
Arsenic-Induced Oxidative Stress and Antioxidant Defense in Plants

 ,
,  , , , ,
, , , , 
Abstract
:1. Introduction
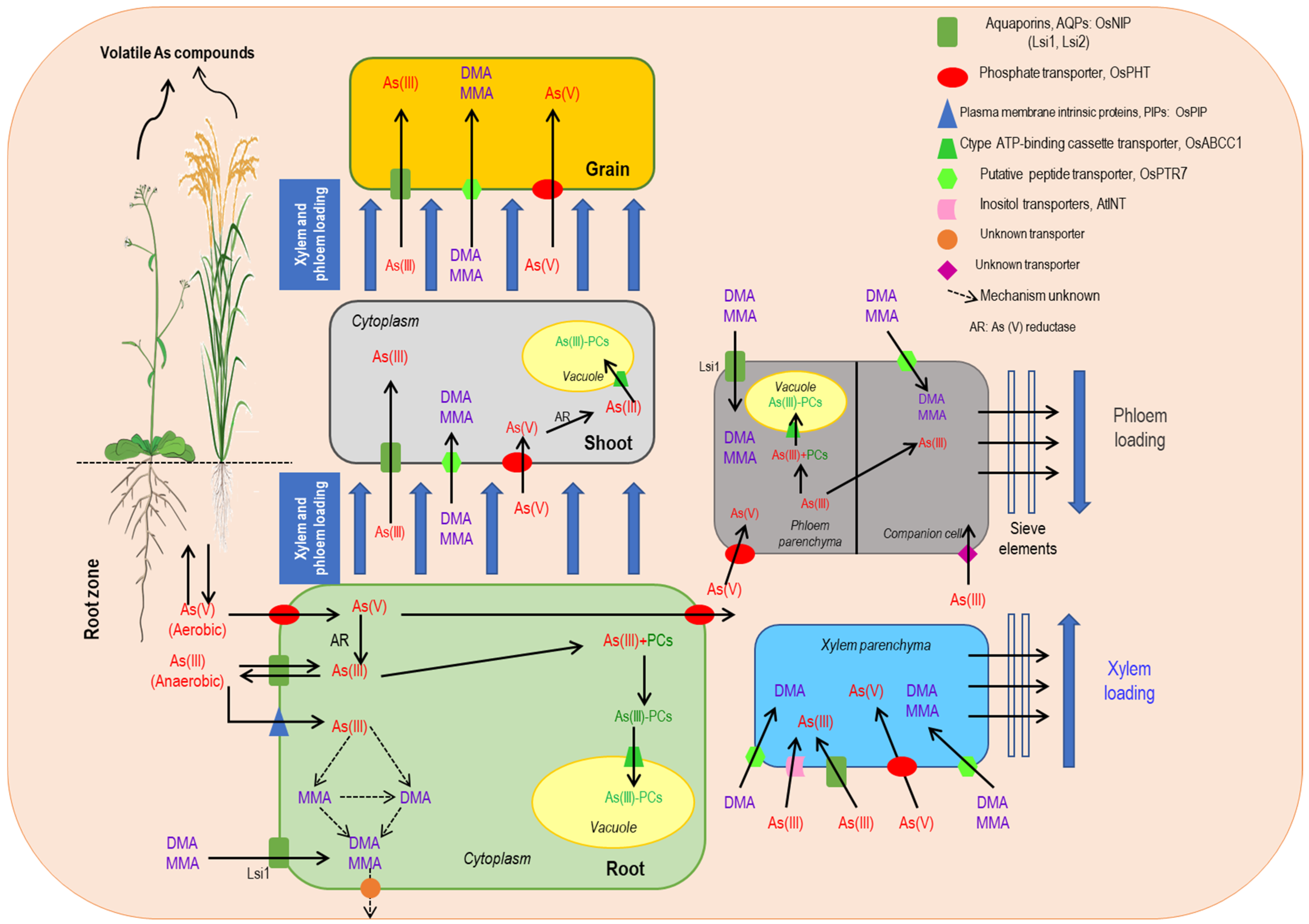
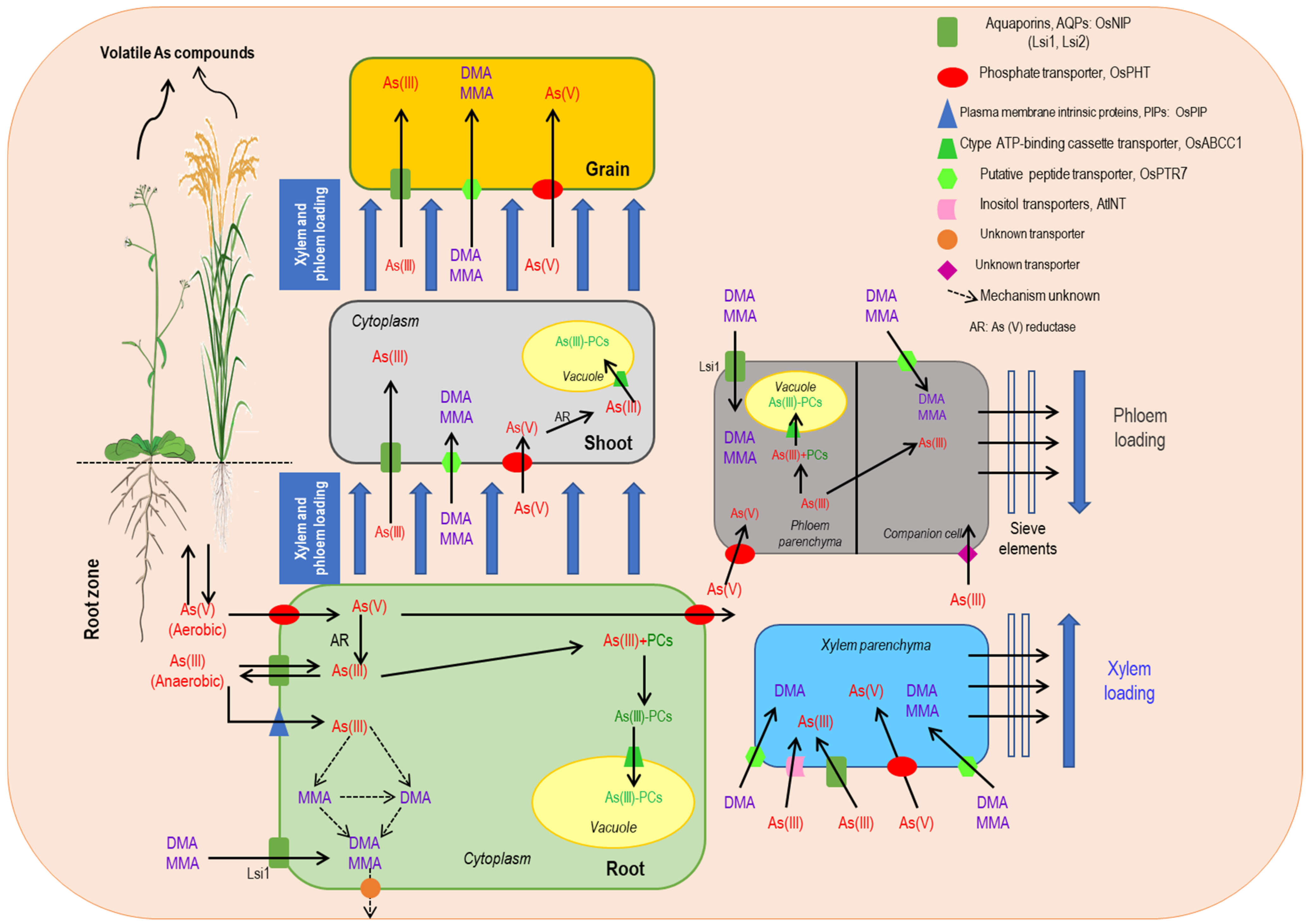
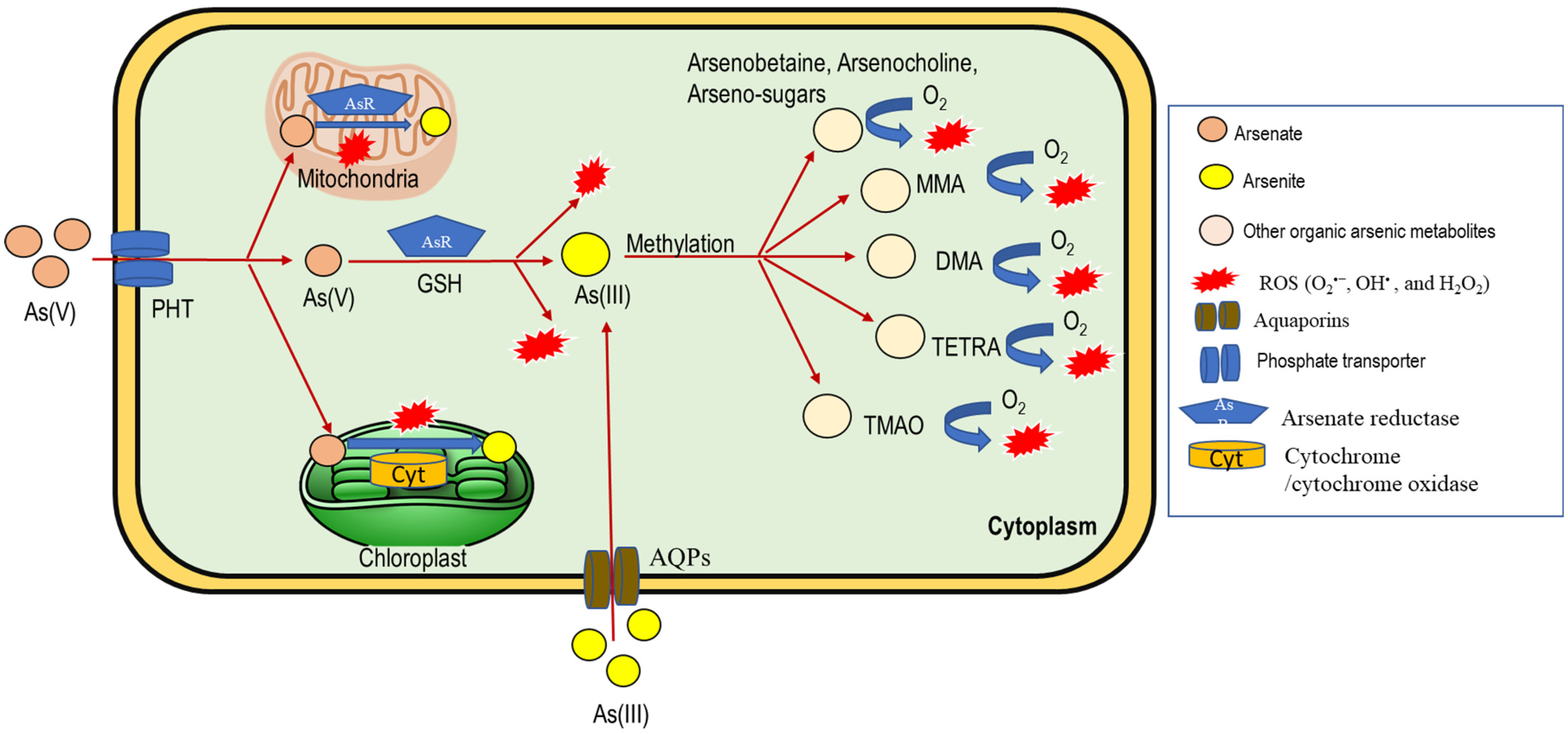
2. Arsenic Uptake and Accumulation in Plants
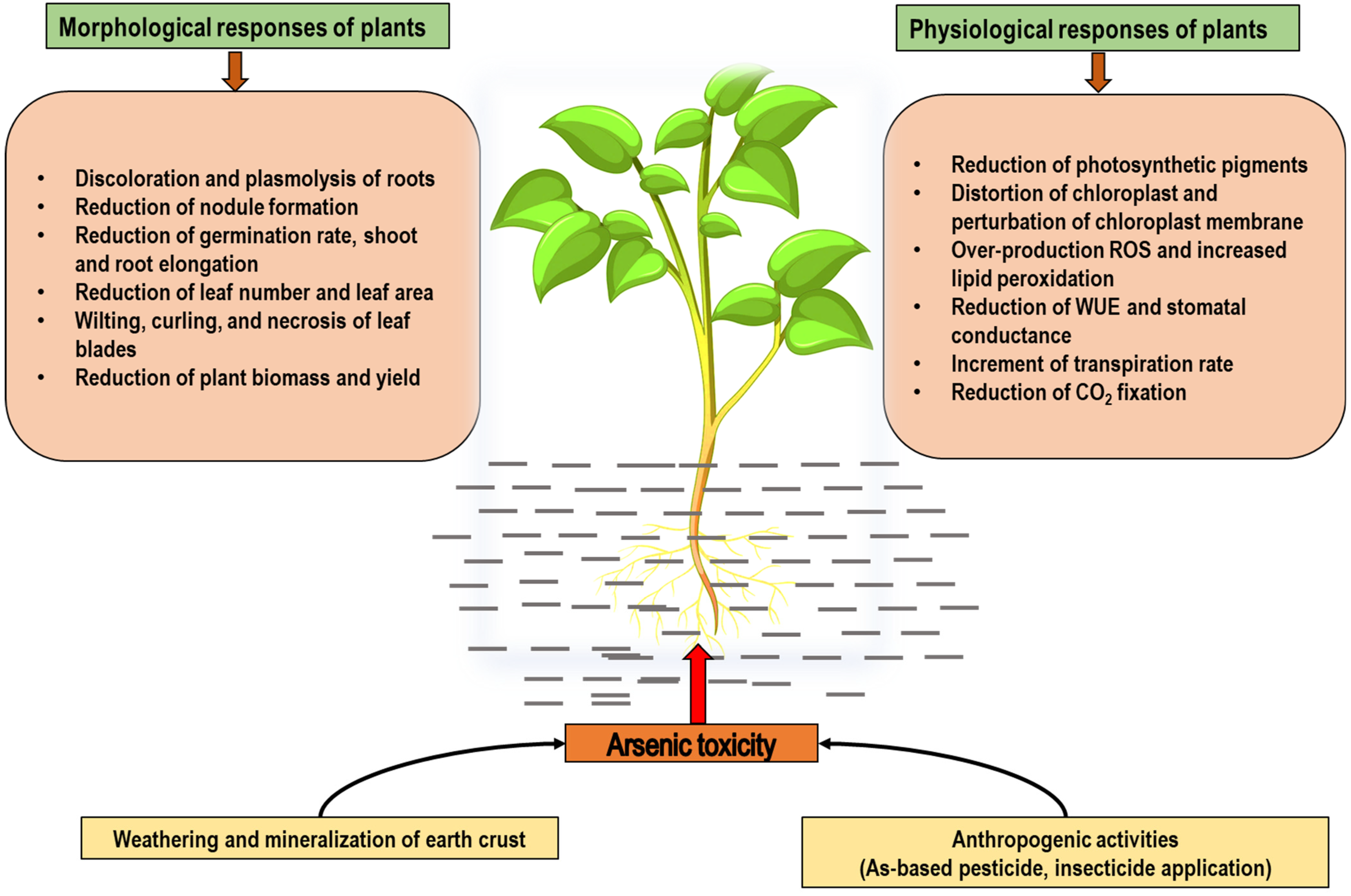
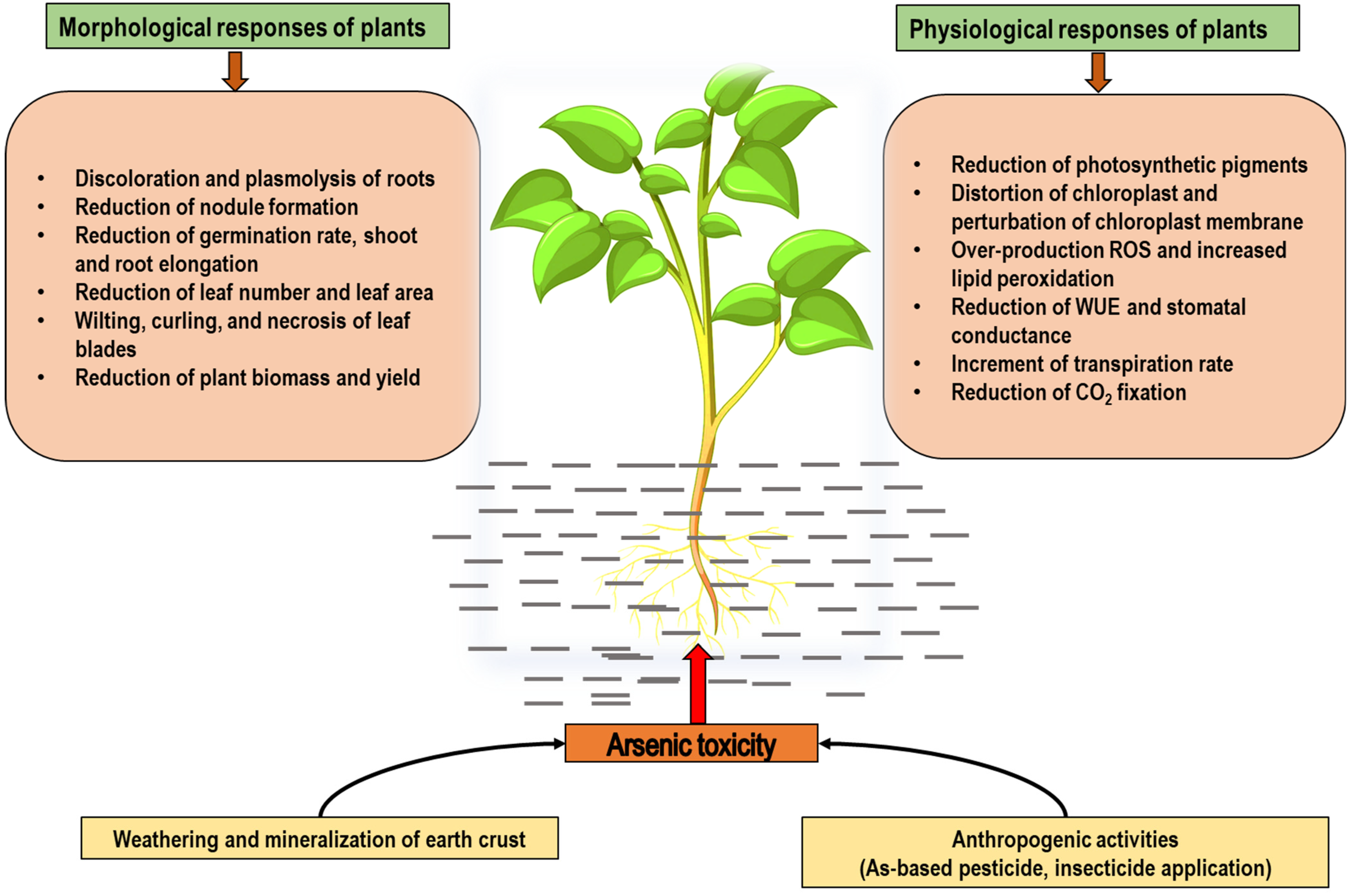
3. Arsenic Toxicity in Plants
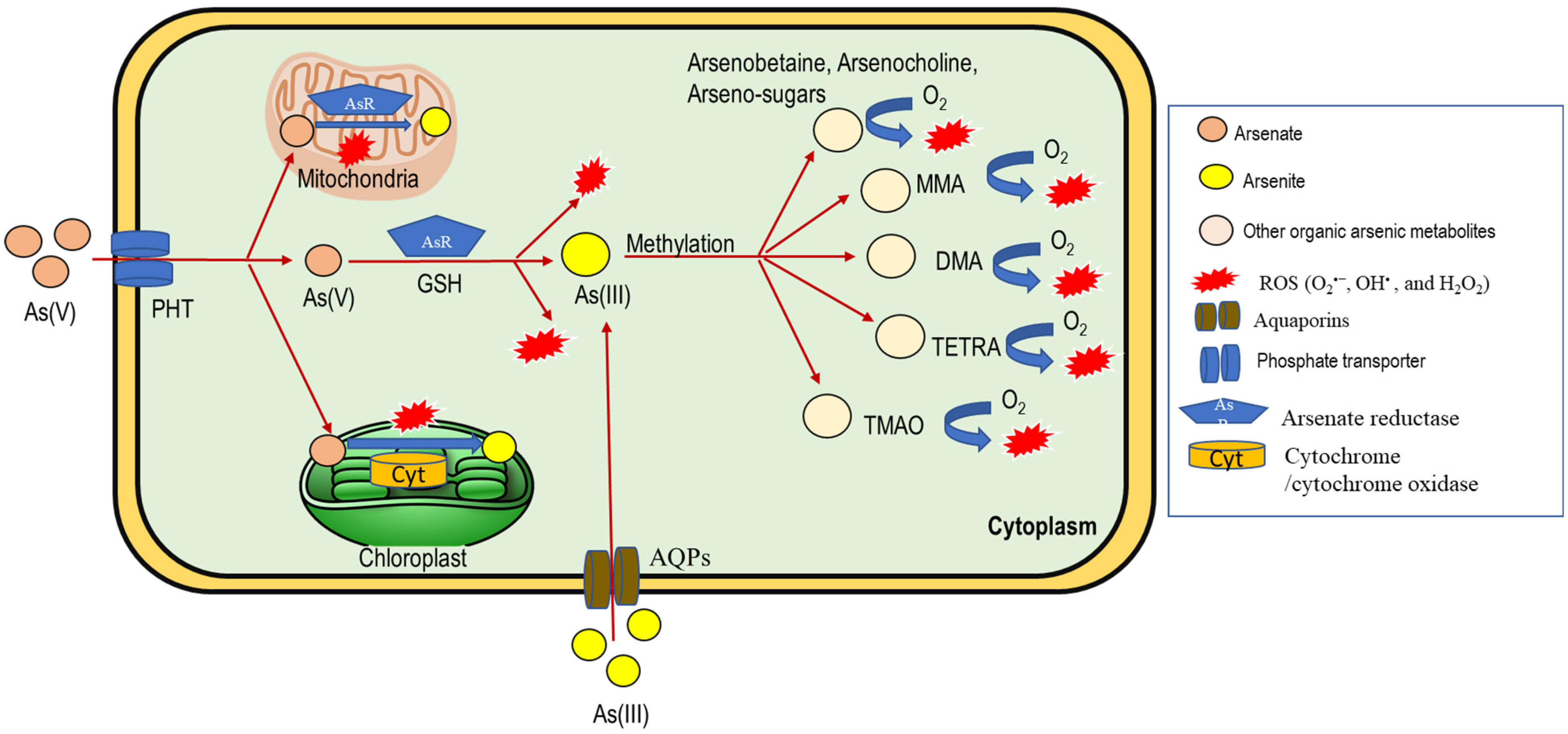
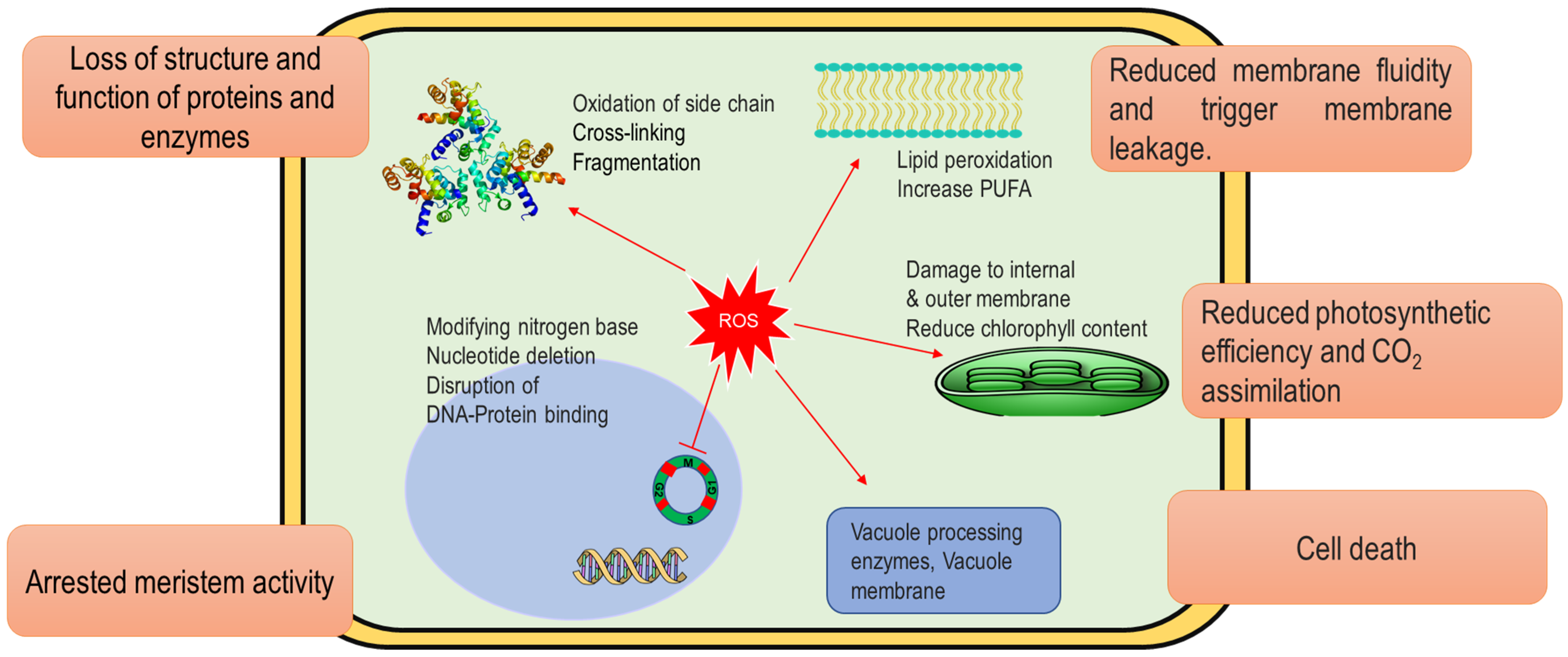
4. Arsenic Toxicity and ROS Generation in Plants
5. Arsenic-Induced Oxidative Stress in Plants
6. Antioxidants and Arsenic Tolerance
7. Approaches in Enhancing Oxidative Stress Tolerance in Plants Exposed to Arsenic
7.1. Use of Plant Nutrients
7.2. Use of Phytohormones
7.3. Use of Signaling Molecules
7.4. Use of Chelating Agents
7.5. Use of Soil Amendments
7.6. Use of Beneficial Microbes
7.7. Other Chemical Elicitors
8. Genetic Engineering in Enhancing Antioxidant Defense towards Arsenic Tolerance
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hasanuzzaman, M.; Nahar, K.; Fujita, M. Arsenic toxicity in plants and possible remediation. In Soil Remediation and Plants: Prospects and Challenges; Hakeem, K.R., Sabir, M., Öztürk, M., Mermut, A.R., Eds.; Academic Press: Amsterdam, The Netherlands, 2015; pp. 433–501. [Google Scholar]
- Chandrakar, V.; Pandey, N.; Keshavkant, S. Plant responses to arsenic toxicity: Morphology and physiology. In Mechanisms of Arsenic Toxicity and Tolerance in Plant; Hasanuzzaman, M., Nahar, K., Fujita, M., Eds.; Springer: Singapore, 2018; pp. 27–48. [Google Scholar]
- Polya, D.A.; Sparrenborn, C.; Datta, S.; Guo, H. Groundwater arsenic biogeochemistry: Key questions and use of tracers to understand arsenic-prone groundwater systems. Geosci. Front. 2019, 10, 1635–1641. [Google Scholar] [CrossRef]
- Finnegan, P.; Chen, W. Arsenic toxicity: The effects on plant metabolism. Front. Physiol. 2012, 3, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandrakar, V.; Naithani, S.C.; Keshavkant, S. Arsenic-induced metabolic disturbances and their mitigation mechanisms in crop plants: A review. Biologia 2016, 71, 367–377. [Google Scholar] [CrossRef]
- Kumar, V.; Vogelsang, L.; Schmidt, R.R.; Sharma, S.S.; Seidel, T.; Dietz, K.J. Remodeling of root growth under combined arsenic and hypoxia stress is linked to nutrient deprivation. Front. Plant Sci. 2020, 11, 569687. [Google Scholar] [CrossRef] [PubMed]
- Garg, N.; Singla, P. Arsenic toxicity in crop plants: Physiological effects and tolerance mechanisms. Environ. Chem. Lett. 2011, 9, 303–321. [Google Scholar] [CrossRef]
- Pandey, C.; Augustine, R.; Panthri, M.; Zia, I.; Bisht, N.C.; Gupta, M. Arsenic affects the production of glucosinolate, thiol and phytochemical compounds: A comparison of two Brassica cultivars. Plant Physiol. Biochem. 2017, 111, 144–154. [Google Scholar] [CrossRef]
- Ali, M.A.; Fahad, S.; Haider, I.; Ahmed, N.; Ahmad, S.; Hussain, S.; Arshad, M. Oxidative stress and antioxidant defense in plants exposed to metal/metalloid toxicity. In Reactive Oxygen, Nitrogen and Sulfur Species in Plants: Production, Metabolism, Signaling and Defense Mechanisms; Hasanuzzaman, M., Fotopoulos, V., Nahar, K., Fujita, M., Eds.; Wiley: Hoboken, NJ, USA, 2019; Volume 1, pp. 353–370. [Google Scholar]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Gupta, D.K.; Inouhe, M.; Rodríguez-Serrano, M.; Romero-Puerta, M.C.; Sandalio, L.M. Oxidative stress and arsenic toxicity: Role of NADPH oxidases. Chemosphere 2013, 90, 1987–1996. [Google Scholar] [CrossRef]
- Most, P.; Papenbrock, J. Possible roles of plant sulfurtransferases in detoxification of cyanide, reactive oxygen species, selected heavy metals and arsenate. Molecules 2015, 20, 1410–1423. [Google Scholar] [CrossRef]
- Singh, N.; Ma, L.Q.; Srivastava, M.; Rathinasabapathi, B. Metabolic adaptations to arsenic-induced oxidative stress in Pteris vittata L. and Pteris ensiformis L. Plant Sci. 2006, 170, 274–282. [Google Scholar] [CrossRef]
- Khatun, M.; Matsushima, D.; Rhaman, M.S.; Okuma, E.; Nakamura, T.; Nakamura, Y.; Munemasa, S.; Murata, Y. Exogenous proline enhances antioxidant enzyme activities but does not mitigate growth inhibition by selenate stress in tobacco BY-2 cells. Biosci. Biotechnol. Biochem. 2020, 84, 2281–2292. [Google Scholar] [CrossRef] [PubMed]
- Kaur, N.; Kaur, J.; Grewal, S.K.; Singh, I. Effect of heat stress on antioxidative defense system and its amelioration by heat acclimation and salicylic acid pre-treatments in three pigeonpea genotypes. Indian J. Agric. Biochem. 2019, 32, 106–110. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.; Anee, T.I.; Parvin, K.; Nahar, K.; Mahmud, J.A.; Fujita, M. Regulation of ascorbate-glutathione pathway in mitigating oxidative damage in plants under abiotic stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef] [Green Version]
- Kalita, J.; Pradhan, A.K.; Shandilya, Z.M.; Tanti, B. Arsenic stress responses and tolerance in rice: Physiological, cellular and molecular approaches. Rice Sci. 2018, 25, 235–249. [Google Scholar] [CrossRef]
- Shahid, M.; Khalid, S.; Bibi, I.; Khalid, S.; Masood, N.; Qaisrani, S.A.; Niazi, N.K.; Dumat, C. Arsenic-induced oxidative stress in Brassica oleracea: Multivariate and literature data analyses of physiological parameters, applied levels and plant organ type. Environ. Geochem. Health 2021, 1–13. [Google Scholar] [CrossRef]
- Abedi, T.; Mojiri, A. Arsenic uptake and accumulation mechanisms in rice species. Plants 2020, 9, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolterbeek, H.T.; van der Meer, A.J.G.M. Transport rate of arsenic, cadmium, copper and zinc in Potamogeton pectinatus L.: Radiotracer experiments with 76As, 109,115Cd, 64Cu and 65,69mZn. Sci. Total Environ. 2002, 287, 13–30. [Google Scholar] [CrossRef]
- Kumarathilaka, P.; Seneweera, S.; Meharg, A.; Bundschuh, J. Arsenic accumulation in rice (Oryza sativa L.) is influenced by environment and genetic factors. Sci. Total Environ. 2018, 642, 485–496. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.J.; Ago, Y.; Mitani, N.; Li, R.Y.; Su, Y.H.; Yamaji, N.; McGrath, S.P.; Ma, J.F. The role of the rice aquaporin Lsi1 in arsenite efflux from roots. New Phytol. 2010, 186, 392–399. [Google Scholar] [CrossRef]
- Sun, H.; Duan, Y.; Qi, X.; Zhang, L.; Huo, H.; Haijun Gong, H. Isolation and functional characterization of CsLsi2, a cucumber silicon efflux transporter gene. Ann. Bot. 2018, 122, 641–648. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Yan, H.; Chen, Y.; Shen, H.; Xu, W.; Zhang, H.; Shi, L.; Zhu, Y.G.; Ma, M. An aquaporin PvTIP4;1 from Pteris vittata may mediate arsenite uptake. New Phytol. 2016, 209, 746–761. [Google Scholar] [CrossRef] [PubMed]
- Kamiya, T.; Fujiwara, T. Arabidopsis NIP1;1 transports antimonite and determines antimonite sensitivity. Plant Cell Physiol. 2009, 50, 1977–1981. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Dai, W.; Yan, H.; Li, S.; Shen, H.; Chen, Y.; Xu, H.; Sun, Y.; He, Z.; Ma, M. Arabidopsis NIP3;1 plays an important role in arsenic uptake and root-to-shoot translocation under arsenite stress conditions. Mol. Plant 2015, 8, 722–733. [Google Scholar] [CrossRef] [Green Version]
- Mosa, K.A.; Kumar, K.; Chhikara, S.; Mcdermott, J.; Liu, Z.; Musante, C.; White, J.C.; Dhankher, O.P. Members of rice plasma membrane intrinsic proteins subfamily are involved in arsenite permeability and tolerance in plants. Transgenic Res. 2012, 21, 1265–1277. [Google Scholar] [CrossRef] [PubMed]
- Shri, M.; Singh, P.K.; Kidiwai, M.; Gautam, N.; Dubey, S.; Verma, G.; Chakrabarty, D. Recent advances in arsenic metabolism in plants: Current status, challenges and highlighted biotechnological intervention to reduce grain arsenic in rice. Metallomics 2019, 11, 519–532. [Google Scholar] [CrossRef]
