
Effects of Water Stress and Mulch Type on Linseed Seed Yield, Physiological Traits, and Oil Compounds

 ,
,  , ,
, ,  ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Growing Conditions and Experimental Design
2.2. Growth Parameters
2.3. Oil Content and Oilseed Yield
2.4. Fatty Acids
2.5. Relative Water Content (RWC)
2.6. Chlorophyll and Carotenoid Content
2.7. Total Soluble Sugar Content
2.8. Proline Content
2.9. Antioxidant Enzyme Extractions and Assays
2.10. Total Polyphenol Content
2.11. DPPH (2,2-Diphenyl-1-picrylhydrazyl-hydrate) Radical Scavenging Activity
2.12. Data Statistical Processing
3. Results
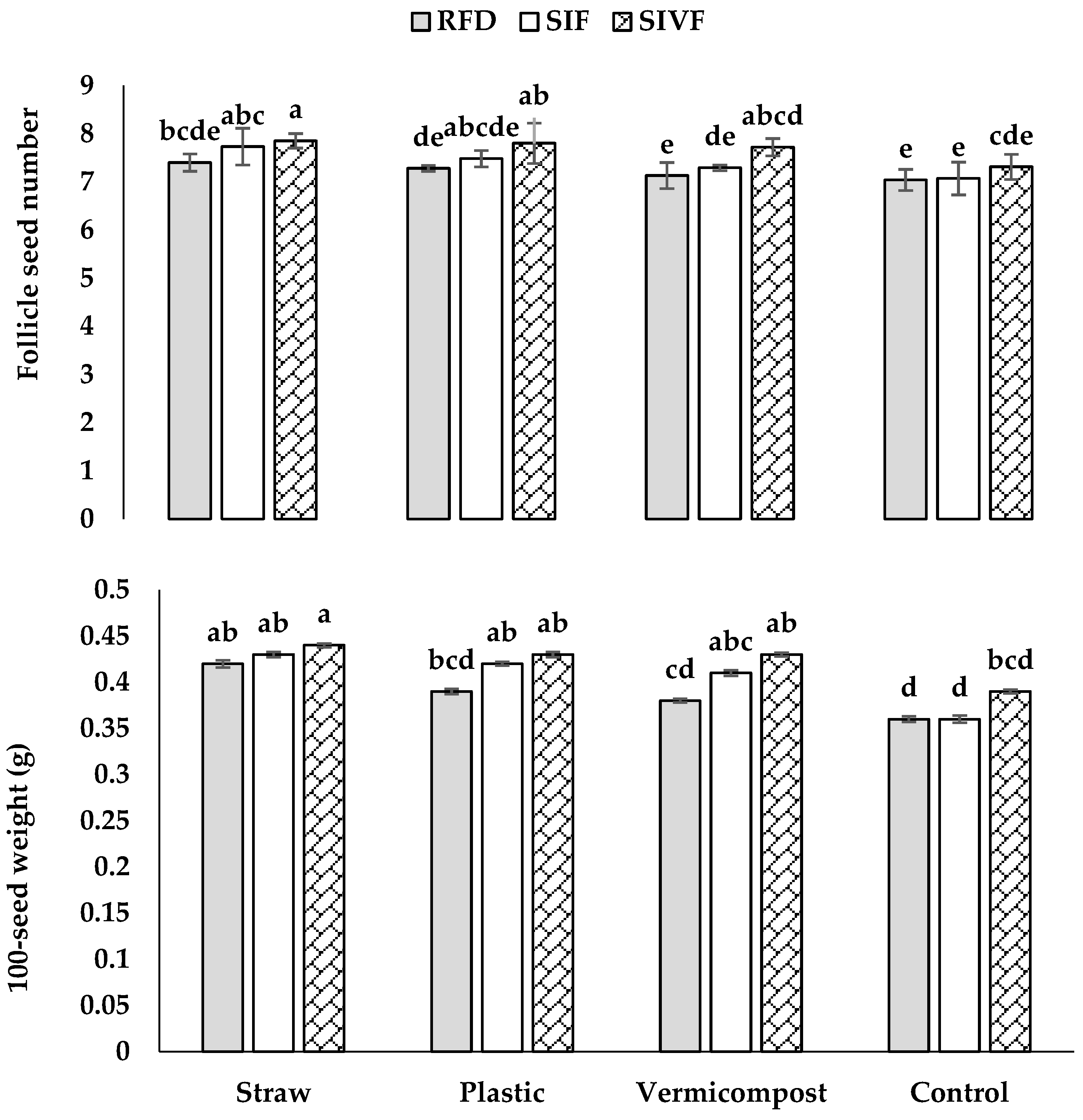
3.1. Biometrical and Seed Yield Parameters
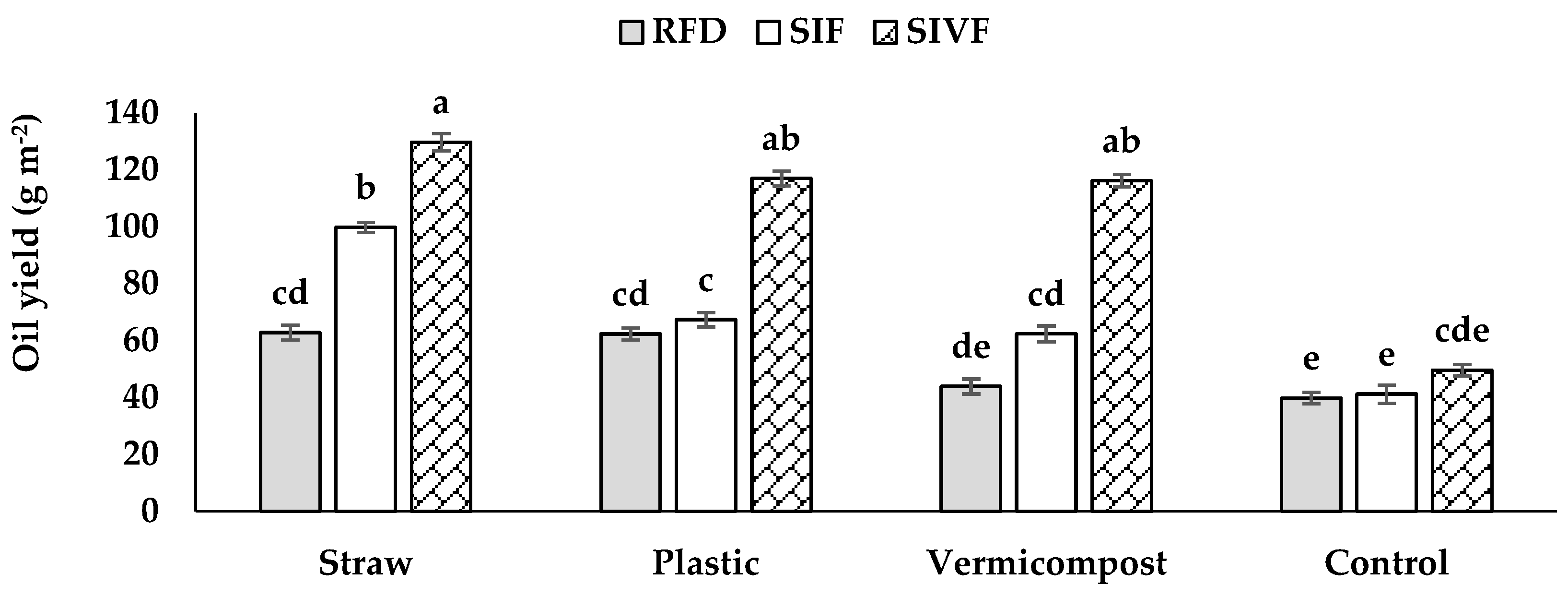
3.2. Oilseed Content, Yield, and Fatty Acid Composition
3.3. Physiological Traits
Relative Water Content
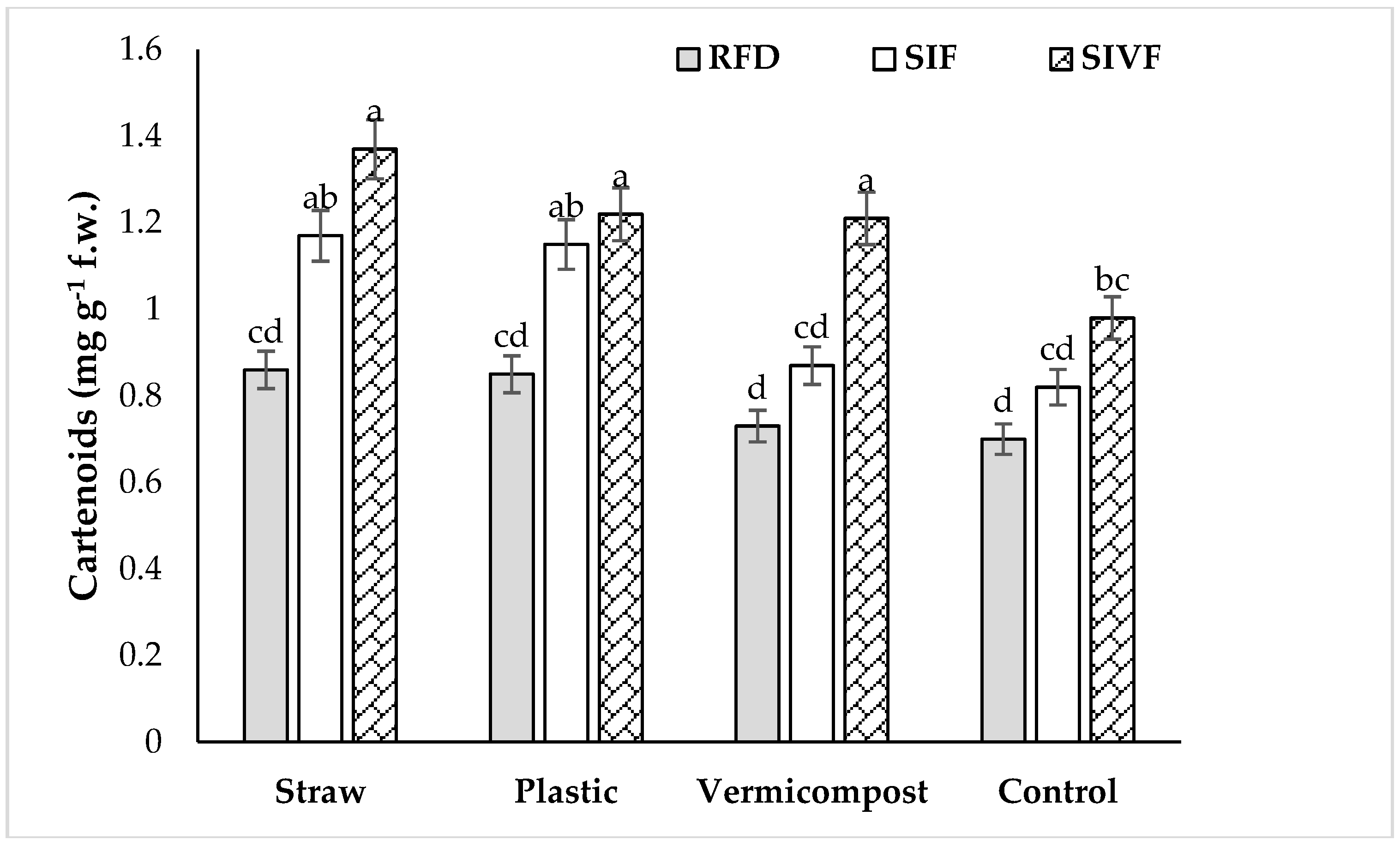
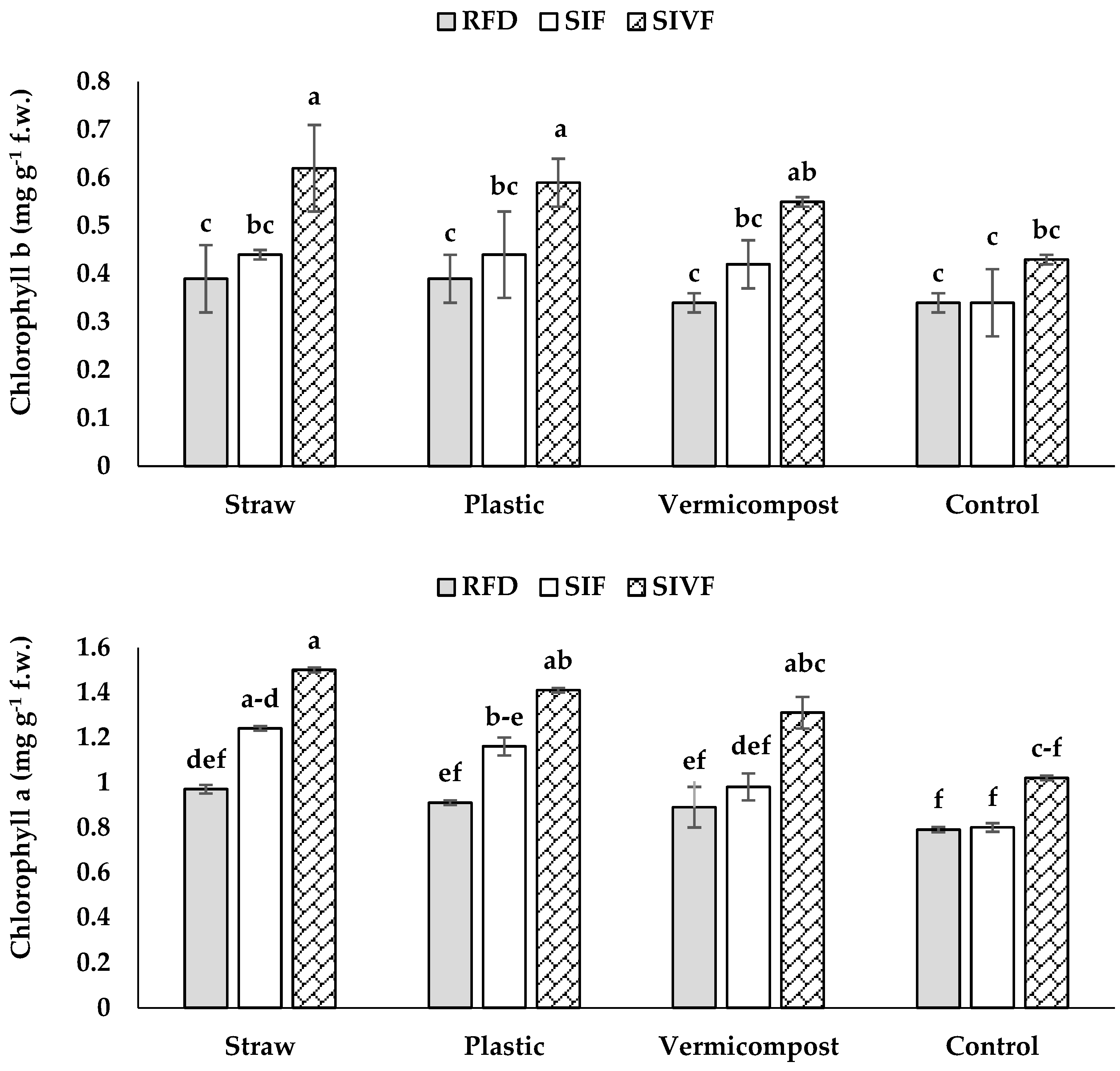
3.4. Photosynthetic Pigments
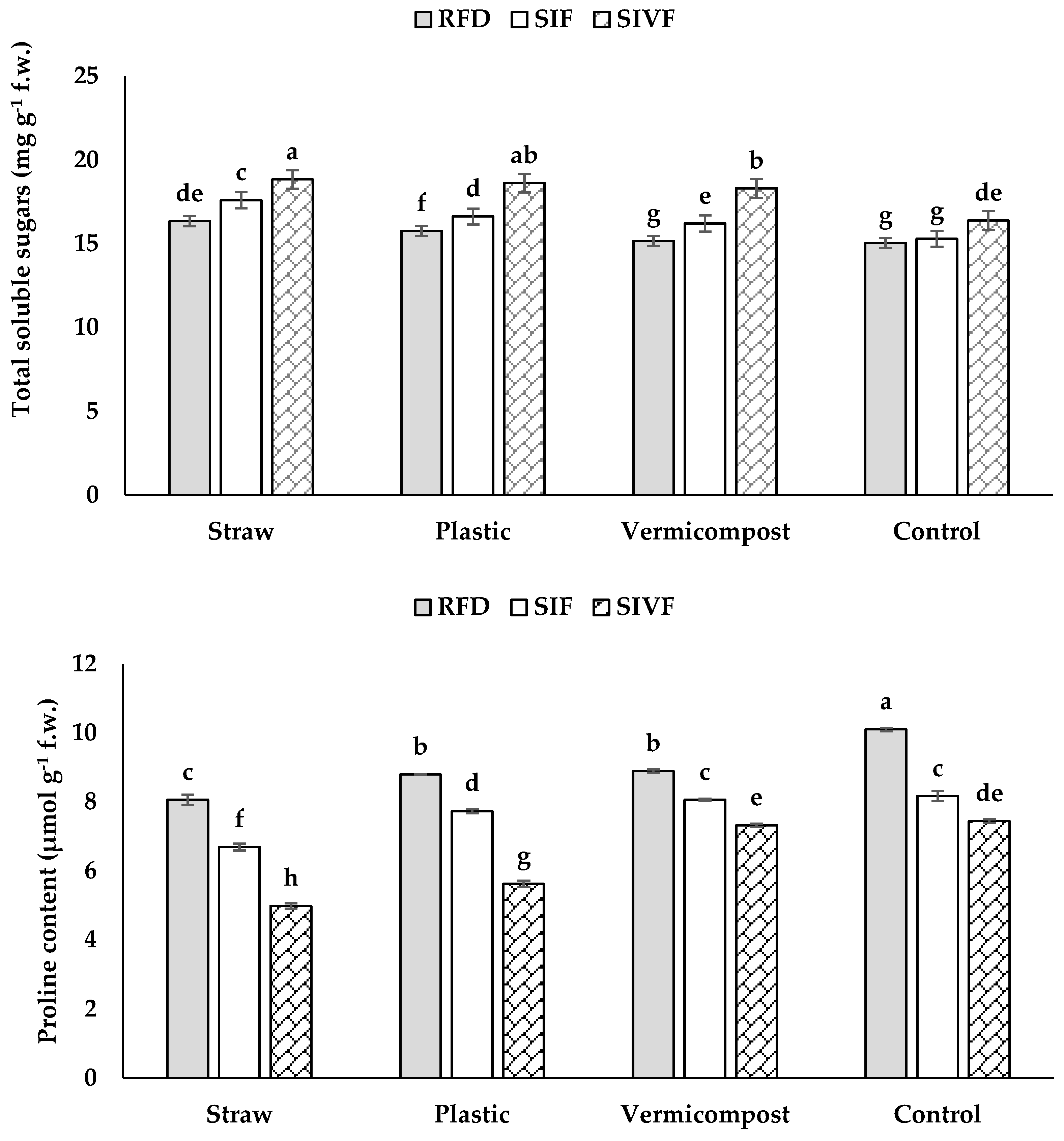
3.5. Total Soluble Sugars and Proline Content
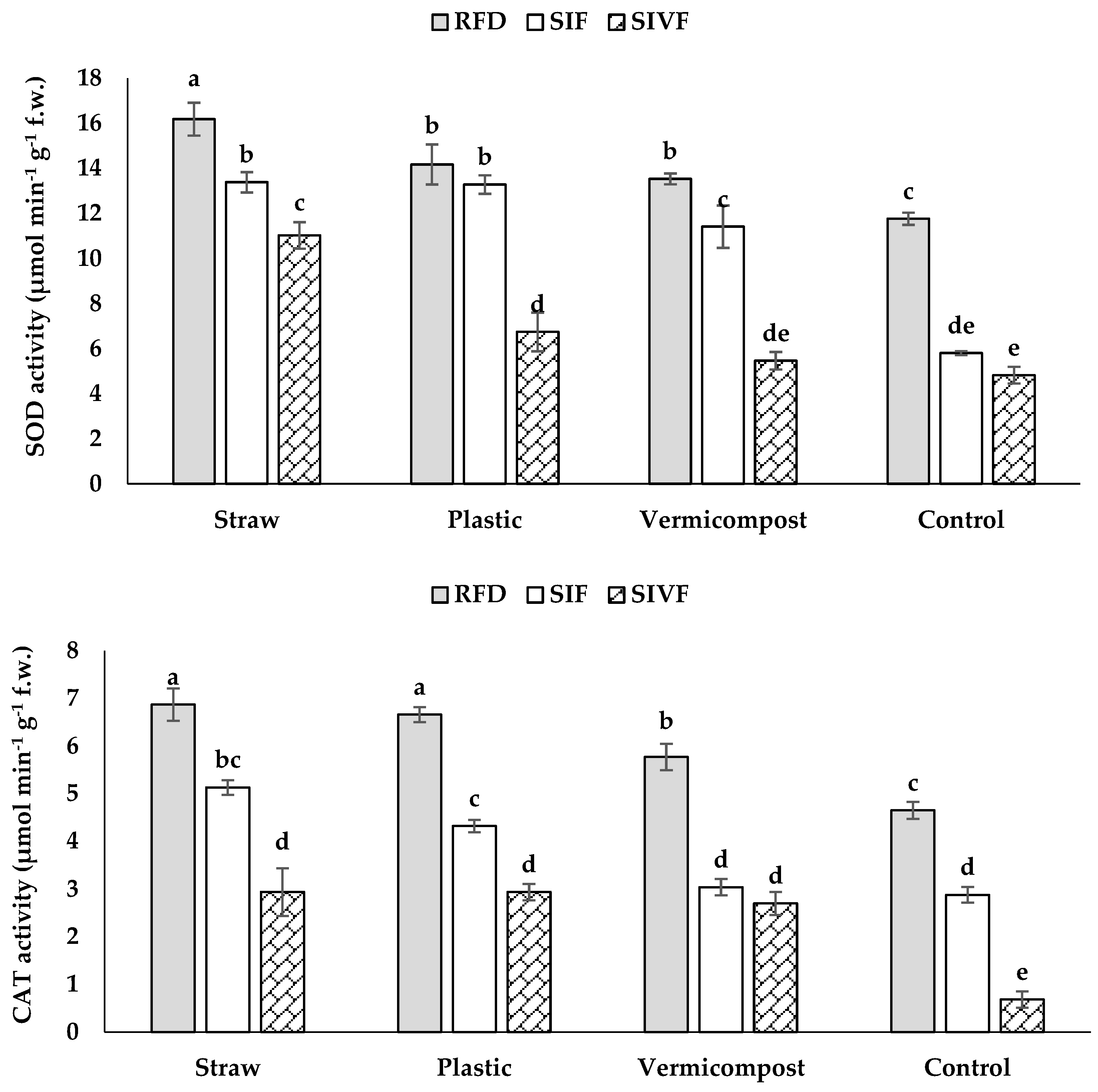
3.6. Antioxidant Enzyme Activity
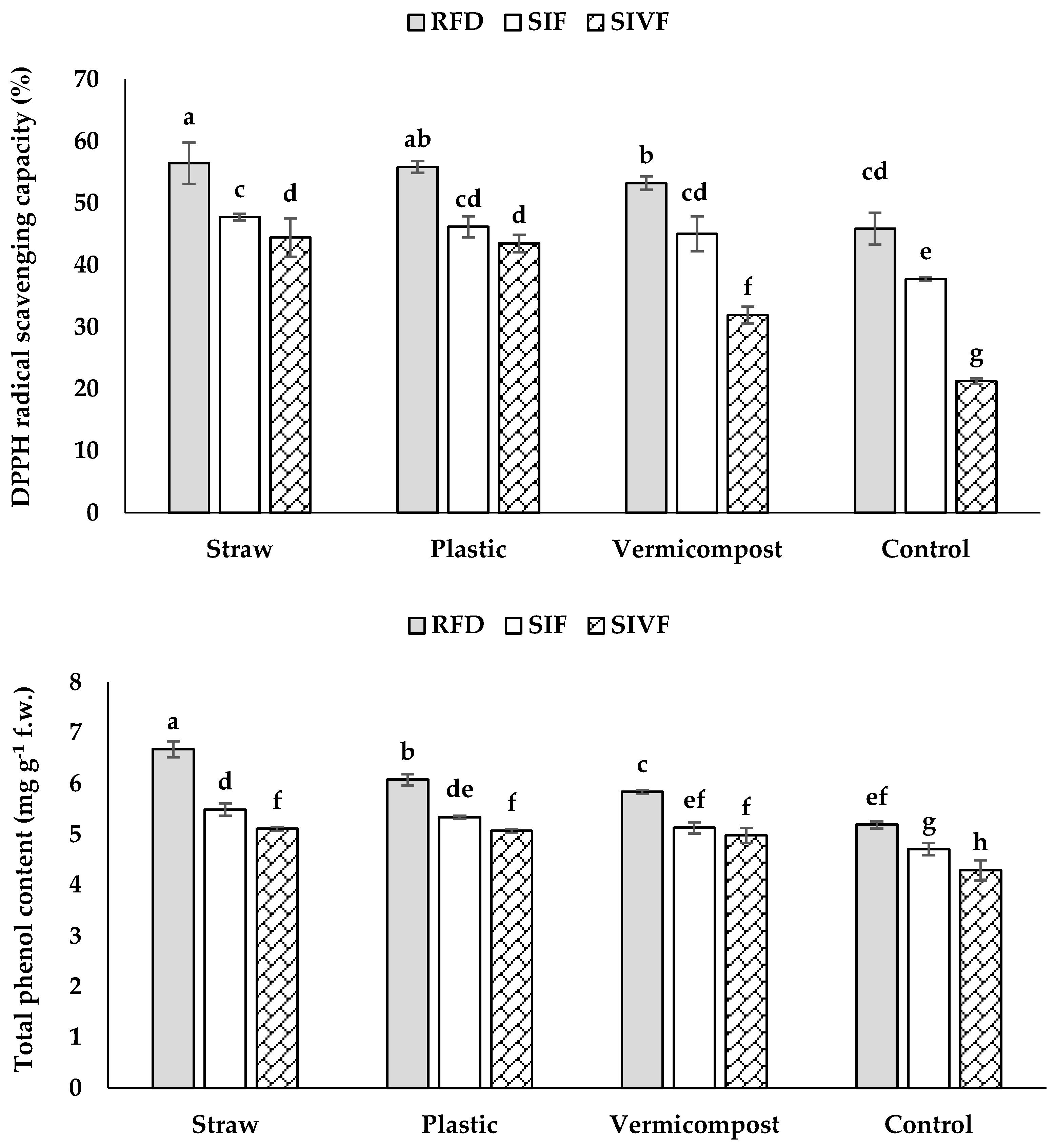
3.7. Total Phenol Content and DPPH Radical Scavenging Capacity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chugh, V.; Kaur, D.; Purwar, S.; Kaushik, P.; Sharma, V.; Kumar, H.; Rai, A.; Singh, C.M.; Kamaluddin Dubey, R.B. Applications of molecular markers for developing abiotic-stress-resilient oilseed crops. Life 2023, 13, 88. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Zou, Y.; Tian, R.; Huang, X.; Liu, L.; Wang, B.; Peng, D. Transcriptome analysis of fiber development under high-temperature stress in flax (Linum usitatissimum L.). Ind. Crops Prod. 2023, 195, 116019. [Google Scholar] [CrossRef]
- Paliwal, S.; Tripathi, M.K.; Tiwari, S.; Tripathi, N.; Payasi, D.K.; Tiwari, P.N.; Singh, K.; Yadav, R.K.; Asati, R.; Chauhan, S. Molecular advances to combat different biotic and abiotic stresses in linseed (Linum usitatissimum L.): A comprehensive review. Genes 2023, 14, 1461. [Google Scholar] [CrossRef]
- Hajibabaie, F.; Abedpoor, N.; Safavi, K.; Taghian, F. Natural remedies medicine derived from flaxseed (secoisolariciresinol diglucoside, lignans, and α-linolenic acid) improve network targeting efficiency of diabetic heart conditions based on computational chemistry techniques and pharmacophore modeling. J. Food Biochem. 2022, 46, 14480. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Qi, F.; Wang, F.; Lin, Y.; Xiaoyang, C.; Peng, Z.; Zhang, B.; Qi, X.; Deyholos, M.K.; Zhang, J. Evaluation of differentially expressed genes in leaves vs. roots subjected to drought stress in flax (Linum usitatissimum L.). Int. J. Mol. Sci. 2023, 24, 12019. [Google Scholar] [CrossRef]
