
Effect of kuratsuki Bacillus and Priestia on Taste of Sake


Abstract
1. Introduction
2. Materials and Methods
2.1. Genome Sequencing of the kuratsuki Bacterial Strains A-10 and B-12
2.2. Cultivation of the Microorganisms
2.3. Measurement of Brix and Acidity
2.4. Estimation of the Sake Taste
2.5. Statistical Analysis
3. Results
3.1. Genome Characteristics
3.2. Brix and Acidity Changes
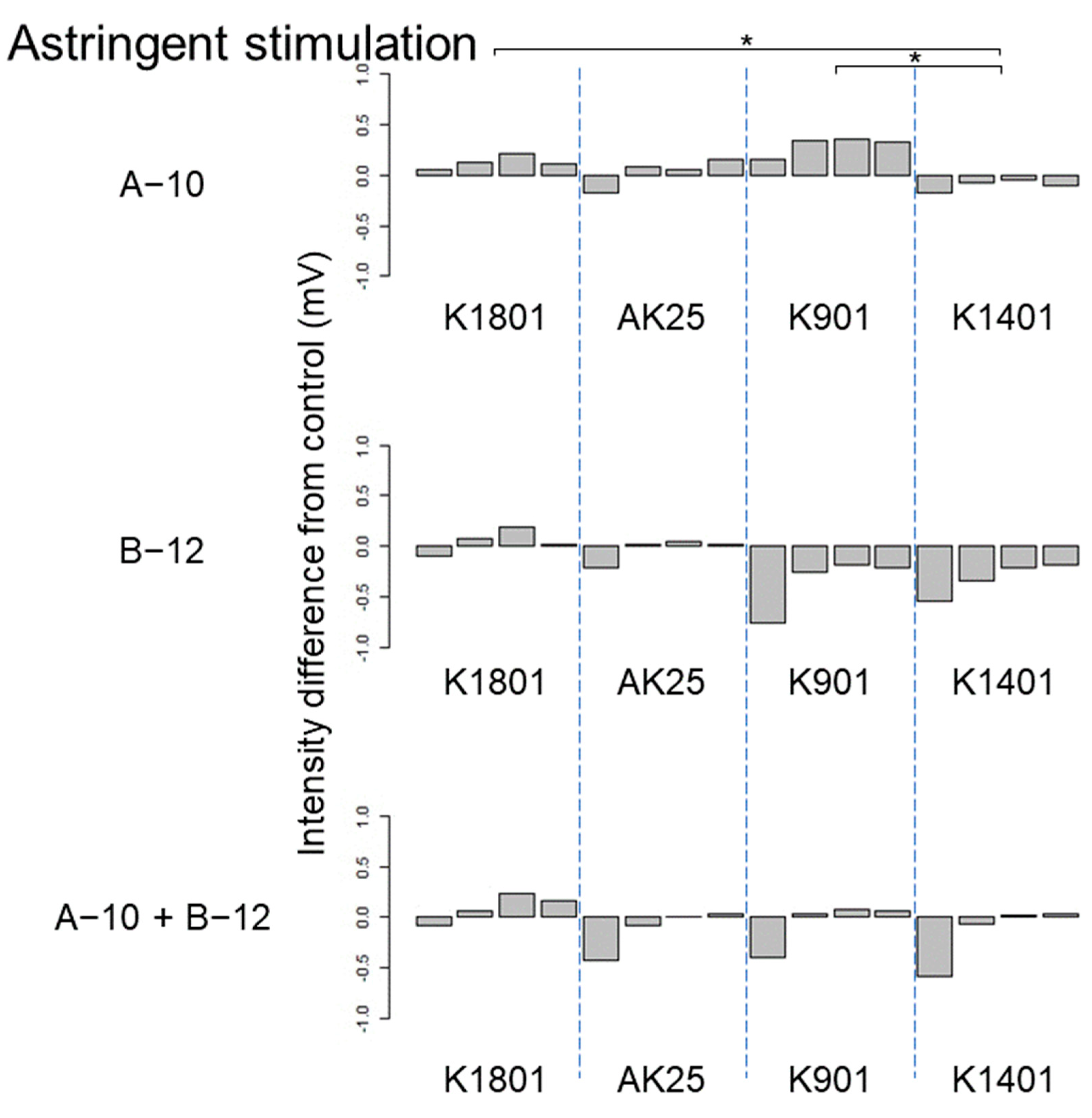
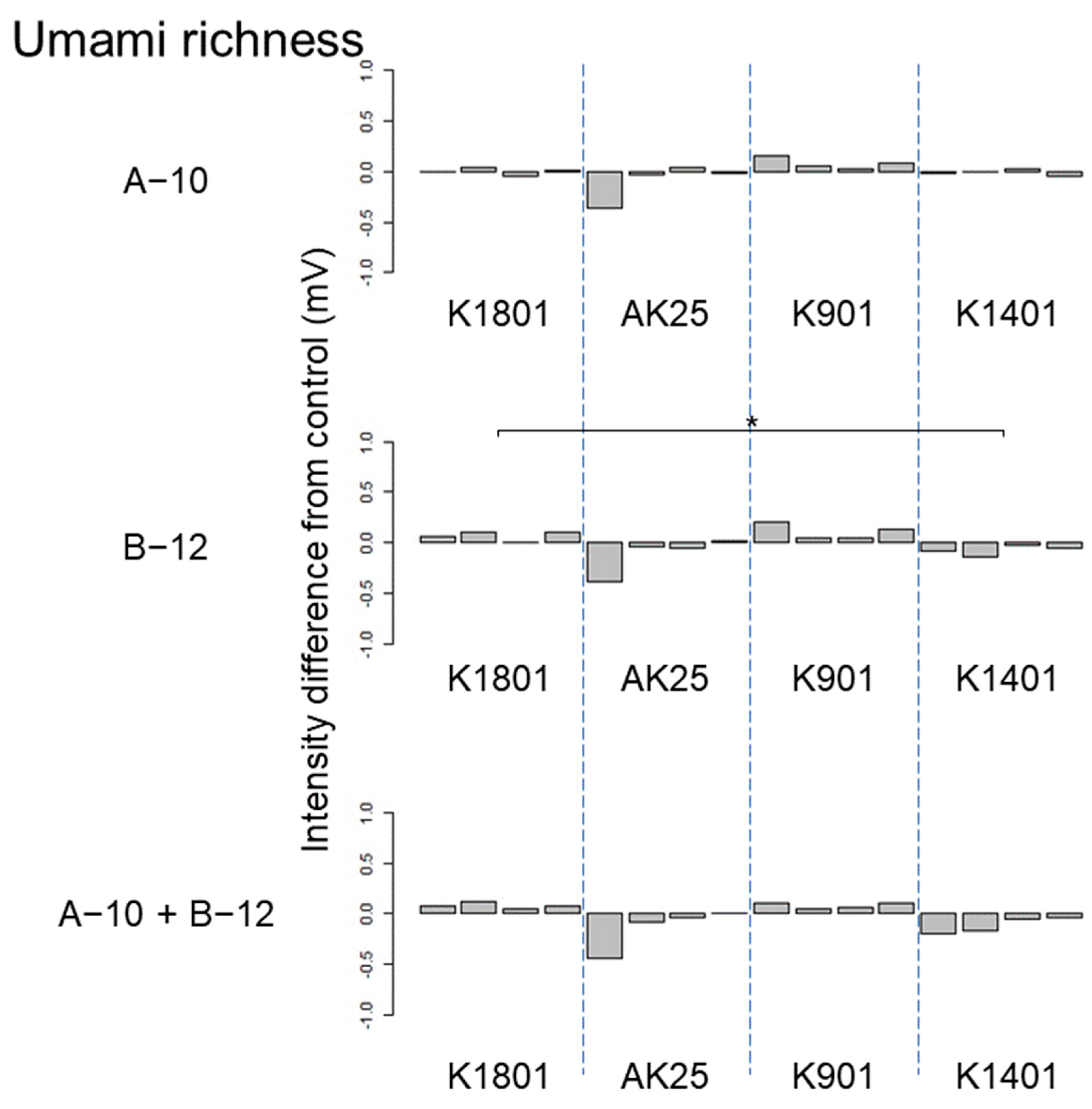
3.3. Effects of kuratsuki Bacillus and Priestia on Sake Taste
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Koyanagi, T.; Kiyohara, M.; Matsui, H.; Yamamoto, K.; Kondo, T.; Katayama, T.; Kumagai, H. Pyrosequencing survey of the microbial diversity of ‘narezushi’, an archetype of modern Japanese sushi. Lett. Appl. Microbiol. 2011, 53, 635–640. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Bamforth, C.W.; Mills, D.A. A review of molecular methods for microbial community profiling of beer and wine. J. Am. Soc. Brew. Chem. 2012, 70, 150–162. [Google Scholar] [CrossRef]
- Koyanagi, T.; Nakagawa, A.; Kiyohara, M.; Matsui, H.; Yamamoto, K.; Barla, F.; Take, H.; Katsuyama, Y.; Tsuji, A.; Shijimaya, M.; et al. Pyrosequencing analysis of microbiota in Kaburazushi, a traditional medieval sushi in Japan. Biosci. Biotechnol. Biochem. 2013, 77, 2125–2130. [Google Scholar] [CrossRef][Green Version]
- Kyung, K.H.; Pradas, E.M.; Kim, S.G.; Lee, Y.J.; Kim, K.H.; Choi, J.J.; Cho, J.H.; Chung, C.H.; Barrangou, R.; Breidt, F. Microbial ecology of watery kimuchi. J. Food Sci. 2015, 80, M1031–M1038. [Google Scholar] [CrossRef] [PubMed]
- Tamang, J.P.; Watanabe, K.; Holzapfel, W.H. Review: Diversity of microorganisms in global fermented foods and beverages. Front. Microbiol. 2016, 7, 377. [Google Scholar] [CrossRef]
- Wuyts, S.; Van Beeck, W.; Allonsius, C.N.; van den Broek, M.F.; Lebeer, S. Applications of plant-based fermented foods and their microbes. Curr. Opin. Biotechnol. 2020, 61, 45–52. [Google Scholar] [CrossRef]
- Whon, T.W.; Ahn, S.W.; Yang, S.; Kim, J.Y.; Kim, Y.B.; Kim, Y.; Hong, J.-M.; Jung, H.; Choi, Y.-E.; Lee, S.H.; et al. ODFM, an omics data resource from microorganisms associated with fermented foods. Sci. Data 2021, 8, 113. [Google Scholar] [CrossRef] [PubMed]