- Song, W.Y.; Park, J.; Mendoza-Cózatl, D.G.; Suter-Grotemeyer, M.; Shim, D.; Hörtensteiner, S.; Geisler, M.; Weder, B.; Rea, P.A.; Rentsch, D. Arsenic tolerance in Arabidopsis is mediated by two ABCC-type phytochelatin transporters. Proc. Natl. Acad. Sci. USA 2010, 107, 21187–21192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Zhang, W.; Mao, C.; Xu, G.; Zhao, F.J. The role of OsPT8 in arsenate uptake and varietal difference in arsenate tolerance in rice. Environ. Exp. Bot. 2016, 67, 6051–6059. [Google Scholar] [CrossRef]
- Shi, S.; Wang, T.; Chen, Z.; Tang, Z.; Wu, Z.; Salt, D.E.; Chao, D.Y.; Zhao, F.J. OsHAC1; 1 and OsHAC1; 2 function as arsenate reductases and regulate arsenic accumulation. Plant Physiol. 2016, 172, 1708–1719. [Google Scholar] [CrossRef] [Green Version]
- Li, R.Y.; Ago, Y.; Liu, W.J.; Mitani, N.; Feldmann, J.; McGrath, S.P.; Ma, J.F.; Zhao, F.-J. The rice aquaporin Lsi1 mediates uptake of methylated arsenic species. Plant Physiol. 2009, 150, 2071–2080. [Google Scholar] [CrossRef] [Green Version]
- Mishra, S.; Mattusch, J.; Wennrich, R. Accumulation and transformation of inorganic and organic arsenic in rice and role of thiol-complexation to restrict their translocation to shoot. Sci. Rep. 2017, 7, 40522. [Google Scholar] [CrossRef]
- Ye, Y.; Li, P.; Xu, T.; Zeng, L.; Cheng, D.; Yang, M.; Luo, J.; Lian, X. OsPT4 contributes to arsenate uptake and transport in rice. Front. Plant Sci. 2017, 8, 2197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.; Sun, Z.; Li, Z.; Xue, R.; Cui, W.; Sun, S.; Liu, T.; Zeng, R.; Song, Y. Deficiency in silicon transporter Lsi1 compromises inducibility of anti-herbivore defense in rice plants. Front. Plant Sci. 2019, 10, 652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Y.; Sun, D.; Mei, H.; Liu, X.; Sun, S.; Xu, G.; Liu, Y.; Chen, Y. Knocking out OsPT4 gene decreases arsenate uptake by rice plants and inorganic arsenic accumulation in rice grains. Environ. Sci. Technol. 2017, 51, 12131–12138. [Google Scholar] [CrossRef] [PubMed]
- Carey, A.-M.; Scheckel, K.G.; Lombi, E.; Newville, M.; Choi, Y.; Norton, G.J.; Charnock, J.M.; Feldmann, J.; Price, A.H.; Meharg, A.A. Grain unloading of arsenic species in rice. Plant Physiol. 2010, 152, 309–319. [Google Scholar] [CrossRef] [Green Version]
- Mitra, A.; Chatterjee, S.; Moogouei, R.; Gupta, D.K. Arsenic accumulation in rice and probable mitigation approaches: A review. Agronomy 2017, 7, 67. [Google Scholar] [CrossRef]
- Tang, Z.; Chen, Y.; Chen, F.; Ji, Y.; Zhao, F.J. OsPTR7 (OsNPF8.1), a putative peptide transporter in rice, is involved in dimethylarsenate accumulation in rice grain. Plant Cell Physiol. 2017, 58, 904–913. [Google Scholar] [CrossRef]
- Abbas, G.; Murtaza, B.; Bibi, I.; Shahid, M.; Niazi, N.K.; Khan, M.I.; Amjad, M.; Hussain, M. Arsenic uptake, toxicity, detoxification, and speciation in plants: Physiological, biochemical, and molecular aspects. Int. J. Environ. Res. Public Health 2018, 15, 10059. [Google Scholar] [CrossRef] [Green Version]
- Kofroňová, M.; Hrdinová, A.; Mašková, P.; Tremlová, J.; Soudek, P.; Petrová, Š.; Pinkas, D.; Lipavská, H. Multi-component antioxidative system and robust carbohydrate status, the essence of plant arsenic tolerance. Antioxidants 2020, 9, 283. [Google Scholar] [CrossRef] [Green Version]
- Zemanová, V.; Pavlíková, D.; Hnilička, F.; Pavlík, M. Arsenic toxicity-induced physiological and metabolic changes in the shoots of Pteris cretica and Spinacia oleracea. Plants 2021, 10, 2009. [Google Scholar] [CrossRef]
- Martínez-Castillo, J.I.; Saldaña-Roblesa, A.; Ozuna, C. Arsenic stress in plants: A metabolomic perspective. Plant Stress 2022, 3, 100055. [Google Scholar] [CrossRef]
- Patel, H.V.; Parmar, S.R.; Chudasama, C.J.; Mangrola, A.V. Interactive studies of zinc with cadmium and arsenic on seed germination and antioxidant properties of Phaseolus aureus Roxb. Int. J. Plant Anim. Environ. Sci. 2013, 3, 166–174. [Google Scholar]
- Singh, R.; Jha, A.B.; Misra, A.N.; Sharma, P. Differential responses of growth, photosynthesis, oxidative stress, metals accumulation and NRAMP genes in contrasting Ricinus communis genotypes under arsenic stress. Environ. Sci. Pollut. Res. 2019, 26, 31166–31177. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Ruiz, M.; Aparicio-Chacón, M.V.; Palma, J.M.; Corpas, F.J. Arsenate disrupts ion balance, sulfur and nitric oxide metabolisms in roots and leaves of pea (Pisum sativum L.) plants. Environ. Exp. Bot. 2019, 161, 143–156. [Google Scholar] [CrossRef]
- Nath, S.; Panda, P.; Mishra, S.; Dey, M.; Choudhury, S.; Sahoo, L.; Panda, S.K. Arsenic stress in rice: Redox consequences and regulation by iron. Plant Physiol. Biochem. 2014, 80, 203–210. [Google Scholar] [CrossRef]
- Niazi, N.K.; Bibi, I.; Fatimah, A.; Shahid, M.; Javed, M.T.; Wang, H.; Ok, Y.S.; Bashir, S.; Murtaza, B.; Saqib, Z.A. Phosphate-assisted phytoremediation of arsenic by Brassica napus and Brassica juncea: Morphological and physiological response. Int. J. Phytoremediat. 2017, 19, 670–678. [Google Scholar] [CrossRef]
- Farooq, M.A.; Li, L.; Ali, B.; Gill, R.A.; Ali, B.; Wang, J.; Islam, F.; Ali, S.; Gill, M.B.; Zhou, W. Oxidative injury and antioxidant enzymes regulation in arsenic-exposed seedlings of four Brassica napus L. cultivars. Environ. Sci. Pollut. Res. 2015, 22, 10699–10712. [Google Scholar] [CrossRef]
- Mubarak, H.; Mirza, N.; Chai, L.-Y.; Yang, Z.-H.; Yong, W.; Tang, C.-J.; Mahmood, Q.; Pervez, A.; Farooq, U.; Fahad, S. Biochemical and metabolic changes in arsenic contaminated Boehmeria nivea L. BioMed Res. Int. 2016, 2016, 1423828. [Google Scholar] [CrossRef] [Green Version]
- Garg, N.; Singla, P. The role of Glomus mosseae on key physiological and biochemical parameters of pea plants grown in arsenic contaminated soil. Sci. Hortic. 2012, 143, 92–101. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Fujita, M. Exogenous sodium nitroprusside alleviates arsenic-induced oxidative stress in wheat (Triticum aestivum L.) seedlings by enhancing antioxidant defense and glyoxalase system. Ecotoxicology 2013, 22, 584–596. [Google Scholar] [CrossRef]
- Gusman, G.S.; Oliveira, J.A.; Farnese, F.S.; Cambraia, J. Arsenate and arsenite: The toxic effects on photosynthesis and growth of lettuce plants. Acta Physiol. Plant. 2013, 35, 1201–1209. [Google Scholar] [CrossRef]
- Srivastava, S.; Sharma, Y.K. Impact of arsenic toxicity on black gram and its amelioration using phosphate. Int. Sch. Res. Not. 2013, 2013, 340925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iriel, A.; Dundas, G.; Cirelli, A.F.; Lagorio, M.G. Effect of arsenic on reflectance spectra and chlorophyll fluorescence of aquatic plants. Chemosphere 2015, 119, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Duman, F.; Ozturk, F.; Aydin, Z. Biological responses of duckweed (Lemna minor L.) exposed to the inorganic arsenic species As(III) and As(V): Effects of concentration and duration of exposure. Ecotoxicology 2010, 19, 983–993. [Google Scholar] [CrossRef] [PubMed]
- Suneja, Y. Physio-Biochemical Responses and Allelic Diversity for Water Deficit Tolerance Related Traits in Aegilops tauschii and Triticum dicoccoides. Ph.D. Thesis, Punjab Agricultural University, Ludhiana, India, 2014. [Google Scholar]
- Emamverdian, A.; Ding, Y.; Mokhberdoran, F.; Xie, Y. Heavy metal stress and some mechanisms of plant defense response. Sci. World J. 2015, 2015, 756120. [Google Scholar] [CrossRef] [PubMed]
- Nabi, A.; Naeem, M.; Aftab, T.; Masroor, M.; Khan, A. Arsenic toxicity induced changes in growth, photosynthetic pigments, antioxidant machinery, essential oil, menthol and other active constituents of menthol mint (Mentha arvensis L.). J. Essent. Oil Bear. Plants 2019, 22, 1333–1348. [Google Scholar] [CrossRef]