- Behzadnejad, J.; Tahmasebi-Sarvestani, Z.; Aein, A.; Mokhtassi-Bidgoli, A. Wheat straw mulching helps improve yield in sesame (Sesamum indicum L.) under drought stress. Int. J. Plant Prod. 2020, 14, 389–400. [Google Scholar] [CrossRef]
- Lahmod, N.R.; Alkooranee, J.T.; Alshammary, A.A.G.; Rodrigo-Comino, J. Effect of wheat straw as a cover crop on the chlorophyll, seed, and oilseed yield of Trigonella foeunm graecum L under water deficiency and weed competition. Plants 2019, 8, 503. [Google Scholar] [CrossRef]
- Ahmad, S.; Zaheer, M.S.; Ali, H.H.; Erinle, K.O.; Wani, S.H.; Iqbal, R.; Okone, O.G.; Raza, A.; Waqas, M.M.; Nawaz, M. Physiological and biochemical properties of wheat (Triticum aestivum L.) under different mulching and water management systems in the semi-arid region of Punjab, Pakistan. Arid. Land Res. Manag. 2022, 36, 181–196. [Google Scholar] [CrossRef]
- Gholami, R.; Fahadi Hoveizeh, N.; Zahedi, S.M.; Arji, I. Effect of organic and synthetic mulches on some morpho-physiological and yield parameters of ‘Zard’ olive cultivar subjected to three irrigation levels in field conditions. S. Afr. J. Bot. 2023, 162, 749–760. [Google Scholar] [CrossRef]
- Rezaei-Chiyaneh, E.; Seyyedi, S.M.; Ebrahimian, E.; Moghaddam, S.S.; Damalas, C.A. Exogenous application of gamma-aminobutyric acid (GABA) alleviates the effect of water deficit stress in black cumin (Nigella sativa L.). Ind. Crops Prod. 2018, 112, 741–748. [Google Scholar] [CrossRef]
- Malik, A.; Shakir, A.S.; Khan, M.J.; Naveedullah, M.L.; Ajmal, M.; Ahmad, S. Effects of different mulching techniques on sugar beet performance under semi-arid subtropical climatic conditions. Pak. J. Bot. 2018, 50, 1219–1224. [Google Scholar]
- Close, D. Mulching for a Healthy Landscape; Virginia Tech Virginia State University: Petersburg, VI, USA, 2017; pp. 426–724. [Google Scholar]
- Bhardwaj, R.L. Effect of mulching on crop production under rainfed condition: A review. Agric. Rev. 2013, 34, 188–197. [Google Scholar] [CrossRef]
- Kumar, R.; Singh, A.; Sharma, R. Water conservation strategies in garlic cultivation. Irrig. Sci. 2022, 40, 765–776. [Google Scholar]
- Maughan, T.; Drost, D. Use of Plastic Mulch for Vegetable Production; Extension Utah State University: Logan, UT, USA, 2016. [Google Scholar]
- Mansouri, F.; Ahmad, B.; Rahman, M. Economic analysis of organic versus inorganic mulching in garlic production. J. Agric. Econ. 2020, 71, 299–311. [Google Scholar]
