
SARS-CoV-2 Targets and COVID-19 Vaccines

Abstract
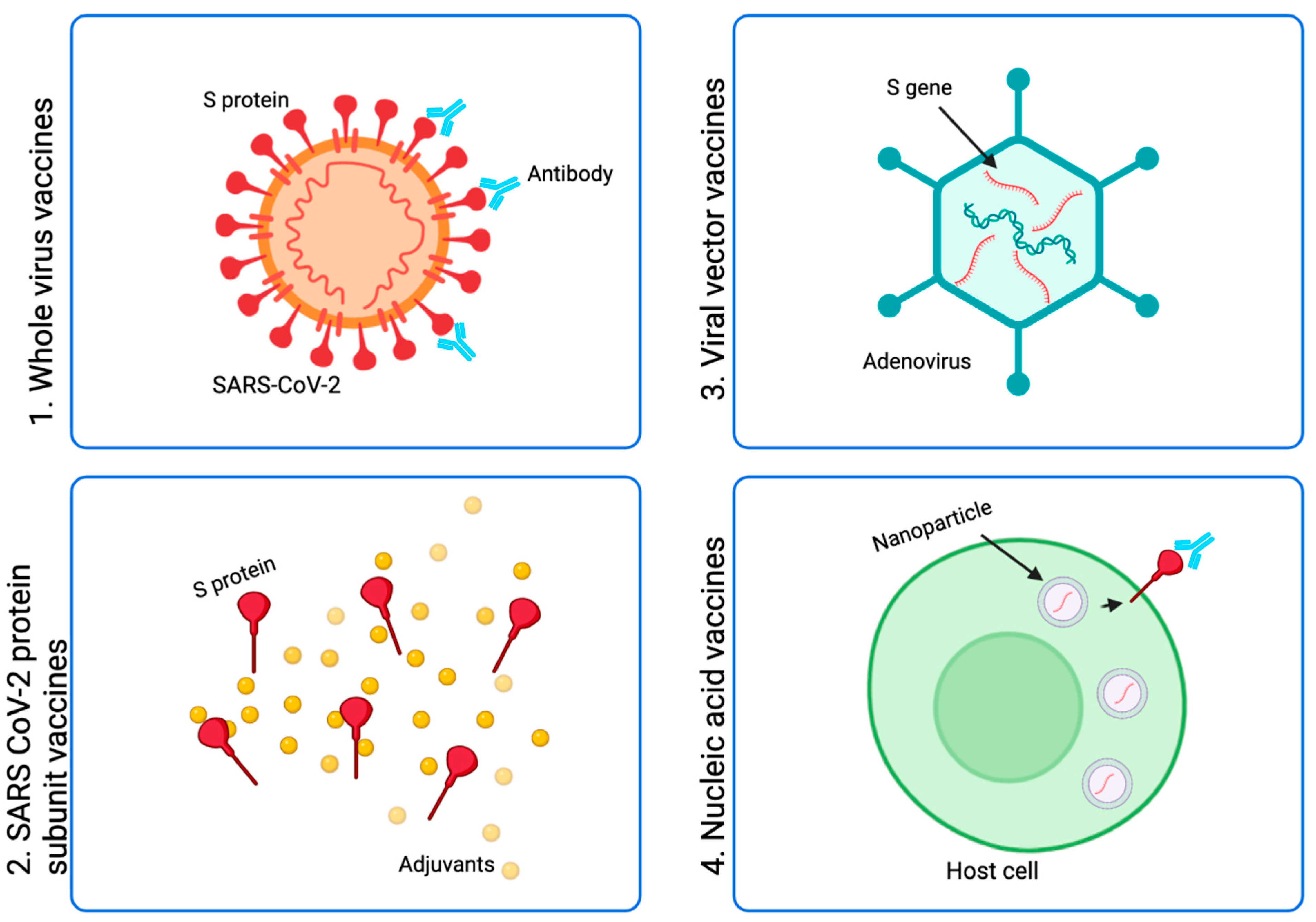
:1. Introduction
1.1. Whole SARS-CoV-2 Virus Vaccines
1.2. SARS-CoV-2 Protein Subunit Vaccines
1.3. Viral Vector Vaccines
1.4. Nucleic Acid Vaccines
2. Efficacy of COVID-19 Vaccines
3. SARS-CoV-2 Variants
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- D’Cruz, R.J.; Currier, A.W.; Sampson, V.B. Laboratory Testing Methods for Novel Severe Acute Respiratory Syndrome-Coronavirus-2 (SARS-CoV-2). Front. Cell Dev. Biol. 2020, 8, 468. [Google Scholar] [CrossRef]
- WHO. Naming the Coronavirus Disease (COVID-19) and the Virus that Causes It. 2020. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019/technical-guidance/naming-the-coronavirus-disease-(covid-2019)-and-the-virus-that-causes-it (accessed on 9 September 2021).