- Bhattacharya, S.; De Sarkar, N.; Banerjee, P.; Banerjee, S.; Mukherjee, S.; Chattopadhyay, D.; Mukhopadhyay, A. Effects of arsenic toxicity on germination, seedling growth and peroxidase activity in Cicer arietinum. Int. J. Agric. Food Sci. 2012, 2, 131–137. [Google Scholar]
- Fatima, A.; Kataria, S.; Prajapati, R.; Jain, M.; Agrawal, A.K.; Singh, B.; Kashyap, Y.; Tripathi, D.K.; Singh, V.P.; Gadre, R. Magneto-priming effects on arsenic stress-induced morphological and physiological variations in soybean involving synchrotron imaging. Physiol. Plant. 2021, 173, 88–99. [Google Scholar]
- Farooq, M.A.; Islam, F.; Ali, B.; Najeeb, U.; Mao, B.; Gill, R.A.; Yan, G.; Siddique, K.H.M.; Zhou, W. Arsenic toxicity in plants: Cellular and molecular mechanisms of its transport and metabolism. Environ. Exp. Bot. 2016, 132, 42–52. [Google Scholar] [CrossRef]
- Gupta, P.; Bhatnagar, A.K. Spatial distribution of arsenic in different leaf tissues and its effect on structure and development of stomata and trichomes in mung bean, Vigna radiata (L.) Wilczek. Environ. Exp. Bot. 2015, 109, 12–22. [Google Scholar] [CrossRef]
- Armendariz, A.L.; Talano, M.A.; Villasuso, A.L.; Travaglia, C.; Racagni, G.E.; Reinoso, H.; Agostini, E. Arsenic stress induces changes in lipid signaling and evokes the stomata closure in soybean. Plant Physiol. Biochem. 2016, 103, 45–52. [Google Scholar] [CrossRef]
- De Oliveira, A.B.; Alencar, N.L.M.; Gomes-Filho, E. Comparison between the water and salt stress effects on plant growth and development. In Responses of Organisms to Water Stress; Akıncı, S., Ed.; Intech: London, UK, 2013. [Google Scholar] [CrossRef] [Green Version]
- Vezza, M.E.; Llanes, A.; Travaglia, C.; Agostini, E.; Talano, M.A. Arsenic stress effects on root water absorption in soybean plants: Physiological and morphological aspects. Plant Physiol. Biochem. 2018, 123, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Li, W.X.; Chen, T.B.; Huang, Z.C.; Lei, M.; Liao, X.Y. Effect of arsenic on chloroplast ultrastructure and calcium distribution in arsenic hyperaccumulator Pteris vittata L. Chemosphere 2006, 62, 803–809. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.; Labory, C.R.; Rangel, W.M.; Alves, E.; Guilherme, L.R. Anatomy and ultrastructure alterations of Leucaena leucocephala (Lam.) inoculated with mycorrhizal fungi in response to arsenic-contaminated soil. J. Hazard. Mater. 2013, 262, 1245–1258. [Google Scholar] [CrossRef] [PubMed]
- Upadhyaya, H.; Shome, S.; Roy, D.; Bhattacharya, M.K. Arsenic induced changes in growth and physiological responses in Vigna radiata seedling: Effect of curcumin interaction. Am. J. Plant Sci. 2014, 5, 3609–3618. [Google Scholar] [CrossRef] [Green Version]
- Farnese, F.S.; Oliveira, J.A.; Paiva, E.A.S.; Menezes-Silva, P.E.; da Silva, A.A.; Campos, F.V.; Ribeiro, C. The Involvement of nitric oxide in Integration of plant physiological and ultrastructural adjustments in response to arsenic. Front. Plant Sci. 2017, 8, 516. [Google Scholar] [CrossRef] [Green Version]
- García-Caparrós, P.; De Filippis, L.; Gul, A.; Hasanuzzaman, M.; Ozturk, M.; Altay, V.; Lao, M.T. Oxidative stress and antioxidant metabolism under adverse environmental conditions: A review. Bot. Rev. 2021, 87, 421–466. [Google Scholar] [CrossRef]
- Mishra, S.; Dwivedi, S.; Mallick, S.; Tripathi, R.D. Redox homeostasis in plants under arsenic stress. In Redox Homeostasis in Plants: From Signalling to Stress Tolerance; Panda, S.K., Yamamoto, Y.Y., Eds.; Springer: Singapore, 2019; pp. 179–198. [Google Scholar]
- Siddiqui, M.H.; Alamri, S.; Khan, M.N.; Corpas, F.J.; Al-Amri, A.A.; Alsubaie, Q.D.; Ali, H.M.; Kalaji, H.M.; Ahmad, P. Melatonin and calcium function synergistically to promote the resilience through ROS metabolism under arsenic-induced stress. J. Hazard. Mater. 2020, 398, 122882. [Google Scholar] [CrossRef]
- Coelho, D.G.; de Andrade, H.M.; Marinato, C.S.; Araujo, S.C.; de Matos, L.P.; da Silva, V.M.; de Oliveira, J.A. Exogenous jasmonic acid enhances oxidative protection of Lemna valdiviana subjected to arsenic. Acta Physiol. Plant. 2020, 42, 97. [Google Scholar] [CrossRef]
- Yadav, P.; Srivastava, S.; Patil, T.; Raghuvanshi, R.; Srivastava, A.K.; Suprasanna, P. Tracking the time-dependent and tissue-specific processes of arsenic accumulation and stress responses in rice (Oryza sativa L.). J. Hazard. Mater. 2021, 406, 124307. [Google Scholar] [CrossRef]
- Meharg, A.A.; Hartley-Whitaker, J. Arsenic uptake and metabolism in arsenic resistant and nonresistant plant species. New Phytol. 2002, 154, 29–43. [Google Scholar] [CrossRef] [Green Version]
- Meadows, R. How plants control arsenic accumulation. PLoS Biol. 2014, 12, e1002008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, I.; Wang, L.; Ollson, C.A.; Cullen, W.R.; Reimer, K.J. The predominance of inorganic arsenic species in plants from Yellowknife, northwest territories, Canada. Environ. Sci. Technol. 2000, 34, 22–26. [Google Scholar] [CrossRef]
- Delnomdedieu, M.; Basti, M.M.; Otvos, J.D.; Thomas, D.J. Reduction and binding of arsenate and dimethylarsinate by glutathione: A magnetic resonance study. Chem. Biol. Interact. 1994, 90, 139–155. [Google Scholar] [CrossRef]
- Ströher, E.; Grassl, J.; Carrie, C.; Fenske, R.; Whelan, J.; Millar, A.H. Glutaredoxin S15 is involved in Fe-S cluster transfer in mitochondria influencing lipoic acid-dependent enzymes, plant growth, and arsenic tolerance in Arabidopsis. Plant Physiol. 2016, 170, 1284–1299. [Google Scholar] [CrossRef] [Green Version]
- Van Breusegem, F.; Dat, J.F. Reactive oxygen species in plant cell death. Plant Physiol. 2006, 141, 384–390. [Google Scholar] [CrossRef] [Green Version]
- Talukdar, D. Arsenic-induced changes in growth and antioxidant metabolism of fenugreek. Russ. J. Plant Physiol. 2013, 60, 652–660. [Google Scholar] [CrossRef]
- Chandrakar, V.; Dubey, A.; Keshavkant, S. Modulation of antioxidant enzymes by salicylic acid in arsenic exposed Glycine max L. J. Soil Sci. Plant Nutr. 2016, 16, 662–676. [Google Scholar] [CrossRef] [Green Version]
- Chandrakar, V.; Yadu, B.; Meena, R.K.; Dubey, A.; Keshavkant, S. Arsenic-induced genotoxic responses and their amelioration by diphenylene iodonium, 24-epibrassinolide and proline in Glycine max L. Plant Physiol. Biochem. 2017, 112, 74–86. [Google Scholar] [CrossRef]
- Mucha, S.; Berezowski, M.; Markowska, K. Mechanisms of arsenic toxicity and transport in microorganisms. Adv. Microbiol. 2019, 56, 88–89. [Google Scholar] [CrossRef]
- Gupta, P.; Seth, C.S. Nitrate supplementation attenuates As(V) toxicity in Solanum lycopersicum L. cv Pusa Rohini: Insights into As(V) sub-cellular distribution, photosynthesis, nitrogen assimilation, and DNA damage. Plant Physiol. Biochem. 2019, 139, 44–55. [Google Scholar] [CrossRef]
- Dho, S.; Camusso, W.; Mucciarelli, M.; Fusconi, A. Arsenate toxicity on the apices of Pisum sativum L. seedling roots: Effects on mitotic activity, chromatin integrity and microtubules. Environ. Exp. Bot. 2010, 69, 17–23. [Google Scholar] [CrossRef]
- Tsukagoshi, H. Defective root growth triggered by oxidative stress is controlled through the expression of cell cycle-related genes. Plant Sci. 2012, 197, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.; Mishra, K.; Srivastava, S.; Kumar, A. Cytotoxic assessment of chromium and arsenic using chromosomal behavior of root meristem in Allium cepa L. Bull. Environ. Contam. Toxicol. 2018, 100, 803–808. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yue, H.; Xing, D. MAP Kinase 6-mediated activation of vacuolar processing enzyme modulates heat shock-induced programmed cell death in Arabidopsis. New Phytol. 2012, 195, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Piacentini, D.; Corpas, F.J.; D’Angeli, S.; Altamura, M.M.; Falasca, G. Cadmium and arsenic-induced-stress differentially modulates Arabidopsis root architecture, peroxisome distribution, enzymatic activities and their nitric oxide content. Plant Physiol. Biochem. 2020, 148, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Sharma, I. Arsenic induced oxidative stress in plants. Biologia 2012, 67, 447–453. [Google Scholar] [CrossRef]