- Rodpai, R.; Sanpool, O.; Thanchomnang, T.; Wangwiwatsin, A.; Sadaow, L.; Phupiewkham, W.; Boonroumkaew, P.; Intapan, P.M.; Maleewong, W. Investigating the microbiota of fermented fish products (Pla-ra) from different communities of northeastern Thailand. PLoS ONE 2021, 16, e0245227. [Google Scholar] [CrossRef]
- Fraiture, M.-A.; Papazova, N.; Roosens, N.H.C. DNA walking strategy to identify unauthorized genetically modified bacteria in microbial fermentation products. Int. J. Food Microbiol. 2021, 337, 108913. [Google Scholar] [CrossRef]
- Walsh, A.M.; Leech, J.; Huttenhower, C.; Delhomme-Nguyen, H.; Crispie, F.; Chervaux, C.; Cotter, P.D. Integrated molecular approaches for fermented food microbiome research. FEMS Microbiol. Rev. 2023, 47, fuad001. [Google Scholar] [CrossRef]
- Le, M.-M.; Zhong, L.-W.; Ren, Z.-W.; An, M.-Q.; Long, Y.-H.; Ling, T.-J. Dynamic changes in the microbial community and metabolite profile during the pile fermentation process of Fuzhuan brink tea. J. Agric. Food Chem. 2023, 71, 19142–19153. [Google Scholar] [CrossRef] [PubMed]
- Fusco, V.; Fanelli, F.; Chieffi, D. Recent and advanced DNA-based technologies for the authentication of probiotic, protected designation of origin (PDO) and protected geographical indication (PGI) fermented foods and beverages. Foods 2023, 12, 3782. [Google Scholar] [CrossRef] [PubMed]
- Palmnäs-Bédard, M.; de Santa Izabel, A.; Dicksved, J.; Landberg, R. Characterization of the bacterial composition of 47 fermented foods in Sweden. Foods 2023, 12, 3827. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Ohta, M.; Lee, M.; Mills, D.A. Indigenous bacteria and fungi drive traditional kimoto sake fermentations. Appl. Environ. Microbiol. 2014, 80, 5522–5529. [Google Scholar] [CrossRef]
- Koyanagi, T.; Nakagawa, A.; Kiyohara, M.; Matsui, H.; Tsuji, A.; Barla, F.; Take, H.; Katsuyama, Y.; Tokuda, K.; Nakamura, S.; et al. Tracing microbiota changes in yamahai-moto, the traditional Japanese sake starter. Biosci. Biotechnol. Biochem. 2016, 80, 399–406. [Google Scholar] [CrossRef]
