Temporal lineage replacements and dominance of imported variants of concer. Res. Sq. 2021. Available online: https://europepmc.org/article/PPR/PPR408814 (accessed on 28 January 2022).
- Githinji, G.; Laurent, Z.R.; de Mohammed, K.S.; Omuoyo, D.O.; Macharia, P.M.; Morobe, J.M.; Otieno, E.; Kinyanjui, S.M.; Agweyu, A.; Maitha, E.; et al. Tracking the introduction and spread of SARS-CoV-2 in coastal Kenya. Nat. Commun. 2021, 12, 4809. [Google Scholar] [CrossRef]
- Grabowski, F.; Kochańczyk, M.; Lipniacki, T. Omicron strain spreads with the doubling time of 3.2–3.6 days in South Africa province of Gauteng that achieved herd immunity to Delta variant. medRxiv 2021. Available online: https://www.medrxiv.org/content/10.1101/2021.12.08.21267494v1 (accessed on 28 January 2022).
- Xia, F.; Yang, X.; Cheke, R.A.; Xiao, Y. Quantifying competitive advantages of mutant strains in a population involving importation and mass vaccination rollout. Infect. Dis. Model. 2021, 6, 988–996. [Google Scholar] [CrossRef]
- Russell, A.; O’Connor, C.; Lasek-Nesselquist, E.; Plitnick, J.; Kelly, J.P.; Lamson, D.M.; George, K.S. Spatiotemporal analyses illuminate the competitive advantage of a SARS-CoV-2 variant of concern over a variant of interest. medRxiv 2021. Available online: https://www.medrxiv.org/content/10.1101/2021.09.14.21262977v1 (accessed on 14 September 2021).
- Garcia-Beltran, W.F.; Lam, E.C.; St Denis, K.; Nitido, A.D.; Garcia, Z.H.; Hauser, B.M.; Feldman, J.; Pavlovic, M.N.; Gregory, D.J.; Poznansky, M.C.; et al. Multiple SARS-CoV-2 variants escape neutralization by vaccine-induced humoral immunity. Cell 2021, 184, 2372–2383.e9. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
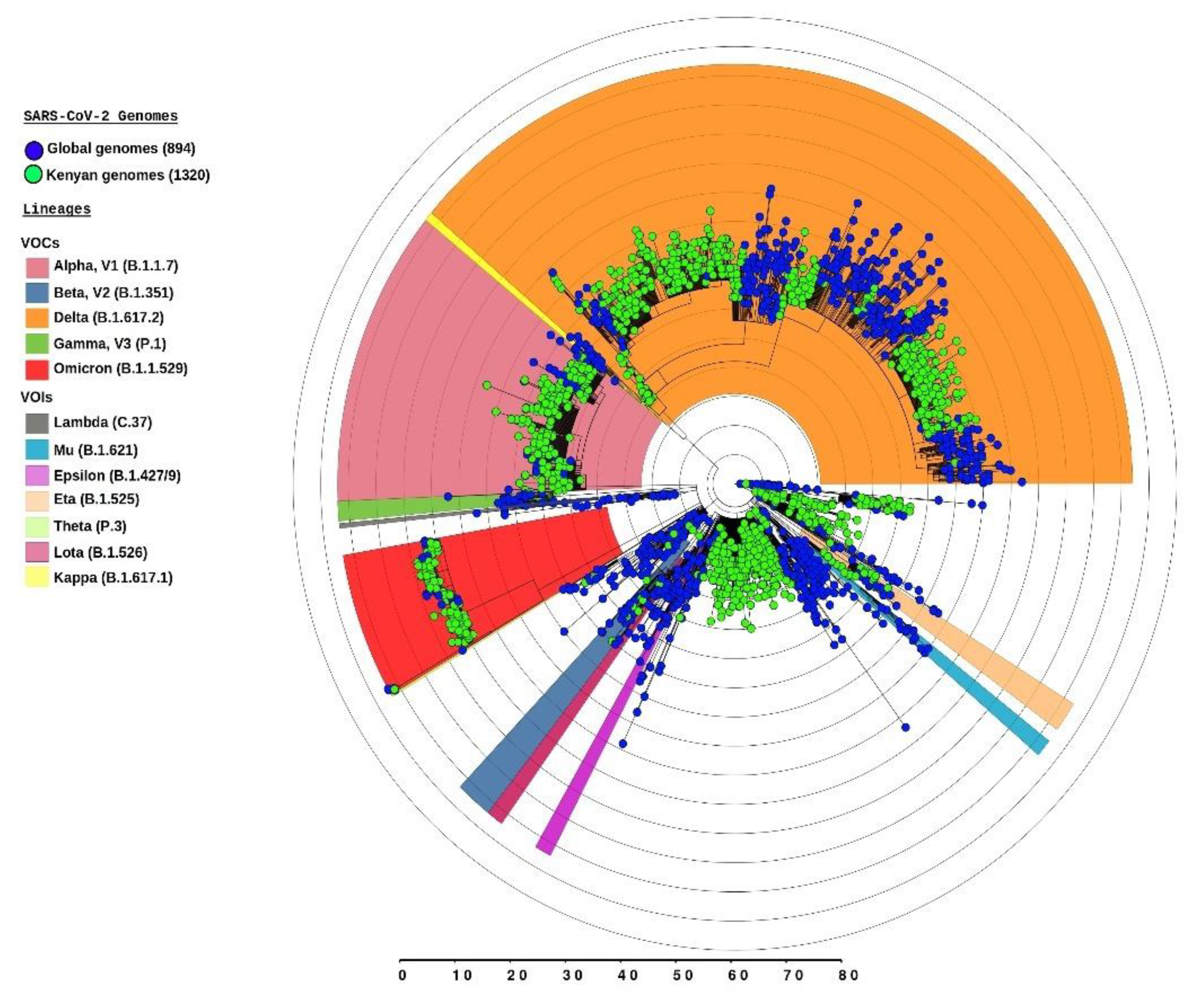{kind=link}
| All Cases | All Deaths | Female Cases | Male Cases | Male Deaths | Female Deaths | |||
|---|---|---|---|---|---|---|---|---|
| Age Group | n | % (95% CI) | n | % (95% CI) | % (95% CI) | % (95% CI) | % (95% CI) | % (95% CI) |
| 0–9 | 12,064 | 3.8 (3.7–3.8) | 89 | 1.6 (1.3–2) | 43.7 (42.8–44.6) | 56.3 (55.4–57.2) | 65.2 (54.3–74.8) | 34.8 (25.2–45.7) |
| 10–19 | 19,832 | 6.2 (6.1–6.3) | 72 | 1.3 (1–1.6) | 47.6 (46.9–48.3) | 52.4 (51.7–53.1) | 58.3 (46.1–69.6) | 41.7 (30.4–53.9) |