- Devireddy, A.R.; Zandalinas, S.I.; Fichman, Y.; Mittler, R. Integration of reactive oxygen species and hormone signaling during abiotic stress. Plant J. 2021, 105, 459–476. [Google Scholar] [CrossRef]
- Gautam, A.; Pandey, A.K.; Dubey, R.S. Unravelling molecular mechanisms for enhancing arsenic tolerance in plants: A review. Plant Gene 2020, 23, 100240. [Google Scholar] [CrossRef]
- Islam, E.; Khan, M.T.; Irem, S. Biochemical mechanisms of signaling: Perspectives in plants under arsenic stress. Ecotoxicol. Environ. Saf. 2015, 114, 126–133. [Google Scholar] [CrossRef]
- Wang, P.; Chen, X.; Xu, X.; Lu, C.; Zhang, W.; Zhao, F.J. Arsenate induced chlorosis 1/TRANSLOCON at the outer envolope membrane of chloroplasts 132 protects chloroplasts from arsenic toxicity. Plant Physiol. 2018, 178, 1568–1583. [Google Scholar] [CrossRef] [Green Version]
- Asgher, M.; Ahmed, S.; Sehar, Z.; Gautam, H.; Gandhi, S.G.; Khan, N.A. Hydrogen peroxide modulates activity and expression of antioxidant enzymes and protects photosynthetic activity from arsenic damage in rice (Oryza sativa L.). J. Hazard. Mater. 2021, 401, 123365. [Google Scholar] [CrossRef] [PubMed]
- Arikan, B.; Ozfidan-Konakci, C.; Yildiztugay, E.; Zengin, G.; Alp, F.; Elbasan, F. Exogenous hesperidin and chlorogenic acid alleviate oxidative damage induced by arsenic toxicity in Zea mays through regulating the water status, antioxidant capacity, redox balance and fatty acid composition. Environ. Pollut. 2022, 292, 118389. [Google Scholar] [CrossRef] [PubMed]
- Alsahli, A.A.; Bhat, J.A.; Alyemeni, M.N.; Ashraf, M.; Ahmad, P. Hydrogen sulfide (H2S) mitigates arsenic (As)-induced toxicity in pea (Pisum sativum L.) plants by regulating osmoregulation, antioxidant defense system, ascorbate glutathione cycle and glyoxalase system. J. Plant Growth Regul. 2021, 40, 2515–2531. [Google Scholar] [CrossRef]
- Li, J.; Zhao, Q.; Xue, B.; Wu, H.; Song, G.; Zhang, X. Arsenic and nutrient absorption characteristics and antioxidant response in different leaves of two ryegrass (Lolium perenne) species under arsenic stress. PLoS ONE 2019, 14, e0225373. [Google Scholar] [CrossRef] [Green Version]
- Kaya, C.; Ashraf, M. Sodium hydrosulfite together with silicon detoxifies arsenic toxicity in tomato plants by modulating the AsA-GSH cycle. Environ. Pollut. 2022, 294, 118608. [Google Scholar] [CrossRef] [PubMed]
- Natasha; Shahid, M.; Khalid, S.; Saleem, M. Unrevealing arsenic and lead toxicity and antioxidant response in spinach: A human health perspective. Environ. Geochem. Health 2022, 44, 487–496. [Google Scholar] [CrossRef]
- Demircan, N.; Cucun, G.; Uzilday, B. Mitochondrial alternative oxidase (AOX1a) is required for the mitigation of arsenic-induced oxidative stress in Arabidopsis thaliana. Plant Biotechnol. Rep. 2020, 14, 235–245. [Google Scholar] [CrossRef]
- Farooq, M.A.; Zhang, K.; Islam, F.; Wang, J.; Athar, H.U.R.; Nawaz, A.; Ullah Zafar, Z.; Xu, J.; Zhou, W. Physiological and iTRAQ-based quantitative proteomics analysis of methyl jasmonate-induced tolerance in Brassica napus under arsenic stress. Proteomics 2018, 18, e1700290. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef] [Green Version]
- Rajput, V.D.; Harish; Singh, R.K.; Verma, K.K.; Sharma, L.; Quiroz-Figueroa, F.R.; Meena, M.; Gour, V.S.; Minkina, T.; Sushkova, S.; et al. Recent developments in enzymatic antioxidant defence mechanism in plants with special reference to abiotic stress. Biology 2021, 10, 267. [Google Scholar] [CrossRef]
- Singh, R.; Misra, A.N.; Sharma, P. Effect of arsenate toxicity on antioxidant enzymes and expression of nicotianamine synthase in contrasting genotypes of bioenergy crop Ricinus communis. Environ. Sci. Pollut. Res. Int. 2021, 28, 31421–31430. [Google Scholar] [CrossRef] [PubMed]
- Marques, D.N.; Stolze, S.C.; Harzen, A.; Nogueira, M.L.; Batagin-Piotto, K.D.; Piotto, F.A.; Mason, C.; Azevedo, R.A.; Nakagami, H. Comparative phosphoproteomic analysis of tomato genotypes with contrasting cadmium tolerance. Plant Cell Rep. 2021, 40, 2001–2008. [Google Scholar] [CrossRef] [PubMed]
- Ahsan, N.; Lee, D.G.; Alam, I.; Kim, P.J.; Lee, J.J.; Ahn, Y.O.; Kwak, S.S.; Lee, I.J.; Bahk, J.D.; Kang, K.Y.; et al. Comparative proteomic study of arsenic-induced differentially expressed proteins in rice roots reveals glutathione plays a central role during as stress. Proteomics 2008, 8, 3561–3576. [Google Scholar] [CrossRef] [PubMed]
- Amna, S.; Qamar, S.; Turab Naqvi, A.A.; Al-Huqail, A.A.; Qureshi, M.I. Role of sulfur in combating arsenic stress through upregulation of important proteins, and in-silico analysis to study the interaction between phosphate transporter (PHO1), arsenic and phosphate in spinach. Plant Physiol. Biochem. 2020, 157, 348–358. [Google Scholar] [CrossRef]
- Chauhan, R.; Awasthi, S.; Indoliya, Y.; Chauhan, A.S.; Mishra, S.; Agrawal, L.; Srivastava, S.; Dwivedi, S.; Singh, P.C.; Mallick, S.; et al. Transcriptome and proteome analyses reveal selenium mediated amelioration of arsenic toxicity in rice (Oryza sativa L.). J. Hazard. Mater. 2020, 390, 122122. [Google Scholar] [CrossRef]
- Dixit, G.; Singh, A.P.; Kumar, A.; Dwivedi, S.; Deeba, F.; Kumar, S.; Suman, S.; Adhikari, B.; Shukla, Y.; Trivedi, P.K.; et al. Sulfur alleviates arsenic toxicity by reducing its accumulation and modulating proteome, amino acids and thiol metabolism in rice leaves. Sci. Rep. 2015, 5, 16205. [Google Scholar] [CrossRef] [Green Version]
- Farooq, M.A.; Hong, Z.; Islam, F.; Noor, Y.; Hannan, F.; Zhang, Y.; Ayyaz, A.; Mwamba, T.M.; Zhou, W.; Song, W. Comprehensive proteomic analysis of arsenic induced toxicity reveals the mechanism of multilevel coordination of efficient defense and energy metabolism in two Brassica napus cultivars. Ecotoxicol. Environ. Saf. 2021, 208, 111744. [Google Scholar] [CrossRef]
- Liu, Y.; Damaris, R.N.; Yang, P. Proteomics analysis identified a DRT protein involved in arsenic resistance in Populus. Plant Cell Rep. 2017, 36, 1855–1869. [Google Scholar] [CrossRef]
- Rai, R.; Pandey, S.; Shrivastava, A.K.; Rai, S.P. Enhanced photosynthesis and carbon metabolism favor arsenic tolerance in Artemisia annua, a medicinal plant as revealed by homology-based proteomics. Int. J. Proteom. 2014, 2014, 163962. [Google Scholar] [CrossRef] [Green Version]
- Requejo, R.; Tena, M. Proteome analysis of maize roots reveals that oxidative stress is a main contributing factor to plant arsenic toxicity. Phytochemistry 2005, 66, 1519–1528. [Google Scholar] [CrossRef]
- Marques, D.N.; Carvalho, M.E.A.; Piotto, F.A.; Batagin-Piotto, K.D.; Nogueira, M.L.; Gaziola, S.A.; Azevedo, R.A. Antioxidant defense response in plants to cadmium stress. In Cadmium Tolerance in Plants: Agronomic, Molecular, Signaling, and Omic Approaches; Hasanuzzaman, M., Narasimha, M., Prasad, M.N.V., Nahar, K., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 423–461. [Google Scholar]
- Marques, D.N.; Gaziola, S.A.; Azevedo, R.A. Phytochelatins and their relationship with modulation of cadmium tolerance in plants. In Handbook of Bioremediation: Physiological, Molecular and Biotechnological Interventions; Hasanuzzaman, M., Prasad, M.N.V., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 91–113. [Google Scholar]
- Soares, C.; Carvalho, M.E.A.; Azevedo, R.A.; Fidalgo, F. Plants facing oxidative challenges-a little help from the antioxidant networks. Environ. Exp. Bot. 2019, 161, 4–25. [Google Scholar] [CrossRef]