- Tsuji, A.; Kozawa, M.; Tokuda, K.; Enomoto, T.; Koyanagi, T. Robust domination of Lactobacillus sakei in microbiota during traditional Japanese sake starter yamahai-moto fermentation and the accompanying changes in metabolites. Curr. Microbiol. 2018, 75, 1498–1505. [Google Scholar] [CrossRef]
- Terasaki, M.; Nishida, H. Bacterial DNA diversity among clear and cloudy sakes, and sake-kasu. Open Bioinform. J. 2020, 13, 74–82. [Google Scholar] [CrossRef]
- Ito, K.; Niwa, R.; Kobayashi, K.; Nakagawa, T.; Hoshino, G.; Tsuchida, Y. A dark matter in sake brewing: Origin of microbes producing a Kimoto-style fermentation starter. Front. Microbiol. 2023, 14, 1112638. [Google Scholar] [CrossRef]
- Nishida, H. Sake brewing and bacteria inhabiting sake breweries. Front. Microbiol. 2021, 12, 602380. [Google Scholar] [CrossRef]
- Terasaki, M.; Kimura, Y.; Yamada, M.; Nishida, H. Genomic information of Kocuria isolates from sake brewing process. AIMS Microbiol. 2021, 7, 114–123. [Google Scholar] [CrossRef]
- Kanamoto, E.; Terashima, K.; Shiraki, Y.; Nishida, H. Diversity of Bacillus isolates from the sake brewing process at a sake brewery. Microorganisms 2021, 9, 1760. [Google Scholar] [CrossRef] [PubMed]
- Nishida, H. Kuratsuki bacteria and sake making. Biosci. Biotechnol. Biochem. 2023, zbad147. [Google Scholar] [CrossRef]
- Park, E.-J.; Roh, S.W.; Kim, M.-S.; Jung, M.-J.; Shin, K.-S.; Bae, J.-W. Kocuria koreensis, sp. nov., isolated from fermented seafood. Int. J. Syst. Evol. Microbiol. 2010, 60, 140–143. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Braun, M.S.; Wang, E.; Zimmermann, S.; Boutin, S.; Wink, M. Kocuria uropygioeca sp. nov. and Kocuria uropygialis sp. nov., isolated form the preen glands of Great Spotted Woodpeckers (Dendrocopos major). Syst. Appl. Microbiol. 2018, 41, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.S.; Patel, S.; Saini, N.; Chen, S. Robust demarcation of 17 distinct Bacillus species clades, proposed as novel Bacillaceae genera, by phylogenomics and comparative genomic analyses: Description of Robertmurraya kyonggiensis sp. nov. and proposal for an emended genus Bacillus limiting it only to the members of the Subtilis and Cereus clades of species. Int. J. Syst. Evol. Microbiol. 2020, 70, 5753–5798. [Google Scholar]