- Gao, Y.; Li, Y.; Wang, Y.; Wu, B.; Ke, J.; Niu, J.; Guo, L. Effect of different film color mulching on dry matter and grain yield of oil flax in dryland. Oil Crop Sci. 2020, 5, 17–22. [Google Scholar] [CrossRef]
- Ye, H.L.; Chen, Z.G.; Jia, T.T.; Su, Q.W.; Su, S.C. Response of different organic mulch treatments on yield and quality of Camellia oleifera. Agric. Water Manag. 2021, 245, 106654. [Google Scholar] [CrossRef]
- Rezaei-Chiyaneh, E.; Mahdavikia, H.; Hadi, H.; Muhittin, K.; Caruso, G.; Siddique, K.H.M. The effect of exogenously applied plant growth regulators and zinc on some physiological characteristics and essential oil constituents of Moldavian balm (Dracocephalum moldavica L.) under water stress. Physiol. Mol. Biol. Plants 2021, 27, 2201–2214. [Google Scholar] [CrossRef]
- Youssef, M.A.; AL-Huqail, A.A.; Ali, E.F.; Majrashi, A. Organic amendment and mulching enhanced the growth and fruit quality of squash plants (Cucurbita pepo L.) grown on silty loam soils. Horticulturae 2021, 7, 269. [Google Scholar] [CrossRef]
- Barreda, D.G.; Bautista, I.; Castell, V.; Lidón, A. Rice straw mulch installation in a vineyard improves weed control and modifies soil characteristics. Agronomy 2023, 13, 3068. [Google Scholar] [CrossRef]
- Paul, P.L.C.; Bell, R.W.; Barrett-Lennard, E.G.; Kabir, E. Straw mulch and irrigation affect solute potential and sunflower yield in a heavy textured soil in the Ganges Delta. Agric. Water Manag. 2020, 239, 106211. [Google Scholar] [CrossRef]
- Noor, M.A.; Nawaz, M.M.; Ma, W.; Zhao, M. Wheat straw mulch improves summer maize productivity and soil properties. Ital. J. Agron. 2020, 16, 1623. [Google Scholar] [CrossRef]
- Aziz, A.; Khan, A.; Asif, M.; Nadeem, A.M.; Ashraf, M.; Javaid, M.M.; Abbas, T.; Akhtar, N.; Munir, S. Impact of organic mulches on weed dynamics and productivity of rainfed wheat (Triticum aestivum L.). Pak. J. Weed Sci. Res. 2022, 28, 1–17. [Google Scholar] [CrossRef]
- Wang, Y.P.; Guo, L.Z.; Gao, Y.H.; Li, J.L.; Liu, H.S.; Xia, Z.X. Effects of biodegradable film mulching and combined application of organic and inorganic nitrogen fertilizer on dry matter accumulation and grain yield of oil flax. Oil Crop Sci. 2021, 6, 123–130. [Google Scholar]
- Liu, B.; Dai, Y.; Cheng, X.; He, X.; Bei, Q.; Wang, Y.; Zhou, Y.; Zhu, B.; Zhang, K.; Tian, X.; et al. Straw mulch improves soil carbon and nitrogen cycle by mediating microbial community structure and function in the maize field. Front. Microbiol. 2023, 14, 1217966. [Google Scholar] [CrossRef]