- Shrotri, M.; Swinnen, T.; Kampmann, B.; Parker, E.P.K. An interactive website tracking COVID-19 vaccine development. Lancet Glob. Health 2021, 9, e590–e592. [Google Scholar] [CrossRef]
- Xia, S.; Duan, K.; Zhang, Y.; Zhao, D.; Zhang, H.; Xie, Z.; Li, X.; Peng, C.; Zhang, Y.; Zhang, W.; et al. Effect of an Inactivated Vaccine Against SARS-CoV-2 on Safety and Immunogenicity Outcomes: Interim Analysis of 2 Randomized Clinical Trials. JAMA 2020, 324, 951–960. [Google Scholar] [CrossRef] [PubMed]
- Moller, L.; Schunadel, L.; Nitsche, A.; Schwebke, I.; Hanisch, M.; Laue, M. Evaluation of virus inactivation by formaldehyde to enhance biosafety of diagnostic electron microscopy. Viruses 2015, 7, 666–679. [Google Scholar] [CrossRef] [Green Version]
- Dai, L.; Gao, G.F. Viral targets for vaccines against COVID-19. Nat. Rev. Immunol. 2021, 21, 73–82. [Google Scholar] [CrossRef]
- Chaplin, D.D. Overview of the immune response. J. Allergy Clin. Immunol. 2010, 125, S3–S23. [Google Scholar] [CrossRef]
- Gao, Q.; Bao, L.; Mao, H.; Wang, L.; Xu, K.; Yang, M.; Li, Y.; Zhu, L.; Wang, N.; Lv, Z.; et al. Development of an inactivated vaccine candidate for SARS-CoV-2. Science 2020, 369, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Ganneru, B.; Jogdand, H.; Daram, V.K.; Das, D.; Molugu, N.R.; Prasad, S.D.; Kannappa, S.V.; Ella, K.M.; Ravikrishnan, R.; Awasthi, A.; et al. Th1 skewed immune response of whole virion inactivated SARS CoV 2 vaccine and its safety evaluation. iScience 2021, 24, 102298. [Google Scholar] [CrossRef]
- Havervall, S.; Marking, U.; Greilert-Norin, N.; Ng, H.; Gordon, M.; Salomonsson, A.C.; Hellstrom, C.; Pin, E.; Blom, K.; Mangsbo, S.; et al. Antibody responses after a single dose of ChAdOx1 nCoV-19 vaccine in healthcare workers previously infected with SARS-CoV-2. EBioMedicine 2021, 70, 103523. [Google Scholar] [CrossRef]
- Zhang, Y.; Zeng, G.; Pan, H.; Li, C.; Hu, Y.; Chu, K.; Han, W.; Chen, Z.; Tang, R.; Yin, W.; et al. Safety, tolerability, and immunogenicity of an inactivated SARS-CoV-2 vaccine in healthy adults aged 18-59 years: A randomised, double-blind, placebo-controlled, phase 1/2 clinical trial. Lancet Infect. Dis. 2021, 21, 181–192. [Google Scholar] [CrossRef]
- Wilder-Smith, A.; Mulholland, K. Effectiveness of an Inactivated SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 385, 946–948. [Google Scholar] [CrossRef]
- Jara, A.; Undurraga, E.A.; Gonzalez, C.; Paredes, F.; Fontecilla, T.; Jara, G.; Pizarro, A.; Acevedo, J.; Leo, K.; Leon, F.; et al. Effectiveness of an Inactivated SARS-CoV-2 Vaccine in Chile. N. Engl. J. Med. 2021, 385, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Mariano, G.; Farthing, R.J.; Lale-Farjat, S.L.M.; Bergeron, J.R.C. Structural Characterization of SARS-CoV-2: Where We Are, and Where We Need to Be. Front. Mol. Biosci. 2020, 7, 605236. [Google Scholar] [CrossRef]