- Park, Y.C.; Shaffer, C.E.H.; Bennett, G.N. Microbial formation of esters. Appl. Microbiol. Biotechnol. 2009, 85, 13–25. [Google Scholar] [CrossRef]
- Saerens, S.M.G.; Delvaux, F.R.; Verstrepen, K.J.; Thevelein, J.M. Production and biological function of volatile esters in Saccharomyces cerevisiae. Microb. Biotechnol. 2010, 3, 165–177. [Google Scholar] [CrossRef]
- Procopio, S.; Qian, F.; Becker, T. Functional and regulation of yeast genes involved in higher alcohol and ester metabolism during beverage fermentation. Eur. Food Res. Technol. 2011, 233, 721–729. [Google Scholar] [CrossRef]
- Kitagaki, H.; Kitamoto, K. Breeding research on sake yeasts in Japan: History, recent technological advances, and future perspectives. Ann. Rev. Food Sci. Technol. 2013, 4, 215–235. [Google Scholar] [CrossRef]
- Dzialo, M.C.; Park, R.; Steensels, J.; Lievens, B.; Verstrepen, K.J. Physiology, ecology and industrial applications of aroma formation in yeast. FEMS Microbiol. Rev. 2017, 41, S95–S128. [Google Scholar] [CrossRef]
- Suto, M.; Kawashima, H. Compound specific carbon isotope analysis in sake by LC/IRMS and brewers’ alcohol proportion. Sci. Rep. 2019, 9, 17635. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Hu, W.; Xia, Y.; Mu, Z.; Tao, L.; Song, X.; Zhang, H.; Ni, B.; Ai, L. Flavor formation in Chinese rice wine (Huangjiu): Impacts of the flavor-active microorganisms, raw materials, and fermentation technology. Front. Microbiol. 2020, 11, 580247. [Google Scholar] [CrossRef] [PubMed]
- Suto, M.; Kawashima, H. Discrimination for sake brewing methods by compound specific isotope analysis and formation mechanism of organic acids in sake. Food Chem. 2022, 381, 132295. [Google Scholar] [CrossRef] [PubMed]
- Tekarslan-Sahin, S.H. Adaptive laboratory evolution of yeasts for aroma compound production. Fermentation 2022, 8, 372. [Google Scholar] [CrossRef]
- Yoshimoto, H.; Bogaki, T. Mechanisms of production and control of acetate esters in yeasts. J. Biosci. Bioeng. 2023, 136, 261–269. [Google Scholar] [CrossRef]
- Maruyama, H. Beer brewed with sake yeast strain has unique sake-like flavors. J. Am. Soc. Brew. Chem. 2023; in press. [Google Scholar] [CrossRef]