- Tripathi, R.D.; Tripathi, P.; Dwivedi, S.; Dubey, S.; Chatterjee, S.; Chakrabarty, D.; Trivedi, P.K. Arsenomics: Omics of arsenic metabolism in plants. Front. Physiol. 2012, 3, 275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, P.; Tripathi, R.D. Metabolome modulation during arsenic stress in plants. In Plant-Metal Interactions; Srivastava, S., Srivastava, A., Suprasanna, P., Eds.; Springer: Singapore, 2019; pp. 119–140. [Google Scholar]
- Shri, M.; Kumar, S.; Chakrabarty, D.; Trivedi, P.K.; Mallick, S.; Misra, P.; Shukla, D.; Mishra, S.; Srivastava, S.; Tripathi, R.D.; et al. Effect of arsenic on growth, oxidative stress, and antioxidant system in rice seedlings. Ecotoxicol. Environ. Saf. 2009, 72, 1102–1110. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, F.; Tandon, P.K.; Srivastava, S. Arsenite and arsenate impact the oxidative status and antioxidant responses in Ocimum tenuiflorum L. Physiol. Mol. Biol. Plants 2015, 21, 453–458. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, F.; Tandon, P.K.; Srivastava, S. Analysis of arsenic induced physiological and biochemical responses in a medicinal plant, Withania somnifera. Physiol. Mol. Biol. Plants 2015, 21, 61–69. [Google Scholar] [CrossRef] [Green Version]
- Saha, I.; Hasanuzzaman, M.; Adak, M.K. Abscisic acid priming regulates arsenite toxicity in two contrasting rice (Oryza sativa L.) genotypes through differential functioning of sub1A quantitative trait loci. Environ. Pollut. 2021, 287, 117586. [Google Scholar] [CrossRef]
- Bianucci, E.; Furlan, A.; Tordable, M.D.C.; Hernández, L.E.; Carpena-Ruiz, R.O.; Castro, S. Antioxidant responses of peanut roots exposed to realistic groundwater doses of arsenate: Identification of glutathione S-transferase as a suitable biomarker for metalloid toxicity. Chemosphere 2017, 181, 551–561. [Google Scholar] [CrossRef] [Green Version]
- Leão, G.A.; Oliveira, J.A.; Felipe, R.T.A.; Farnese, F.S.; Gusman, G.S. Anthocyanins, thiols, and antioxidant scavenging enzymes are involved in Lemna gibba tolerance to arsenic. J. Plant Interact. 2013, 9, 143–151. [Google Scholar] [CrossRef]
- Mishra, S.; Jha, A.B.; Dubey, R.S. Arsenite treatment induces oxidative stress, upregulates antioxidant system, and causes phytochelatin synthesis in rice seedlings. Protoplasma 2011, 248, 565–577. [Google Scholar] [CrossRef]
- Leão, G.A.; Oliveia, J.A.; Felipe, R.T.A.; Farnese, F.S. Phytoremediation of arsenic-contaminated water: The role of antioxidant metabolism of Azolla caroliniana Willd. (Salviniales). Acta Bot. Bras. 2017, 31, 161–168. [Google Scholar] [CrossRef] [Green Version]
- Saha, J.; Majumder, B.; Mumtaz, B.; Biswas, A.K. Arsenic-induced oxidative stress and thiol metabolism in two cultivars of rice and its possible reversal by phosphate. Acta Physiol. Plant 2017, 39, 263. [Google Scholar] [CrossRef]
- Anjum, S.A.; Tanveer, M.; Hussain, S.; Shahzad, B.; Ashraf, U.; Fahad, S.; Hassan, W.; Jan, S.; Khan, I.; Saleem, M.F.; et al. Osmoregulation and antioxidant production in maize under combined cadmium and arsenic stress. Environ. Sci. Pollut. Res. Int. 2016, 23, 11864–11875. [Google Scholar] [CrossRef] [PubMed]
- Dave, R.; Tripathi, R.D.; Dwivedi, S.; Tripathi, P.; Dixit, G.; Sharma, Y.K.; Trivedi, P.K.; Corpas, F.J.; Barroso, J.B.; Chakrabarty, D. Arsenate and arsenite exposure modulate antioxidants and amino acids in contrasting arsenic accumulating rice (Oryza sativa L.) genotypes. J. Hazard. Mater. 2013, 262, 1123–1131. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.; Ahmad, M.A. Arsenate induced differential response in rice genotypes. Ecotoxicol. Environ. Saf. 2014, 107, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Majumder, B.; Das, S.; Mukhopadhyay, S.; Biswas, A.K. Identification of arsenic-tolerant and arsenic-sensitive rice (Oryza sativa L.) cultivars on the basis of arsenic accumulation assisted stress perception, morpho-biochemical responses, and alteration in genomic template stability. Protoplasma 2019, 256, 193–211. [Google Scholar] [CrossRef] [PubMed]
- Majumder, B.; Das, S.; Pal, B.; Biswas, A.K. Evaluation of arsenic induced toxicity based on arsenic accumulation, translocation and its implications on physio-chemical changes and genomic instability in indica rice (Oryza sativa L.) cultivars. Ecotoxicol. Environ. Saf. 2020, 29, 13–34. [Google Scholar] [CrossRef]
- Mallick, S.; Sinam, G.; Sinha, S. Study on arsenate tolerant and sensitive cultivars of Zea mays L.: Differential detoxification mechanism and effect on nutrients status. Ecotoxicol. Environ. Saf. 2011, 74, 1316–1324. [Google Scholar] [CrossRef]
- Mallick, S.; Kumar, N.; Sinha, S.; Dubey, A.K.; Tripathi, R.D.; Srivastav, V. H2O2 pretreated rice seedlings specifically reduces arsenate not arsenite: Difference in nutrient uptake and antioxidant defense response in a contrasting pair of rice cultivars. Physiol. Mol. Biol. Plants 2014, 20, 435–447. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, S.; Srivastava, A.K.; Suprasanna, P.; D’Souza, S.F. Comparative biochemical and transcriptional profiling of two contrasting varieties of Brassica juncea L. in response to arsenic exposure reveals mechanisms of stress perception and tolerance. J. Exp. Bot. 2009, 60, 3419–3431. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, S.; Srivastava, A.K.; Suprasanna, P.; D’Souza, S.F. Comparative antioxidant profiling of tolerant and sensitive varieties of Brassica juncea L. to arsenate and arsenite exposure. Bull. Environ. Contam. Toxicol. 2010, 84, 342–346. [Google Scholar] [CrossRef]
- Tripathi, P.; Singh, R.P.; Sharma, Y.K.; Tripathi, R.D. Arsenite stress variably stimulates pro-oxidant enzymes, anatomical deformities, photosynthetic pigment reduction, and antioxidants in arsenic-tolerant and sensitive rice seedlings. Environ. Toxicol. Chem. 2015, 34, 1562–1571. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Upadhyay, A.K.; Singh, D.P. Regulation of oxidative stress and mineral nutrient status by selenium in arsenic treated crop plant Oryza sativa. Ecotoxicol. Environ. Saf. 2018, 148, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, D.K.; Rai, P.; Guerriero, G.; Sharma, S.; Corpas, F.J.; Singh, V.P. Silicon induces adventitious root formation in rice under arsenate stress with involvement of nitric oxide and indole-3-acetic acid. J. Exp. Bot. 2021, 72, 4457–4471. [Google Scholar] [CrossRef] [PubMed]
- Surgun-Acar, Y.; Zemheri-Navruz, F. 24-Epibrassinolide promotes arsenic tolerance in Arabidopsis thaliana L. by altering stress responses at biochemical and molecular level. J. Plant Physiol. 2019, 238, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Kaya, C.; Ashraf, M.; Alyemeni, M.N.; Corpas, F.J.; Ahmad, P. Salicylic acid-induced nitric oxide enhances arsenic toxicity tolerance in maize plants by upregulating the ascorbate-glutathione cycle and glyoxalase system. J. Hazard. Mater. 2020, 399, 123020. [Google Scholar] [CrossRef]
- Singh, A.P.; Dixit, G.; Kumar, A.; Mishra, S.; Kumar, N.; Dixit, S.; Singh, P.K.; Dwivedi, S.; Trivedi, P.K.; Pandey, V.; et al. A protective role for nitric oxide and salicylic acid for arsenite phytotoxicity in rice (Oryza sativa L.). Plant Physiol. Biochem. 2017, 115, 163–173. [Google Scholar] [CrossRef] [Green Version]
- Farooq, M.A.; Islam, F.; Yang, C.; Nawaz, A.; Athar, H.R.; Gill, R.A.; Ali, B.; Song, W.; Zhou, W. Methyl jasmonate alleviates arsenic-induced oxidative damage and modulates the ascorbate–glutathione cycle in oilseed rape roots. J. Plant Growth Regul. 2018, 84, 135–148. [Google Scholar] [CrossRef]
- Praveen, A.; Gupta, M. Nitric oxide confronts arsenic stimulated oxidative stress and root architecture through distinct gene expression of auxin transporters, nutrient related genes and modulates biochemical responses in Oryza sativa L. Environ. Pollut. 2019, 240, 950–962. [Google Scholar] [CrossRef]