- Graham, R.L.; Baric, R.S. Recombination, reservoirs, and the modular spike: Mechanisms of coronavirus cross-species transmission. J. Virol. 2010, 84, 3134–3146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neuman, B.W.; Kiss, G.; Kunding, A.H.; Bhella, D.; Baksh, M.F.; Connelly, S.; Droese, B.; Klaus, J.P.; Makino, S.; Sawicki, S.G.; et al. A structural analysis of M protein in coronavirus assembly and morphology. J. Struct. Biol. 2011, 174, 11–22. [Google Scholar] [CrossRef]
- He, Y.; Zhou, Y.; Wu, H.; Luo, B.; Chen, J.; Li, W.; Jiang, S. Identification of immunodominant sites on the spike protein of severe acute respiratory syndrome (SARS) coronavirus: Implication for developing SARS diagnostics and vaccines. J. Immunol. 2004, 173, 4050–4057. [Google Scholar] [CrossRef]
- Sun, J.; Zhuang, Z.; Zheng, J.; Li, K.; Wong, R.L.; Liu, D.; Huang, J.; He, J.; Zhu, A.; Zhao, J.; et al. Generation of a Broadly Useful Model for COVID-19 Pathogenesis, Vaccination, and Treatment. Cell 2020, 182, 734–743.e5. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; He, Y.; Jiang, S.; Zheng, B.J. Development of subunit vaccines against severe acute respiratory syndrome. Drugs Today 2008, 44, 63–73. [Google Scholar] [CrossRef]
- Long, Q.X.; Liu, B.Z.; Deng, H.J.; Wu, G.C.; Deng, K.; Chen, Y.K.; Liao, P.; Qiu, J.F.; Lin, Y.; Cai, X.F.; et al. Antibody responses to SARS-CoV-2 in patients with COVID-19. Nat. Med. 2020, 26, 845–848. [Google Scholar] [CrossRef]
- Pinto, D.; Sauer, M.M.; Czudnochowski, N.; Siong Low, J.; Tortorici, M.A.; Housley, M.P.; Noack, J.; Walls, A.C.; Bowen, J.E.; Guarino, B.; et al. A human antibody that broadly neutralizes betacoronaviruses protects against SARS-CoV-2 by blocking the fusion machinery. bioRxiv 2021, 442808. [Google Scholar] [CrossRef]
- Akerstrom, S.; Tan, Y.J.; Mirazimi, A. Amino acids 15-28 in the ectodomain of SARS coronavirus 3a protein induces neutralizing antibodies. FEBS Lett. 2006, 580, 3799–3803. [Google Scholar] [CrossRef] [Green Version]
- Callaway, E. The race for coronavirus vaccines: A graphical guide. Nature 2020, 580, 576–577. [Google Scholar] [CrossRef]
- Bachmann, M.F.; Jennings, G.T. Vaccine delivery: A matter of size, geometry, kinetics and molecular patterns. Nat. Rev. Immunol. 2010, 10, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Tai, W.; Chen, J.; Zhao, G.; Geng, Q.; He, L.; Chen, Y.; Zhou, Y.; Li, F.; Du, L. Rational Design of Zika Virus Subunit Vaccine with Enhanced Efficacy. J. Virol. 2019, 93, e02187-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keech, C.; Albert, G.; Cho, I.; Robertson, A.; Reed, P.; Neal, S.; Plested, J.S.; Zhu, M.; Cloney-Clark, S.; Zhou, H.; et al. Phase 1-2 Trial of a SARS-CoV-2 Recombinant Spike Protein Nanoparticle Vaccine. N. Engl. J. Med. 2020, 383, 2320–2332. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Yisimayi, A.; Bai, Y.; Huang, W.; Li, X.; Zhang, Z.; Yuan, T.; An, R.; Wang, J.; Xiao, T.; et al. Humoral immune response to circulating SARS-CoV-2 variants elicited by inactivated and RBD-subunit vaccines. Cell Res. 2021, 31, 732–741. [Google Scholar] [CrossRef]