- Oyege, I.; Balaji Bhaskar, M.S. Effects of vermicompost on soil and plant health and promoting sustainable agriculture. Soil Syst. 2023, 7, 101. [Google Scholar] [CrossRef]
- Li, Y.; Xie, H.; Ren, Z.; Ding, Y.; Long, M.; Zhang, G.; Qin, X.; Siddique, K.H.; Liao, Y. Response of soil microbial community parameters to plastic film mulch: A meta-analysis. Geoderma 2022, 418, 115851. [Google Scholar] [CrossRef]
- Bandopadhyay, S.; Martin-Closas, L.; Pelacho, A.M.; DeBruyn, J.M. Biodegradable plastic mulch films: Impacts on soil microbial communities and ecosystem functions. Front. Microbiol. 2018, 9, 819. [Google Scholar] [CrossRef]
- Walker, W.R. Irrigation engineering—Sprinkler, trickle, surface irrigation principles, design and agricultural practices. Agric. Water Manag. 1984, 9, 263–264. [Google Scholar] [CrossRef]
- AOCS. Official Methods and Recommended Practices of the American Oil, 4th ed.; AOCS: Champaign, IL, USA, 1993; pp. 6–38. [Google Scholar]
- Zamani, F.; Amirnia, R.; Rezaei-Chiyaneh, E.; Gheshlaghi, M.; von Cossel, M.; Siddique, K.H.M. Optimizing essential oil, fatty acid profiles, and phenolic compounds of dragon’s head (Lallemantia iberica) intercropped with chickpea (Cicer arietinum L.) with biofertilizer inoculation under rainfed conditions in a semi-arid region. Arch. Agron. Soil Sci. 2022, 69, 1687–1704. [Google Scholar] [CrossRef]
- Taghizadeh, Y.; Amirnia, R.; Rezaei-Chiyaneh, E.; Ghiyasi, M.; Razavi, B.S.; Siddique, K.H.M. Co-Inoculation of mycorrhizal fungi with bacterial fertilizer along with intercropping scenarios improves seed yield and oil constituents of sesame. J. Soil Sci. Plant Nutr. 2023, 23, 2258–2272. [Google Scholar] [CrossRef]
- Saadat, B.; Pirzad, A.; Jalilian, J. Yield-related biochemical response of understory mycorrhizal yellow sweet clover (Melilotus officinalis L.) to drought in agrisilviculture. Arch. Agron. Soil Sci. 2020, 67, 1603–1620. [Google Scholar] [CrossRef]
- Lichtenthaler, H. Determination of total carotenoids and chlorophylls a and b of leaf in different solvents. Biochem. Soc. Trans. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Tejera, N.A.; Campos, R.; Sanjuan, J.; Lluch, C. Nitrogenase and antioxidant enzyme activities in Phaseolus vulgaris nodules formed by Rhizobium tropid isogenic strains with varying tolerance to salt stress. J. Plant Physiol. 2004, 161, 329–338. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Beyer, W.F.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Shin, Y.; Liu, R.H.; Nock, J.F.; Holliday, D.; Watkins, C.B. Temperature and relative humidity effects on quality, total ascorbic acid, phenolics and flavonoid concentrations, and antioxidant activity of strawberry. Postharvest Biol. Technol. 2007, 45, 349–357. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Rezaei-Chiyaneh, E.; Mahdavikia, H.; Alipour, H.; Dolatabadian, A.; Battaglia, L.M.; Harrison, T.M. Biostimulants alleviate water deficit stress and enhance essential oil productivity: A case study with savory. Sci. Rep. 2023, 13, 720. [Google Scholar] [CrossRef]