- Gonçalves, M.; Pontes, A.; Almeida, P.; Barbosa, R.; Serra, M.; Libkind, D.; Hutzler, M.; Gonçalves, P.; Sampaio, J.P. Distinct domestication trajectories in top-fermenting beer yeasts and wine yeasts. Curr. Biol. 2016, 26, 2750–2761. [Google Scholar] [CrossRef]
- Mizoguchi, H.; Ikeda, T.; Hara, S. Differences in the intracellular lipids of sake yeast in main mash seeded respectively with two kinds of seed mash: Kimoto and sokujo-moto. J. Ferment. Bioeng. 1995, 80, 586–591. [Google Scholar] [CrossRef]
- Sawada, K.; Sato, T.; Harajima, H.; Jayakody, L.N.; Hirata, M.; Yamashiro, M.; Tajima, M.; Mitsutake, S.; Nagao, K.; Tsuge, K.; et al. Glucosylceramide contained in koji mold-cultured cereal confers membrane and flavor modification and stress tolerance to Saccharomyces cerevisiae during coculture fermentation. Appl. Environ. Microbiol. 2015, 81, 3688–3698. [Google Scholar] [CrossRef]
- Huang, Z.-R.; Hong, J.-L.; Xu, J.-X.; Li, L.; Guo, W.-L.; Pan, Y.-Y.; Chen, S.-J.; Bai, W.-D.; Rao, P.-F.; Ni, L.; et al. Exploring core functional microbiota responsible for the production of volatile flavor during the traditional brewing of Wuyi Hong Qu glutinous rice wine. Food Microbiol. 2018, 76, 487–496. [Google Scholar] [CrossRef]
- Xu, J.-Z.; Zhang, Y.-Y.; Zhang, W.-G. Correlation between changes in flavor compounds and microbial community ecological succession in the liquid fermentation of rice wine. World J. Microbiol. Biotechnol. 2023, 40, 17. [Google Scholar] [CrossRef] [PubMed]
- Ito, K.; Niwa, R.; Yamagishi, Y.; Kobayashi, K.; Tsuchida, Y.; Hoshino, G.; Nakagawa, T.; Watanabe, T. A unique case in which Kimoto-style fermentation was completed with Leuconostoc as the dominant genus without transitioning to Lactobacillus. J. Biosci. Bioeng. 2023, 135, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, H.; Watanabe, K.; Wakai, Y. Combination of four bacterial strains isolated from Yamahai-shubo in traditional Japanese sake brewing. Food Sci. Nutr. 2023, 11, 2990–3001. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, D.; Kumano, M.; Sugimoto, Y.; Ito, M.; Ohashi, M.; Sunada, K.; Takahashi, T.; Yamada, T.; Takagi, H. Metabolic switching of sake yeast by kimoto lactic acid bacteria through the [GAR+] non-genetic element. J. Biosci. Bioeng. 2018, 126, 624–629. [Google Scholar] [CrossRef]