- Ramanathan, M.; Ferguson, I.D.; Miao, W.; Khavari, P.A. SARS-CoV-2 B.1.1.7 and B.1.351 spike variants bind human ACE2 with increased affinity. Lancet Infect. Dis. 2021, 21, 1070. [Google Scholar] [CrossRef]
- Antonelli, M.; Penfold, R.S.; Merino, J.; Sudre, C.H.; Molteni, E.; Berry, S.; Canas, L.S.; Graham, M.S.; Klaser, K.; Modat, M.; et al. Risk factors and disease profile of post-vaccination SARS-CoV-2 infection in UK users of the COVID Symptom Study app: A prospective, community-based, nested, case-control study. Lancet Infect. Dis. 2021. [Google Scholar] [CrossRef]
- Liu, H.; Wei, P.; Zhang, Q.; Aviszus, K.; Linderberger, J.; Yang, J.; Liu, J.; Chen, Z.; Waheed, H.; Reynoso, L.; et al. The Lambda variant of SARS-CoV-2 has a better chance than the Delta variant to escape vaccines. bioRxiv 2021. [Google Scholar] [CrossRef]
- Korber, B.; Fischer, W.M.; Gnanakaran, S.; Yoon, H.; Theiler, J.; Abfalterer, W.; Hengartner, N.; Giorgi, E.E.; Bhattacharya, T.; Foley, B.; et al. Tracking Changes in SARS-CoV-2 Spike: Evidence that D614G Increases Infectivity of the COVID-19 Virus. Cell 2020, 182, 812–827 e819. [Google Scholar] [CrossRef] [PubMed]
- Mandolesi, M.; Sheward, D.J.; Hanke, L.; Ma, J.; Pushparaj, P.; Perez Vidakovics, L.; Kim, C.; Adori, M.; Lenart, K.; Lore, K.; et al. SARS-CoV-2 protein subunit vaccination of mice and rhesus macaques elicits potent and durable neutralizing antibody responses. Cell Rep. Med. 2021, 2, 100252. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.M.; Wang, W.; Song, Z.G.; Hu, Y.; Tao, Z.W.; Tian, J.H.; Pei, Y.Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [Green Version]
- Bangaru, S.; Ozorowski, G.; Turner, H.L.; Antanasijevic, A.; Huang, D.; Wang, X.; Torres, J.L.; Diedrich, J.K.; Tian, J.H.; Portnoff, A.D.; et al. Structural analysis of full-length SARS-CoV-2 spike protein from an advanced vaccine candidate. Science 2020, 370, 1089–1094. [Google Scholar] [CrossRef]
- Dubin, G.; Toussaint, J.F.; Cassart, J.P.; Howe, B.; Boyce, D.; Friedland, L.; Abu-Elyazeed, R.; Poncelet, S.; Han, H.H.; Debrus, S. Investigation of a regulatory agency enquiry into potential porcine circovirus type 1 contamination of the human rotavirus vaccine, Rotarix: Approach and outcome. Hum. Vaccin. Immunother. 2013, 9, 2398–2408. [Google Scholar] [CrossRef] [Green Version]
- Folegatti, P.M.; Ewer, K.J.; Aley, P.K.; Angus, B.; Becker, S.; Belij-Rammerstorfer, S.; Bellamy, D.; Bibi, S.; Bittaye, M.; Clutterbuck, E.A.; et al. Safety and immunogenicity of the ChAdOx1 nCoV-19 vaccine against SARS-CoV-2: A preliminary report of a phase 1/2, single-blind, randomised controlled trial. Lancet 2020, 396, 467–478. [Google Scholar] [CrossRef]