- Javaid, M.M.; Algwaiz, H.I.M.; Waheed, H.; Ashraf, M.; Mahmood, A.; Li, F.M.; Attia, K.A.; Nadeem, M.A.; AlKahtani, M.D.; Fiaz, S.; et al. Ridge-furrow mulching enhances capture and utilization of rainfall for improved maize production under rain-fed conditions. Agronomy 2022, 12, 1187. [Google Scholar] [CrossRef]
- Qu, Y.; Feng, B.L. Straw mulching improved yield of field buckwheat (Fagopyrum) by increasing water-temperature use and soil carbon in rain-fed farmland. Acta Ecol. Sin. 2022, 42, 11–16. [Google Scholar] [CrossRef]
- Chai, Y.; Chai, Q.; Yang, C.; Chen, Y.; Li, R.; Li, Y.; Chang, L.; Lan, X.; Cheng, H.; Chai, S. Plastic film mulching increases yield, water productivity, and net income of rain-fed winter wheat compared with no mulching in semiarid Northwest China. Agric. Water Manag. 2022, 262, 107420. [Google Scholar] [CrossRef]
- Wang, N.; Chen, H.; Ding, D.; Zhang, T.; Li, C.; Luo, X.; Chu, X.; Feng, H.; Wei, Y.; Siddique, K.H. Plastic film mulching affects field water balance components, grain yield, and water productivity of rainfed maize in the Loess Plateau, China: A synthetic analysis of multi-site observations. Agric. Water Manag. 2022, 266, 107570. [Google Scholar] [CrossRef]
- Zare, S.; Mirlohi, A.; Sabzalian, M.R.; Saeidi, G.; Koçak, M.Z.; Hano, C. Water stress and seed color interacting to impact seed and oil yield, protein, mucilage, and secoisolariciresinol diglucoside content in cultivated flax (Linum usitatissimum L.). Plants 2023, 12, 1632. [Google Scholar] [CrossRef]
- Cui, Z.; Yan, B.; Gao, Y.; Wu, B.; Wang, Y.; Wang, H.; Xu, P.; Zhao, B.; Cao, Z.; Zhang, Y.; et al. Agronomic cultivation measures on productivity of oilseed flax: A review. Oil Crop Sci. 2022, 7, 53–62. [Google Scholar] [CrossRef]
- Hasanvand, R.K.; Jalal-Abadi, A.L.; Abdali Mashhadi, A.; Kochekzadeh, A.; Siyahpoosh, A. The effect of different types of mulch and different cultivation methods on the quantitative and qualitative traits of sunflower (Helianthus annuus L.) in Ahvaz climate. J. Cent. Eur. Agric. 2023, 24, 115–125. [Google Scholar] [CrossRef]
- Ye H, Wen Y, Chen Z, Zhang T, Li S, Guan M, Zhang Y, Su S Relationship of soil microbiota to seed kernel metabolism in Camellia oleifera under mulched. Front. Plant Sci. 2022, 13, 920604. [CrossRef]