- Praveen, A.; Pandey, A.; Gupta, M. Nitric oxide alters nitrogen metabolism and PIN gene expressions by playing protective role in arsenic challenged Brassica juncea L. Ecotoxiol. Environ. Saf. 2019, 176, 95–107. [Google Scholar] [CrossRef]
- Kumar, A.; Pal, L.; Agrawal, V. Glutathione and citric acid modulates lead-and arsenic-induced phytotoxicity and genotoxicity responses in two cultivars of Solanum lycopersicum L. Acta Physiol. Plant. 2017, 39, 151. [Google Scholar] [CrossRef]
- Zheng, W.; Zhong, Z.Y.; Wang, H.B.; Wang, H.J.; Wu, D.M. Effects of oxalic acid on arsenic uptake and the physiological responses of Hydrilla verticillata exposed to different forms of arsenic. Bull. Environ. Contam. Toxicol. 2018, 100, 653–658. [Google Scholar] [CrossRef] [PubMed]
- Samanta, S.; Banerjee, A.; Roychoudhury, A. Melatonin application differentially modulates the enzymes associated with antioxidative machinery and ascorbate-glutathione cycle during arsenate exposure in indica rice varieties. Plant Biol. 2021, 23, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Das, J.; Sarkar, P. Remediation of arsenic in mung bean (Vigna radiata) with growth enhancement by unique arsenic-resistant bacterium Acinetobacter lwoffii. Sci. Total Environ. 2018, 624, 1106–1118. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ahammed, G.J.; Zhang, X.N.; Zhang, L.; Yan, P.; Zhang, L.P.; Fu, J.Y.; Han, W.Y. Melatonin-mediated regulation of anthocyanin biosynthesis and antioxidant defense confer tolerance to arsenic stress in Camellia sinensis L. J. Hazard. Mater. 2021, 403, 123922. [Google Scholar] [CrossRef] [PubMed]
- Dolui, D.; Hasanuzzaman, M.; Saha, I.; Ghosh, A.; Adak, M.K. Amelioration of sodium and arsenic toxicity in Salvinia natans L. with 2, 4-D priming through physiological responses. Environ. Sci. Pollut. Res. 2022, 29, 9232–9247. [Google Scholar] [CrossRef]
- Singh, S.; Husain, T.; Kushwaha, B.K.; Suhel, M.; Fatima, A.; Mishra, V.; Singh, S.K.; Tripathi, D.K.; Rai, M.; Prasad, S.M.; et al. Regulation of ascorbate-glutathione cycle by exogenous nitric oxide and hydrogen peroxide in soybean roots under arsenate stress. J. Hazard. Mater. 2020, 409, 123686. [Google Scholar] [CrossRef]
- Rahman, A.; Mostofa, M.G.; Alam, M.M.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Calcium mitigates arsenic toxicity in rice seedlings by reducing arsenic uptake and modulating the antioxidant defense and glyoxalase systems and stress markers. BioMed Res. Int. 2015, 2015, 340812. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Parihar, P.; Prasad, S.M. Sulphur and calcium attenuate arsenic toxicity in Brassica by adjusting ascorbate–glutathione cycle and sulphur metabolism. J. Plant Growth Regul. 2020, 91, 221–235. [Google Scholar] [CrossRef]
- Rohman, M.M.; Tonny, S.H.; Alam, S.S.; Omy, S.H.; Akhi, A.H.; Raihan, H.U.Z.; Akter, T.; Parvin, K.; Hasanuzzaman, M. Role of phytohormones in antioxidant metabolism in plants under salinity and water stress. In Managing Plant Production under Changing Environment; Hasanuzzaman, M., Ahammed, G.J., Nahar, K., Eds.; Springer: Singapore, 2022; pp. 151–191. [Google Scholar]
- Farooq, M.A.; Gill, R.A.; Islam, F.; Ali, B.; Liu, H.; Xu, J.; He, S.; Zhou, W. Methyl jasmonate regulates antioxidant defense and suppresses arsenic uptake in Brassica napus L. Front. Plant Sci. 2016, 7, 468. [Google Scholar] [CrossRef] [Green Version]
- Verma, G.; Srivastava, D.; Narayan, S.; Shirke, P.A.; Chakrabarty, D. Exogenous application of methyl jasmonate alleviates arsenic toxicity by modulating its uptake and translocation in rice (Oryza sativa L.). Ecotoxicol. Environ. Saf. 2020, 201, 110735. [Google Scholar] [CrossRef]
- Mousavi, S.R.; Niknejad, Y.; Fallah, H.; Tari, D.B. Methyl jasmonate alleviates arsenic toxicity in rice. Plant Cell Rep. 2020, 39, 1041–1060. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Cui, S.; Wu, D.; Yang, X.; Wang, H.; Wang, Z. Effects of kinetin on arsenic speciation and antioxidative enzymes in fronds of the arsenic hyperaccumulator Pteris cretica var. nervosa and non-hyperaccumulator Pteris ensiformis. Environ. Exp. Bot. 2021, 191, 104622. [Google Scholar] [CrossRef]
- Singh, M.; Singh, V.P.; Dubey, G.; Prasad, S.M. Exogenous proline application ameliorates toxic effects of arsenate in Solanum melongena L. seedlings. Ecotoxicol. Environ. Saf. 2015, 117, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Da-Silva, C.J.; Canatto, R.A.; Cardoso, A.A.; Ribeiro, C.; de Oliveira, J.A. Oxidative stress triggered by arsenic in a tropical macrophyte is alleviated by endogenous and exogenous nitric oxide. Braz. J. Bot. 2018, 41, 21–28. [Google Scholar] [CrossRef]
- Ghorbani, A.; Pishkar, L.; Roodbari, N.; Pehlivan, N.; Wu, C. Nitric oxide could allay arsenic phytotoxicity in tomato (Solanum lycopersicum L.) by modulating photosynthetic pigments, phytochelatin metabolism, molecular redox status and arsenic sequestration. Plant Physiol. Biochem. 2021, 167, 337–348. [Google Scholar] [CrossRef]
- Ghorbani, A.; Pishkar, L.; Roodbari, N.; Tavakoli, S.A.; Jahromi, E.M.; Wu, C. Nitrate reductase is needed for methyl jasmonate-mediated arsenic toxicity tolerance of rice by modulating the antioxidant defense system, glyoxalase system and arsenic sequestration mechanism. J. Plant Growth Regul. 2022, 1–13. [Google Scholar] [CrossRef]
- Bhat, J.A.; Bhat, M.A.; Abdalmegeed, D.; Yu, D.; Chen, J.; Bajguz, A.; Ahmad, A.; Ahmad, P. Newly-synthesized iron-oxide nanoparticles showed synergetic effect with citric acid for alleviating arsenic phytotoxicity in soybean. Environ. Pollut. 2021, 295, 118693. [Google Scholar] [CrossRef]
- Shahid, M.; Rafiq, M.; Niazi, N.K.; Dumat, C.; Shamshad, S.; Khalid, S.; Bibi, I. Arsenic accumulation and physiological attributes of spinach in the presence of amendments: An implication to reduce health risk. Environ. Sci. Pollut. Res. 2017, 24, 16097–16106. [Google Scholar] [CrossRef]
- Rafiq, M.; Shahid, M.; Abbas, G.; Shamshad, S.; Khalid, S.; Niazi, N.K.; Dumat, C. Comparative effect of calcium and EDTA on arsenic uptake and physiological attributes of Pisum sativum. Int. J. Phytoremediat. 2017, 19, 662–669. [Google Scholar] [CrossRef]
- Hakeem, K.R.; Alharby, H.F.; Bamagoos, A.A.; Pirzadah, T.B. Biochar promotes arsenic (As) immobilization in contaminated soils and alleviates the As-toxicity in soybean (Glycine max (L.) Merr.). Chemosphere 2022, 292, 133407. [Google Scholar] [CrossRef]
- Kamran, M.A.; Bibi, S.; Chen, B. Preventative effect of crop straw-derived biochar on plant growth in an arsenic polluted acidic ultisol. Sci. Total Environ. 2021, 812, 151469. [Google Scholar] [CrossRef] [PubMed]
- Xiao, A.W.; Li, Z.; Li, W.C.; Ye, Z.H. The effect of plant growth-promoting rhizobacteria (PGPR) on arsenic accumulation and the growth of rice plants (Oryza sativa L.). Chemosphere 2020, 242, 125136. [Google Scholar]
- Mondal, S.; Pramanik, K.; Ghosh, S.K.; Pal, P.; Mondal, T.; Soren, T.; Maiti, T.K. Unraveling the role of plant growth-promoting rhizobacteria in the alleviation of arsenic phytotoxicity: A review. Microbiol. Res. 2021, 250, 126809. [Google Scholar] [CrossRef]
- Pandey, N.; Bhatt, R. Role of soil associated Exiguobacterium in reducing arsenic toxicity and promoting plant growth in Vigna radiata. Eur. J. Soil Biol. 2016, 75, 142–150. [Google Scholar] [CrossRef]
- Pandey, S.; Ghosh, P.K.; Ghosh, S.; De, T.K.; Maiti, T.K. Role of heavy metal resistant Ochrobactrum sp. and Bacillus spp. strains in bioremediation of a rice cultivar and their PGPR like activities. J. Microbiol. 2013, 51, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Xiong, D.; Zhao, P.; Yu, X.; Tu, B.; Wang, G. Effect of applying an arsenic-resistant and plant growth–promoting rhizobacterium to enhance soil arsenic phytoremediation by Populus deltoides LH05-17. J. Appl. Microbiol. 2011, 111, 1065–1074. [Google Scholar] [CrossRef]