- Voysey, M.; Clemens, S.A.C.; Madhi, S.A.; Weckx, L.Y.; Folegatti, P.M.; Aley, P.K.; Angus, B.; Baillie, V.L.; Barnabas, S.L.; Bhorat, Q.E.; et al. Safety and efficacy of the ChAdOx1 nCoV-19 vaccine (AZD1222) against SARS-CoV-2: An interim analysis of four randomised controlled trials in Brazil, South Africa, and the UK. Lancet 2021, 397, 99–111. [Google Scholar] [CrossRef]
- Sadoff, J.; Gray, G.; Vandebosch, A.; Cardenas, V.; Shukarev, G.; Grinsztejn, B.; Goepfert, P.A.; Truyers, C.; Fennema, H.; Spiessens, B.; et al. Safety and Efficacy of Single-Dose Ad26.COV2.S Vaccine against Covid-19. N. Engl. J. Med. 2021, 384, 2187–2201. [Google Scholar] [CrossRef]
- Yu, J.; Tostanoski, L.H.; Mercado, N.B.; McMahan, K.; Liu, J.; Jacob-Dolan, C.; Chandrashekar, A.; Atyeo, C.; Martinez, D.R.; Anioke, T.; et al. Protective efficacy of Ad26.COV2.S against SARS-CoV-2 B.1.351 in macaques. Nature 2021, 596, 423–427. [Google Scholar] [CrossRef]
- Takuva, S.; Takalani, A.; Garrett, N.; Goga, A.; Peter, J.; Louw, V.; Opie, J.; Jacobson, B.; Sanne, I.; Gail-Bekker, L.; et al. Thromboembolic Events in the South African Ad26.COV2.S Vaccine Study. N. Engl. J. Med. 2021, 385, 570–571. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Perez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Jackson, L.A.; Anderson, E.J.; Rouphael, N.G.; Roberts, P.C.; Makhene, M.; Coler, R.N.; McCullough, M.P.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; et al. An mRNA Vaccine against SARS-CoV-2—Preliminary Report. N. Engl. J. Med. 2020, 383, 1920–1931. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Schmidt, F.; Weisblum, Y.; Muecksch, F.; Barnes, C.O.; Finkin, S.; Schaefer-Babajew, D.; Cipolla, M.; Gaebler, C.; Lieberman, J.A.; et al. mRNA vaccine-elicited antibodies to SARS-CoV-2 and circulating variants. Nature 2021, 592, 616–622. [Google Scholar] [CrossRef]
- Lopez Bernal, J.; Andrews, N.; Gower, C.; Gallagher, E.; Simmons, R.; Thelwall, S.; Stowe, J.; Tessier, E.; Groves, N.; Dabrera, G.; et al. Effectiveness of Covid-19 Vaccines against the B.1.617.2 (Delta) Variant. N. Engl. J. Med. 2021, 385, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Joyce, M.; Chen, W.; Sankhala, R.; Hajduczki, A.; Thomas, P.; Choe, M.; Chang, W.; Peterson, C.E.; Martinez, E.; Morrison, E.B.; et al. SARS-CoV-2 ferritin nanoparticle vaccines elicit broad SARS coronavirus immunogenicity. bioRxiv 2021. [Google Scholar] [CrossRef]
- Klein, N.P.; Lewis, N.; Goddard, K.; Fireman, B.; Zerbo, O.; Hanson, K.E.; Donahue, J.G.; Kharbanda, E.O.; Naleway, A.; Nelson, J.C.; et al. Surveillance for Adverse Events After COVID-19 mRNA Vaccination. JAMA 2021, 14, 1390–1399. [Google Scholar] [CrossRef]
- Edara, V.; Lai, L.; Sahoo, S.; Floyd, K.; Sibai, M.; Solis, D.; Flowers, M.; Hussaini, L.; Ciric, C.R.; Bechnack, S.; et al. Infection and Vaccine-Induced Neutralizing Antibody Responses to the SARS-CoV-2 B.1.617.1 Variant. bioRxiv 2021. Available online: https://www.biorxiv.org/content/10.1101/2021.05.09.443299v1 (accessed on 9 September 2021).