- Rahangdale, N.; Kumawat, N.; Jadav, M.L.; Bhagat, D.V.; Singh, M.; Yadav, R.K. Symbiotic efficiency, productivity and profitability of soybean as influenced by liquid bio-inoculants and straw mulch. Int. J. Bio-Resour. Stress Manag. 2022, 13, 9–16. [Google Scholar] [CrossRef]
- Kikhazaleh, M.; Ramroudi, M.; Galavi, M.; Ghanbari, S.A.; Fanaei, H.R. Effect of potassium on yield and some qualitative and physiological traits of safflower (Carthamus tinctorius L.) under drought stress conditions. J. Plant Nutr. 2023, 46, 2380–2392. [Google Scholar] [CrossRef]
- Chakraborty, D.; Nagarajan, S.; Aggarwal, P.; Gupta, V.K.; Tomar, R.K.; Garg, R.N.; Sahoo, R.N.; Sarkar, A.; Chopra, U.K.; Sarma, K.S.; et al. Effect of mulching on soil and plant water status, and the growth and yield of wheat (Triticum aestivum L.) in a semi-arid environment. Agric. Water Manag. 2008, 95, 1323–1334. [Google Scholar] [CrossRef]
- Gholami, R.; Zahedi, S.M. Effects of deficit irrigation and mulching on morpho-physiological and biochemical characteristics of konservolia olives. Gesunde Pflanz. 2020, 72, 49–55. [Google Scholar] [CrossRef]
- Abd El-Mageed, T.A.; El-Samnoudi, I.M.; Ibrahim, A.E.A.M.; Abd El Tawwab, A.R. Compost and mulching modulates morphological, physiological responses and water use efficiency in sorghum (bicolor L. Moench) under low moisture regime. Agric. Water Manag. 2018, 208, 431–439. [Google Scholar] [CrossRef]
- Maroof, T.; Tahir, M.; Zahid, N.; Khan, M.N.; Younas, M.T.; Shahzad, J.; Zafar, T.; Zeeshan, M.; Pervaiz, A.Y.A. Effect of different mulching materials on weed emergence and growth characteristics of strawberry under rainfed condition at Rawalakot Azad Jammu and Kashmir. J. Appl. Res. Plant Sci. 2024, 5, 46–57. [Google Scholar]
- Liao, Z.; Zeng, H.; Fan, J.; Lai, Z.; Zhang, C.; Zhang, F.; Wang, H.; Cheng, M.; Guo, J.; Li, Z.; et al. Effects of plant density, nitrogen rate and supplemental irrigation on photosynthesis, root growth, seed yield and water-nitrogen use efficiency of soybean under ridge-furrow plastic mulching. Agric. Water Manag. 2022, 268, 107688. [Google Scholar] [CrossRef]
- Zamani, F.; Amirnia, R.; Rezaei-Chiyaneh, E.; Gheshlaghi, M.; von Cossel, M.; Siddique, K.H.M. Alleviating plant water stress with biofertilizers: A case study for dragon’s head (Lallemantia iberica) and chickpea (Cicer arietinum L.) in a rainfed intercropping system. Int. J. Plant Prod. 2023, 17, 251–264. [Google Scholar] [CrossRef]