| 20–29 | 60,860 | 19 (18.9–19.1) | 162 | 2.9 (2.5–3.4) | 47.3 (46.9–47.7) | 52.7 (52.3–53.1) | 51.9 (43.9–59.7) | 48.1 (40.3–56.1) |
| 30–39 | 83,067 | 25.9 (25.8–26.1) | 428 | 7.7 (7–8.4) | 43.1 (42.8–43.5) | 56.9 (56.5–57.2) | 51.9 (47–56.7) | 48.1 (43.3–53) |
| 40–49 | 59,099 | 18.5 (18.3–18.6) | 683 | 12.2 (11.4–13.1) | 40.1 (39.7–40.5) | 59.9 (59.5–60.3) | 63.5 (59.8–67.1) | 36.5 (32.9–40.2) |
| 50–59 | 41,394 | 12.9 (12.8–13) | 996 | 17.8 (16.9–18.9) | 41.5 (41.1–42) | 58.5 (58–58.9) | 68.4 (65.4–71.2) | 31.6 (28.8–34.6) |
| ≥60 | 43,998 | 13.7 (13.6–13.9) | 3152 | 56.5 (55.2–57.8) | 44.2 (43.8–44.7) | 55.8 (55.3–56.2) | 64.6 (62.9–66.3) | 35.4 (33.7–37.1) |
| Total | 320,314 | - | 5582 | - | 43.6 (43.4–43.8) | 56.4 (56.2–56.6) | 63.7 (62.5–65) | 36.3 (35–37.5) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nasimiyu, C.; Matoke-Muhia, D.; Rono, G.K.; Osoro, E.; Ouso, D.O.; Mwangi, J.M.; Mwikwabe, N.; Thiong’o, K.; Dawa, J.; Ngere, I.; et al. Imported SARS-CoV-2 Variants of Concern Drove Spread of Infections across Kenya during the Second Year of the Pandemic. COVID 2022, 2, 586-598. https://doi.org/10.3390/covid2050044
Nasimiyu C, Matoke-Muhia D, Rono GK, Osoro E, Ouso DO, Mwangi JM, Mwikwabe N, Thiong’o K, Dawa J, Ngere I, et al. Imported SARS-CoV-2 Variants of Concern Drove Spread of Infections across Kenya during the Second Year of the Pandemic. COVID. 2022; 2(5):586-598. https://doi.org/10.3390/covid2050044
Chicago/Turabian StyleNasimiyu, Carolyne, Damaris Matoke-Muhia, Gilbert K. Rono, Eric Osoro, Daniel O. Ouso, J. Milkah Mwangi, Nicholas Mwikwabe, Kelvin Thiong’o, Jeanette Dawa, Isaac Ngere, and et al. 2022. "Imported SARS-CoV-2 Variants of Concern Drove Spread of Infections across Kenya during the Second Year of the Pandemic" COVID 2, no. 5: 586-598. https://doi.org/10.3390/covid2050044
APA StyleNasimiyu, C., Matoke-Muhia, D., Rono, G. K., Osoro, E., Ouso, D. O., Mwangi, J. M., Mwikwabe, N., Thiong’o, K., Dawa, J., Ngere, I., Gachohi, J., Kariuki, S., Amukoye, E., Mureithi, M., Ngere, P., Amoth, P., Were, I., Makayotto, L., Nene, V., ... Oyola, S. O. (2022). Imported SARS-CoV-2 Variants of Concern Drove Spread of Infections across Kenya during the Second Year of the Pandemic. COVID, 2(5), 586-598. https://doi.org/10.3390/covid2050044