- McDonald, I.; Murray, S.M.; Reynolds, C.J.; Altmann, D.M.; Boyton, R.J. Comparative systematic review and meta-analysis of reactogenicity, immunogenicity and efficacy of vaccines against SARS-CoV-2. NPJ Vaccines 2021, 6, 74. [Google Scholar] [CrossRef] [PubMed]
- Grannis, S.J.; Rowley, E.A.; Ong, T.C.; Stenehjem, E.; Klein, N.P.; DeSilva, M.; Naleway, A.L.; Natarajan, K.M.G.T. Interim Estimates of COVID-19 Vaccine Effectiveness Against COVID-19–Associated Emergency Department or Urgent Care Clinic Encounters and Hospitalizations Among Adults During SARS-CoV-2 B.1.617.2 (Delta) Variant Predominance—Nine States, June–August 2021. MMWR Morb Mortal Wkly Rep. 2021, 70, 1291. [Google Scholar] [CrossRef]
- Tre-Hardy, M.; Cupaiolo, R.; Wilmet, A.; Antoine-Moussiaux, T.; Della Vecchia, A.; Horeanga, A.; Papleux, E.; Vekemans, M.; Beukinga, I.; Blairon, L. Six-month interim analysis of ongoing immunogenicity surveillance of the mRNA-1273 vaccine in healthcare workers: A third dose is expected. J. Infect. 2021. [Google Scholar] [CrossRef] [PubMed]
- Wheatley, A.K.; Juno, J.A.; Wang, J.J.; Selva, K.J.; Reynaldi, A.; Tan, H.X.; Lee, W.S.; Wragg, K.M.; Kelly, H.G.; Esterbauer, R.; et al. Evolution of immune responses to SARS-CoV-2 in mild-moderate COVID-19. Nat. Commun. 2021, 12, 1162. [Google Scholar] [CrossRef] [PubMed]
- Dickey, T.H.; Tang, W.K.; Butler, B.; Ouahes, T.; Orr-Gonzalez, S.; Salinas, N.D.; Lambert, L.E.; Tolia, N.H. Design of the SARS-CoV-2 RBD Vaccine Antigen Improves Neutralizing Antibody Response. bioRxiv 2021. Available online: https://www.biorxiv.org/content/10.1101/2021.05.09.443238v1 (accessed on 9 September 2021). [CrossRef]
- Robbiani, D.F.; Gaebler, C.; Muecksch, F.; Lorenzi, J.C.C.; Wang, Z.; Cho, A.; Agudelo, M.; Barnes, C.O.; Gazumyan, A.; Finkin, S.; et al. Convergent antibody responses to SARS-CoV-2 in convalescent individuals. Nature 2020, 584, 437–442. [Google Scholar] [CrossRef]
- Yang, S.; Li, Y.; Dai, L.; Wang, J.; He, P.; Li, C.; Fang, X.; Wang, C.; Zhao, X.; Huang, E.; et al. Safety and immunogenicity of a recombinant tandem-repeat dimeric RBD-based protein subunit vaccine (ZF2001) against COVID-19 in adults: Two randomised, double-blind, placebo-controlled, phase 1 and 2 trials. Lancet Infect. Dis. 2021, 21, 1107–1119. [Google Scholar] [CrossRef]
- Jones, I.; Roy, P. Sputnik V COVID-19 vaccine candidate appears safe and effective. Lancet 2021, 397, 642–643. [Google Scholar] [CrossRef]

{kind=link}
| Vaccine | Phase | Platform | No. of Participants | No. of Doses | Location | Trial Number |
|---|---|---|---|---|---|---|
| Ad5-nCoV/ZF2001 prime-boost | IV | Inactivated | 27,711 | 2 | Brazil | NCT04747821 |
| AZLB ZF2001 | IV | RNA/Vector (non-replicating) | 10,000 | ≥1 | Denmark | NCT04760132 |
| Bharat Covaxin | IV | Inactivated | 4400 | 2 or 3 | China | NCT04863638 |
| BIBP BBIBP-CorV | IV | Inactivated | 2067 | 2 | Brazil | NCT04754698 |
| BIBP BBIBP-CorV | IV | RNA | 540 | 2 | Sweden | NCT04780659 |
| BioNTech BNT162 (b1/b2) | IV | Vector (non-replicating)/Protein subunit | 120 | 2 | China | NCT04833101 |
| BNT162/mRNA-1273/ChAdOx1-S | IV | RNA | 120 | 1 (boost) | Canada | NCT04885907 |
| CAMS vaccine | III | Vector (non-replicating) | 500,000 | 1 | South Africa | NCT04838795 |
| Cansino Ad5-nCoV | III | Protein subunit | 48,000 | 3 | Cuba | IG/CIGB-66I/CVD19/2103 |
| CIGB CIGB-66/Abdala | III | Inactivated | 45,000 | 2 | Bahrain, Jordan, Egypt, UAE | NCT04510207 |
| CureVac CVnCoV | III | Protein subunit | 44,010 | 2 or 3 | Cuba | IFV/COR/09 |