- Watanabe, D.; Takagi, H. Yeast prion-based metabolic reprogramming induced by bacteria in fermented foods. FEMS Yeast Res. 2019, 19, foz061. [Google Scholar] [CrossRef]
- Terasaki, M.; Inoue, A.; Kanamoto, E.; Yoshida, S.; Yamada, M.; Toda, H.; Nishida, H. Co-cultivation of sake yeast and Kocuria isolates from the sake brewing process. FEMS Microbiol. Lett. 2021, 368, fnab053. [Google Scholar] [CrossRef]
- Yazaki, A.; Nishida, H. Effect of kuratsuki Kocuria on sake brewing in different koji conditions. FEMS Microbiol. Lett. 2023, 370, fnad020. [Google Scholar] [CrossRef]
- Yazaki, A.; Nishida, H. Effect of kuratsuki Kocuria on sake’s taste varies depending on the sake yeast strain used in sake brewing. Arch. Microbiol. 2023, 205, 290. [Google Scholar] [CrossRef]
- Saito, M.; Nishida, H. Molecular hydrogen treatment of sake yeast and kuratsuki bacteria affects sake taste. Fermentation 2023, 9, 516. [Google Scholar] [CrossRef]
- Toko, K. Taste sensor. Sens. Actuators B Chem. 2000, 64, 205–215. [Google Scholar] [CrossRef]
- Toko, K. Research and development of taste sensors as a novel analytical tool. Proc. Jpn. Acad. Ser. B 2023, 99, 173–189. [Google Scholar] [CrossRef] [PubMed]
- Wick, R.R.; Judd, L.M.; Cerdeira, L.T.; Hawkey, J.; Méric, G.; Vezina, B.; Wyres, K.L.; Holt, K.E. Trycycler: Consensus long-read assemblies for bacterial genomes. Genome Biol. 2021, 22, 266. [Google Scholar] [CrossRef]
- Hunt, M.; De Silva, N.; Otto, T.D.; Parkhill, J.; Keane, J.A.; Harris, S.R. Circlator: Automated circularization of genome assemblies using long sequencing reads. Genome Biol. 2015, 16, 294. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler Transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef]
- Ohya, Y.; Kashima, M. History, lineage and phenotypic differentiation of sake yeast. Biosci. Biotechnol. Biochem. 2019, 83, 1442–1448. [Google Scholar] [CrossRef]
- Zhang, K.; Wu, W.; Yan, Q. Research advances on sake rice, koji, and sake yeast: A review. Food Sci. Nutr. 2020, 8, 2995–3003. [Google Scholar] [CrossRef] [PubMed]
- Negoro, H.; Ishida, H. Development of sake yeast breeding and analysis of genes related to its various phenotypes. FEMS Yeast Res. 2022, 22, foac057. [Google Scholar] [CrossRef]