- Pinter, I.F.; Salomon, M.V.; Berli, F.; Bottini, R.; Piccoli, P. Characterization of the As (III) tolerance conferred by plant growth promoting rhizobacteria to in vitro-grown grapevine. Appl. Soil Ecol. 2017, 109, 60–68. [Google Scholar] [CrossRef]
- Ghosh, P.K.; Maiti, T.K.; Pramanik, K.; Ghosh, S.K.; Mitra, S.; De, T.K. The role of arsenic resistant Bacillus aryabhattai MCC3374 in promotion of rice seedlings growth and alleviation of arsenic phytotoxicity. Chemosphere 2018, 211, 407–419. [Google Scholar] [CrossRef]
- Soto, J.; Ortiz, J.; Herrera, H.; Fuentes, A.; Almonacid, L.; Charles, T.C.; Arriagada, C. Enhanced arsenic tolerance in Triticum aestivum inoculated with arsenic-resistant and plant growth promoter microorganisms from a heavy metal-polluted soil. Microorganisms 2019, 7, 348. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, A.; Pramanik, K.; Bhattacharya, S.; Mondal, S.; Ghosh, S.K.; Ghosh, P.K.; Maiti, T.K. Abatement of arsenic-induced phytotoxic effects in rice seedlings by an arsenic-resistant Pantoea dispersa strain. Environ. Sci. Pollut. Res. 2021, 28, 21633–21649. [Google Scholar] [CrossRef]
- Ghorbani, A.; Tafteh, M.; Roudbari, N.; Pishkar, L.; Zhang, W.; Wu, C. Piriformospora indica augments arsenic tolerance in rice (Oryza sativa) by immobilizing arsenic in roots and improving iron translocation to shoots. Ecotoxicol. Environ. Saf. 2021, 209, 111793. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, M.A.; Jiao, Y.; Chen, C.; Shireen, F.; Zheng, Z.; Imtiaz, M.; Bie, Z.; Huang, Y. Melatonin pretreatment improves vanadium stress tolerance of watermelon seedlings by reducing vanadium concentration in the leaves and regulating melatonin biosynthesis and antioxidant-related gene expression. J. Plant Physiol. 2018, 220, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Farouk, S.; Al-Amri, S.M. Exogenous melatonin-mediated modulation of arsenic tolerance with improved accretion of secondary metabolite production, activating antioxidant capacity and improved chloroplast ultrastructure in rosemary herb. Ecotoxicol. Environ. Saf. 2019, 180, 333–347. [Google Scholar] [CrossRef] [PubMed]
- Samanta, S.; Banerjee, A.; Roychoudhury, A. Arsenic toxicity is counteracted by exogenous application of melatonin to different extents in arsenic-susceptible and arsenic-tolerant rice cultivars. J. Plant Growth Regul. 2021. [Google Scholar] [CrossRef]
- Jung, H.I.; Kong, M.S.; Chae, M.J.; Lee, E.J.; Jung, G.B.; Kim, Y.H. Effect of ascorbate on the arsenic uptake, ROS-scavenging capacity, and antioxidant homeostasis in rice. Korean J. Soil Sci. Fertil. 2018, 51, 90–100. [Google Scholar] [CrossRef]
- Jung, H.; Kong, M.S.; Lee, B.R.; Kim, T.H.; Chae, M.J.; Lee, E.J.; Jung, G.B.; Lee, C.H.; Sung, J.K.; Kim, Y.H. Exogenous glutathione increases arsenic translocation into shoots and alleviates arsenic-induced oxidative stress by sustaining ascorbate–glutathione homeostasis in rice seedlings. Front. Plant Sci. 2019, 10, 1089. [Google Scholar] [CrossRef]
- Jiang, M.; Song, Y.; Kanwar, M.K.; Ahammed, G.J.; Shao, S.; Zhou, J. Phytonanotechnology applications in modern agriculture. J. Nanobiotechnol. 2021, 19, 430. [Google Scholar] [CrossRef]
- Singh, A.; Tiwari, S.; Pandey, J.; Lata, C.; Singh, I.K. Role of nanoparticles in crop improvement and abiotic stress management. J. Biotechnol. 2021, 337, 57–70. [Google Scholar] [CrossRef]
- Bidi, H.; Fallah, H.; Niknejad, Y.; Barari Tari, D. Iron oxide nanoparticles alleviate arsenic phytotoxicity in rice by improving iron uptake, oxidative stress tolerance and diminishing arsenic accumulation. Plant Physiol. Biochem. 2021, 163, 348–357. [Google Scholar] [CrossRef]
- Yan, S.; Wu, F.; Zhou, S.; Yang, J.; Tang, X.; Ye, W. Zinc oxide nanoparticles alleviate the arsenic toxicity and decrease the accumulation of arsenic in rice (Oryza sativa L.). BMC Plant Biol. 2021, 21, 150. [Google Scholar] [CrossRef]
- Kiany, T.; Pishkar, L.; Sartipnia, N.; Iranbakhsh, A.; Barzin, G. Effects of silicon and titanium dioxide nanoparticles on arsenic accumulation, phytochelatin metabolism, and antioxidant system by rice under arsenic toxicity. Environ. Sci. Pollut. Res. 2022. [Google Scholar] [CrossRef] [PubMed]
- Zeeshan, M.; Hu, Y.X.; Iqbal, A.; Salam, A.; Liu, Y.X.; Muhammad, I.; Ahmad, S.; Khan, A.H.; Hale, B.; Wu, H.Y.; et al. Amelioration of AsV toxicity by concurrent application of ZnO-NPs and Se-NPs is associated with differential regulation of photosynthetic indexes, antioxidant pool and osmolytes content in soybean seedling. Ecotoxicol. Environ. Saf. 2021, 225, 112738. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, D.K.; Singh, S.; Singh, V.P.; Prasad, S.M.; Chauhan, D.K.; Dubey, N.K. Silicon nanoparticles more efficiently alleviate arsenate toxicity than silicon in maize cultiver and hybrid differing in arsenate tolerance. Front. Environ. Sci. 2016, 4, 46. [Google Scholar] [CrossRef] [Green Version]
- González-Moscoso, M.; Juárez-Maldonado, A.; Cadenas-Pliego, G.; Meza-Figueroa, D.; SenGupta, B.; Martínez-Villegas, N. Silicon nanoparticles decrease arsenic translocation and mitigate phytotoxicity in tomato plants. Environ. Sci. Pollut. Res. Int. 2022, 1–17. [Google Scholar] [CrossRef]
- Shabnam, N.; Kim, M.; Kim, H. Iron (III) oxide nanoparticles alleviate arsenic induced stunting in Vigna radiata. Ecotoxicol. Environ. Saf. 2019, 183, 109496. [Google Scholar] [CrossRef]
- Yang, J.; Gao, M.X.; Hu, H.; Ding, X.M.; Lin, H.W.; Wang, L.; Xu, J.M.; Mao, C.Z.; Zhao, F.J.; Wu, Z.C. OsCLT1, a CRT-like transporter 1, is required for glutathione homeostasis and arsenic tolerance in rice. New Phytol. 2016, 211, 658–670. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Wang, X.; Liu, J.; Jian, P.; You, S.; Ding, N.; Guo, Q.; Lin, F. Applications of nanomaterials for heavy metal removal from water and soil: A review. Sustainability 2021, 13, 713. [Google Scholar] [CrossRef]
- Kandhol, N.; Aggarwal, B.; Bansal, R.; Parveen, N.; Singh, V.P.; Chauhan, D.K.; Sonah, H.; Sahi, S.; Grillo, R.; Peralta-Videa, J.; et al. Nanoparticles as a potential protective agent for arsenic toxicity alleviation in plants. Environ. Pollut. 2022, 300, 118887. [Google Scholar] [CrossRef]
- Wu, F.; Fang, Q.; Yan, S.; Pan, L.; Tang, X.; Ye, W. Effects of zinc oxide nanoparticles on arsenic stress in rice (Oryza sativa L.): Germination, early growth, and arsenic uptake. Environ. Sci. Pollut. Res. 2020, 27, 26974–26981. [Google Scholar] [CrossRef]
- Srivastava, S.; Suprasanna, P.; D’souza, S.F. Mechanisms of arsenic tolerance and detoxification in plants and their application in transgenic technology: A critical appraisal. Int. J. Phytoremediat. 2012, 14, 506–517. [Google Scholar] [CrossRef]
- Kumari, P.; Rastogi, A.; Shukla, A.; Srivastava, S.; Yadav, S. Prospects of genetic engineering utilizing potential genes for regulating arsenic accumulation in plants. Chemosphere 2018, 211, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, S.; Chauhan, R.; Srivastava, S.; Tripathi, R.D. The journey of arsenic from soil to grain in rice. Front. Plant Sci. 2017, 8, 1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Bermejo, E.; Castrillo, G.; Del Llano, B.; Navarro, C.; Zarco-Fernandez, S.; Martinez-Herrera, D.J.; Leo-del Puerto, Y.; Munoz, R.; Camara, C.; Paz-Ares, J.; et al. Natural variation in arsenate tolerance identifies an arsenate reductase in Arabidopsis thaliana. Nat. Commun. 2014, 5, 4617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, D.Y.; Chen, Y.; Chen, J.; Shi, S.; Chen, Z.; Wang, C. Genome-wide association mapping identifies a new arsenate reductase enzyme critical for limiting arsenic accumulation in plants. PLoS Biol. 2014, 12, e1002009. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Shi, S.; Wang, L.; Tang, Z.; Lv, T.; Zhu, X. OsHAC4 is critical for arsenate tolerance and regulates arsenic accumulation in rice. New Phytol. 2017, 215, 1090–1101. [Google Scholar] [CrossRef] [Green Version]