| Gamaleya Gam-COVID-Vac/Sputnik V | III | Vector (non-replicating) | 40,000 | 1 | Argentina, Chile, Mexico, others | NCT04526990 |
| Instituto Finlay de Vacunas FINLAY-FR-2 | III | Protein subunit | 40,000 | 3 | Pending | NCT04887207 |
| Janssen Ad26.COV2.S | III | Inactivated | 34,020 | 2 | Brazil, Malaysia | NCT04659239 |
| Janssen Ad26.COV2.S | III | Vector (non-replicating) | 33,758 | 2 | Russia | NCT04530396 |
| Moderna mRNA-1273 | III | Protein subunit | 33,000 | 2 | USA, Mexico, Puerto Rico | NCT04611802 |
| Novavax NVX-CoV2373 | III | Vector (non-replicating) | 30,000 | 2 | USA, Belgium, Brazil, others | NCT04614948 |
| Novavax NVX-CoV2373 | III | Protein subunit | 29,000 | Unclear | China, Ecuador, Indonesia, others | NCT04646590 |
| PLA-AMS ARCoV | III | RNA | 28,000 | 2 | Pending | NCT04847102 |
| RIBSP QazCOVID-in | III | Inactivated | 28,000 | 2 | Pending | NCT04852705 |
| Shenzhen Kangtai KCONVAC | III | Inactivated | 25,800 | 2 | India | NCT04641481 |
| Sinovac CoronaVac | III | Protein subunit | 15,000 | 2 | UK | NCT04583995 |
| Sinovac CoronaVac | III | Inactivated | 13,000 | 2 | Turkey | NCT04582344 |
| Sinovac CoronaVac | III | Inactivated | 12,000 | 2 | Peru | NCT04612972 |
| Vector Institute EpiVacCorona | III | Inactivated/Vector (non-replicating) | 4000 | 2 | UK | NCT04864561 |
| VLA2001/ChAdOx1-S | III | Inactivated | 3000 | 2 | Argentina | NCT04560881 |
| West China Hospital protein subunit vaccine | III | Inactivated | 3000 | 2 | Kazakhstan | NCT04691908 |
| WIBP vaccine | III | Protein subunit | 3000 | 2 | Russia | NCT04780035 |
| WIBP/BIBP vaccines | III | RNA | 2520 | 2 | Germany | NCT04674189 |
| WIBP/BIBP vaccines | III | Inactivated | 600 | 2 | Morocco | ChiCTR2000039000 |
| Vaccine | Efficacy | Source |
|---|---|---|
| Anhui Zhifei Longcom Biopharmaceutical ZF2001 | 76% | Primary efficacy analysis [35] |
| Bharat Covaxin/BBV152 | 81% | Press release: 3 March 2021 |
| BioNTech/Pfizer BNT162b2 | 95% | Primary efficacy analysis [36] |
| Beijing/Sinopharm BBIBP-CorV | 79% | Press release: 30 December 2020 |
| Cansino Ad5-nCoV | 65.28% | Press release: 24 February 2021 |
| CIGB CIGB-66/Abdala | 92.28% | Press release: 22 June 2021 |
| Chumakov Center CoviVac | Pending | WHO; COVID-19 tracker |
| Gamaleya Gam-COVID-Vac/Sputnik V | 92% | Primary efficacy analysis [37] |
| Janssen Ad26.COV2.S | 67% | Primary efficacy analysis [38] |
| Moderna mRNA-1273 | 94% | Primary efficacy analysis [39] |
| Oxford/AstraZeneca ChAdOx1-S | 62–90% | Interim data [40] |
| Research Institute for Biological Safety Problems QazCOVID-in | Pending | WHO; COVID-19 tracker |
| Sinovac CoronaVac | 50% | Press release: 3 April 2021 |
| Vector Institute EpiVacCorona | Pending | WHO; COVID-19 tracker |
| CoronaVac/Sinopharm vaccine | Pending | WHO; COVID-19 tracker [12] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Currier, A.W.; Jeshurin, M.C.; Sampson, V.B. SARS-CoV-2 Targets and COVID-19 Vaccines. COVID 2021, 1, 608-621. https://doi.org/10.3390/covid1030051
Currier AW, Jeshurin MC, Sampson VB. SARS-CoV-2 Targets and COVID-19 Vaccines. COVID. 2021; 1(3):608-621. https://doi.org/10.3390/covid1030051
Chicago/Turabian StyleCurrier, Arthur W., Madeline C. Jeshurin, and Valerie B. Sampson. 2021. "SARS-CoV-2 Targets and COVID-19 Vaccines" COVID 1, no. 3: 608-621. https://doi.org/10.3390/covid1030051
APA StyleCurrier, A. W., Jeshurin, M. C., & Sampson, V. B. (2021). SARS-CoV-2 Targets and COVID-19 Vaccines. COVID, 1(3), 608-621. https://doi.org/10.3390/covid1030051