- Perveen, S.; Parvaiz, M.; Shahbaz, M.; Saeed, M.; Zafar, S. Triacontanol positively influences growth, yield, biochemical attributes and antioxidant enzymes of two linseed (Linum usitatissimum L.) accessions differing in drought tolerance. Pak. J. Bot. 2022, 54, 843–853. [Google Scholar] [CrossRef]
- Heydarzadeh, S.; Jalilian, J.; Pirzad, A.; Jamei, R.; Petrussa, E. Fodder value and physiological aspects of rainfed smooth vetch affected by biofertilizers and supplementary irrigation in an agri-silviculture system. Agrofor. Syst. 2022, 96, 221–232. [Google Scholar] [CrossRef]
- Heydarzadeh, S.; Arena, C.; Vitale, E.; Rahimi, A.; Mirzapour, M.; Nasar, J.; Kisaka, O.; Sow, S.; Ranjan, S.; Gitari, H. Impact of different fertilizer sources under supplemental irrigation and rainfed conditions on eco-physiological responses and yield characteristics of dragon’s head (Lallemantia iberica). Plants 2023, 12, 1693. [Google Scholar] [CrossRef]
- Hussain, S.; Wang, J.J.; Asad Naseer, M.; Saqib, M.; Siddiqui, M.H.; Ihsan, F.; Xiaoli, C.; Xiaolong, R.; Hussain, S.; Ramzan, H.N. Water stress memory in wheat/maize intercropping regulated photosynthetic and antioxidative responses under rainfed conditions. Sci. Rep. 2023, 13, 13688. [Google Scholar] [CrossRef]
- Sałata, A.; Nurzyńska-Wierdak, R.; Kalisz, A.; Moreno-Ramón, H. Impacts of alexandrian clover living mulch on the yield, phenolic content, and antioxidant capacity of leek and shallot. Agronomy 2022, 12, 2602. [Google Scholar] [CrossRef]
- Zheng, Z.; Zhou, W.; Yang, Q.; Jiang, J.; Miao, M. Effect of furrow straw mulching and straw decomposer application on celery (Apium graveolens L.) production and soil improvement. Agronomy 2023, 13, 2774. [Google Scholar] [CrossRef]
- Murariu, F.; Voda, A.D.; Murariu, O.C. Researches on food safety assessment—Supporting a healthy lifestyle for the population from NE of Romania. J. Biotechnol. 2019, 305, S68. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oct | Nov | Dec | Jan | Feb | Mar | Apr | May | June | July | Aug | Sep | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Monthly average temperature (°C) | 3.3 | 30.6 | 48.9 | 19.6 | 13.4 | 20.9 | 18.1 | 58.4 | 1.2 | 1.4 | 0.1 | 0.1 |
| Monthly average precipitation (mm) | 16.3 | 9.1 | 5.4 | 1.7 | 0.5 | 5.3 | 10.7 | 16.4 | 22.4 | 25.6 | 28.0 | 25.3 |
| Source of Variation | Plant Height | Branch Number | Follicle Number | Follicle Seed Number | 100-Seed Weight | Seed Yield | Biomass Yield | Oilseed Content | Oilseed Yield |
|---|---|---|---|---|---|---|---|---|---|
| Mulch (M) | ** | ** | ** | ** | ** | ** | ** | ** | ** |
| Irrigation condition (IC) | ** | * | ** | * | ** | * | * | ** | ** |