- Capozzi, V.; Spano, G. Food microbial biodiversity and “microbes of protected origin”. Front. Microbiol. 2011, 2, 237. [Google Scholar] [CrossRef]
- Capozzi, V.; Fragasso, M.; Russo, P. Microbiological safety and the management of microbial resources in artisanal foods and beverages: The need for a transdisciplinary assessment to conciliate actual trends and risks avoidance. Microorganisms 2020, 8, 306. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain A-10 | |
| Total length (bp) | 3,749,764 |
| No. of sequences | 1 |
| Accession number | BSYL01000001 |
| GC content (%) | 41.6 |
| No. of CDSs | 3750 |
| No. of rRNA | 24 |
| No. of tRNA | 81 |
| Coding ratio (%) | 87.6 |
| Transposase gene | IS3 family: LOCUS_02590, LOCUS_06730, LOCUS_07270, LOCUS_25590 |
| Strain B-12 | |
| Total length (bp) | 5,420,896 |
| No. of sequences | 8 |
| Accession number | BSYK01000001-BSYK01000008 |
| GC content (%) | 38.2 |
| No. of CDSs | 5506 |
| No. of rRNA | 39 |
| No. of tRNA | 132 |
| Coding ratio (%) | 83.3 |
| Transposase gene | IS4 family: LOCUS_34940, LOCUS_52540, LOCUS_52940, LOCUS_54130, LOCUS_54850, LOCUS_54860, LOCUS_55060 IS110 family: LOCUS_05740, LOCUS_10520, LOCUS_31750, LOCUS_32200, LOCUS_42770, LOCUS_54720 IS21 family: LOCUS_52840, LOCUS_53190, LOCUS_53360 IS1326 family: LOCUS_53690, LOCUS_54170 IS3 family: LOCUS_36670 IS466 family: LOCUS_53420 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kobayashi, K.; Nishida, H. Effect of kuratsuki Bacillus and Priestia on Taste of Sake. Appl. Microbiol. 2024, 4, 147-161. https://doi.org/10.3390/applmicrobiol4010011
Kobayashi K, Nishida H. Effect of kuratsuki Bacillus and Priestia on Taste of Sake. Applied Microbiology. 2024; 4(1):147-161. https://doi.org/10.3390/applmicrobiol4010011
Chicago/Turabian StyleKobayashi, Karin, and Hiromi Nishida. 2024. "Effect of kuratsuki Bacillus and Priestia on Taste of Sake" Applied Microbiology 4, no. 1: 147-161. https://doi.org/10.3390/applmicrobiol4010011
APA StyleKobayashi, K., & Nishida, H. (2024). Effect of kuratsuki Bacillus and Priestia on Taste of Sake. Applied Microbiology, 4(1), 147-161. https://doi.org/10.3390/applmicrobiol4010011