| M × IC | n.s. | n.s. | n.s. | * | ** | n.s. | n.s. | n.s. | * |
| No. | Component | Treatment | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RFD | RFD | RFD | RFD | SIF | SIF | SIF | SIF | SIVF | SIVF | SIVF | SIVF | ||
| C | V | PM | SM | C | V | PM | SM | C | V | PM | SM | ||
| 1 | Palmitic acid | 6.09 | 6.18 | 6.19 | 6.41 | 6.3 | 6.49 | 7.38 | 6.96 | 5.08 | 5.1 | 4.49 | 4.68 |
| 2 | Stearic acid | 5.08 | 5.29 | 5.78 | 5.53 | 5.61 | 5.67 | 5.5 | 5.66 | 5.15 | 5.09 | 4.97 | 5.45 |
| 3 | Oleic acid | 21.54 | 21.93 | 23.02 | 23.28 | 23.65 | 24.22 | 24.12 | 24.78 | 23.11 | 25.02 | 24.96 | 25.99 |
| 4 | Linoleic acid | 12.06 | 12.57 | 12.18 | 12.28 | 12.05 | 12.15 | 12.32 | 12.66 | 10.67 | 12.85 | 12.58 | 12.91 |
| 5 | Linolenic acid | 50.26 | 52.56 | 51.99 | 51.71 | 50.73 | 50.86 | 49.05 | 51.46 | 41.09 | 41.3 | 38.31 | 40.46 |
| 6 | Arachidic acid | 0.65 | 0.21 | 0.21 | 0.21 | 0.21 | 0.21 | 0.21 | 0.21 | 0.21 | 0.21 | 0.21 | 0.21 |
| Source of Variation | Relative Water Content | Chlorophyll a | Chlorophyll b | Carotenoids | Total Soluble Sugars | Proline Content | CAT | SOD | Total Phenol Content | DPPH Radical Scavenging Capacity |
|---|---|---|---|---|---|---|---|---|---|---|
| Mulch (M) | ** | ** | ** | * | ** | ** | ** | ** | ** | ** |
| Irrigation regime (IR) | * | * | n.s. | ** | ** | ** | ** | * | ** | ** |
| M × IR | n.s. | ** | * | ** | ** | ** | ** | ** | ** | ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moazzamnia, E.; Rezaei-Chiyaneh, E.; Dolatabadian, A.; Murariu, O.C.; Sannino, M.; Caruso, G.; Siddique, K.H.M. Effects of Water Stress and Mulch Type on Linseed Seed Yield, Physiological Traits, and Oil Compounds. Crops 2025, 5, 37. https://doi.org/10.3390/crops5030037
Moazzamnia E, Rezaei-Chiyaneh E, Dolatabadian A, Murariu OC, Sannino M, Caruso G, Siddique KHM. Effects of Water Stress and Mulch Type on Linseed Seed Yield, Physiological Traits, and Oil Compounds. Crops. 2025; 5(3):37. https://doi.org/10.3390/crops5030037
Chicago/Turabian StyleMoazzamnia, Elnaz, Esmaeil Rezaei-Chiyaneh, Aria Dolatabadian, Otilia Cristina Murariu, Maura Sannino, Gianluca Caruso, and Kadambot H. M. Siddique. 2025. "Effects of Water Stress and Mulch Type on Linseed Seed Yield, Physiological Traits, and Oil Compounds" Crops 5, no. 3: 37. https://doi.org/10.3390/crops5030037
APA StyleMoazzamnia, E., Rezaei-Chiyaneh, E., Dolatabadian, A., Murariu, O. C., Sannino, M., Caruso, G., & Siddique, K. H. M. (2025). Effects of Water Stress and Mulch Type on Linseed Seed Yield, Physiological Traits, and Oil Compounds. Crops, 5(3), 37. https://doi.org/10.3390/crops5030037