- Sundaram, S.; Wu, S.; Ma, L.Q.; Rathinasabapathi, B. Expression of a Pteris vittate glutaredoxin PvGRX5 in transgenic Arabidopsis thaliana increases plant arsenic tolerance and decreases arsenic accumulation in the leaves. Plant Cell Environ. 2009, 32, 851–858. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Chopra, P.; Chhillar, H.; Ahanger, M.A.; Hussain, S.J.; Maheshwari, C. Regulatory hubs and strategies for improving heavy metal tolerance in plants: Chemical messengers, omics and genetic engineering. Plant Physiol. Biochem. 2021, 164, 260–278. [Google Scholar] [CrossRef]
- Kumar, S.; Trivedi, P.K. Glutathione S-transferases: Role in combating abiotic stresses including arsenic detoxification in plants. Front. Plant Sci. 2018, 9, 751. [Google Scholar] [CrossRef] [Green Version]
- Yadav, S.K. Heavy metals toxicity in plants: An overview on the role of glutathione and phytochelatins in heavy metal stress tolerance of plants. S. Afr. J. Bot. 2010, 76, 167–179. [Google Scholar] [CrossRef] [Green Version]
- Ronzan, M.; Zanella, L.; Fattorini, L.; Della Rovere, F.; Urgast, D.; Cantamessa, S.; Nigro, A.; Barbieri, M.; Sanità di Toppi, L.; Berta, G.; et al. The morphogenic responses and phytochelatin complexes induced by arsenic in Pteris vittata change in the presence of cadmium. Environ. Exp. Bot. 2017, 133, 176–187. [Google Scholar] [CrossRef]
- Pathare, V.; Srivastava, S.; Suprasanna, P. Evaluation of effects of arsenic on carbon, nitrogen and sulphur metabolism in two contrasting varieties of Brassica juncea. Acta Physiol. Plant. 2013, 35, 3377–3389. [Google Scholar] [CrossRef]
- Chen, J.; Yang, L.; Yan, X.; Liu, Y.; Wang, R.; Fan, T.; Ren, Y.; Tang, X.; Xiao, F.; Liu, Y.; et al. Zinc-Finger transcription factor ZAT6 positively regulates cadmium tolerance through the glutathione-dependent pathway in Arabidopsis. Plant Physiol. 2016, 171, 707–719. [Google Scholar] [CrossRef] [PubMed]
- Khare, D.; Mitsuda, N.; Lee, S.; Song, W.Y.; Hwang, D.; Ohme-Takagi, M.; Hwang, J.U. Root avoidance of toxic metals requires the GeBP-LIKE 4 transcription factor in Arabidopsis thaliana. New Phytol. 2017, 213, 1257–1273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuce, M.; Taspinar, M.S.; Aydin, M.; Agar, G. Response of NAC transcription factor genes against chromium stress in sunflower (Helianthus annuus L.). Plant Cell Tissue Organ Cult. 2019, 136, 479–487. [Google Scholar] [CrossRef]
- Zhang, P.; Wang, R.; Ju, Q.; Li, W.; Tran, L.S.P.; Xu, J. The R2R3-MYB transcription factor MYB49 regulates cadmium accumulation. Plant Physiol. 2019, 180, 529–542. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, Y.; Liu, Y.; Xu, H.; Zhang, T.; Hu, Z.; Lou, L.; Cai, Q. Ectopic expression of wheat aquaglyceroporin TaNIP2; 1 alters arsenic accumulation and tolerance in Arabidopsis thaliana. Ecotoxicol. Environ. Saf. 2020, 205, 111131. [Google Scholar] [CrossRef]
- Manuka, R.; Saddhe, A.A.; Srivastava, A.K.; Kumar, K.; Penna, S. Overexpression of rice OsWNK9 promotes arsenite tolerance in transgenic Arabidopsis plants. J. Biotechnol. 2021, 332, 114–125. [Google Scholar] [CrossRef]
- Kidwai, M.; Dhar, Y.V.; Gautam, N.; Tiwari, M.; Ahmad, I.Z.; Asif, M.H.; Chakrabarty, D. Oryza sativa class III peroxidase (OsPRX38) overexpression in Arabidopsis thaliana reduces arsenic accumulation due to apoplastic lignification. J. Hazard. Mater. 2019, 362, 383–393. [Google Scholar] [CrossRef]
- Verma, P.K.; Verma, S.; Pande, V.; Mallick, S.; Deo Tripathi, R.; Dhankher, O.P.; Chakrabarty, D. Overexpression of rice glutaredoxin OsGrx_C7 and OsGrx_C2.1 reduces intracellular arsenic accumulation and increases tolerance in Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 740. [Google Scholar] [CrossRef] [Green Version]
- Zanella, L.; Fattorini, L.; Brunetti, P.; Roccotiello, E.; Cornara, L.; D’Angeli, S.; Della Rovere, F.; Cardarelli, M.; Barbieri, M.; Sanita di Toppi, L.; et al. Overexpression of AtPCS1 in tobacco increases arsenic and arsenic plus cadmium accumulation and detoxification. Planta 2016, 243, 605–622. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, H.Y.; Chen, Y.F. The transcription factor MYB40 is a central regulator in arsenic resistance in Arabidopsis. Plant Commun. 2021, 2, 100234. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, M.; Kidwai, M.; Dutta, P.; Narayan, S.; Gautam, N.; Chawda, K.; Shirke, P.A.; Mishra, A.K.; Chakrabarty, D. A tau class glutathione-S-transferase (OsGSTU5) confers tolerance against arsenic toxicity in rice by accumulating more arsenic in root. J. Hazard. Mater. 2022, 426, 128100. [Google Scholar] [CrossRef] [PubMed]
- Nishida, S.; Duan, G.; Ohkama-Ohtsu, N.; Uraguchi, S.; Fujiwara, T. Enhanced arsenic sensitivity with excess phytochelatin accumulation in shoots of a SULTR1; 2 knockout mutant of Arabidopsis thaliana (L.) Heynh. Soil Sci. Plant Nutr. 2016, 62, 367–372. [Google Scholar] [CrossRef] [Green Version]
- Luan, M.; Liu, J.; Liu, Y.; Han, X.; Sun, G.; Lan, W.; Luan, S. Vacuolar phosphate transporter 1 (VPT1) affects arsenate tolerance by regulating phosphate homeostasis in Arabidopsis. Plant Cell Physiol. 2018, 59, 1345–1352. [Google Scholar] [CrossRef]
- Kim, D.; Bahmani, R.; Modareszadeh, M.; Hwang, S. Mechanism for higher tolerance to and lower accumulation of arsenite in NtCyc07-overexpressing tobacco. Plants 2020, 9, 1480. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Species | Arsenic Levels and Growth Condition | Stress Period | Major Effects | Reference |
|---|---|---|---|---|
| Oryza sativa Japonica type | As2O3; 10, 20, 30, 40 and 50 μM; mixture of soil, perlite and vermicompost | 30 days from sowing | H2O2 and TBARS increased under As stress in dose dependent manner and reach up to 1.93 folds and 1.71-folds in 50 μM As, as compared to control. | [97] |
| Zea mays cv. DKC5741 | Na2HAsO4⋅7H2O; 100 μM; hydroponic | 14 days | 11% H2O2 and 61% TBARS content increased with As-stress over control. | [98] |
| Pisum sativum | NaAsO2; 20 μM; mixture of sand, perlite and pit; Hoagland solution | 40 days | 74 and 63% higher content of H2O2 and MDA was obtained as compared to control. Increased electrolyte leakage (69%) was observed over control (11%). | [99] |
| Lolium perenne cv. Mathilde, L. multiflorum cv. Idyll | NaAsO2; 2.51 mg mL−1; soil | 60 days | Increased H2O2 content by 44% and 32% in expanded leaves of perennial cultivar (Mathilde) and annual cultivar (Idyll). In mature, expanded and emerging leaves of perennial ryegrass, superoxide anion (O2−) increased by 26, 29 and 47%, respectively. While in annual ryegrass, mature, expanded and emerging leaves of annual ryegrass 30, 5 and 65% respectively, higher O2•−. | [100] |
| Solanum lycopersicum cv. SC 2121 | Na2HAsO4⋅7H2O; 50 μM; hydroponic | 10 days | Leaf H2O2 and MDA content increased by 242 and 272% over control. Activity of LOX in leaf enhanced by 127%. EL increased by 325% over control. | [101] |
| Spinacia oleracea | 25 μM and 125 μM As, NaAsO2; hydroponics | Four weeks | Increase H2O2 in roots by 32% and 65% in dose dependent manner while in leaves it was reduced over control. TBARS content increased by 90 and 92%, respectively, in 25 μM and 125 μM As over control. | [102] |
| Glycine max cv. JS 335 | 10 and 100 μM NaAsO2; pre-soaked filter paper | 2 and 5 days, respectively | ROS levels significantly enhanced in concentration dependent manner (OH•: 198 to 524%, H2O2: 234 to 539% from 10 to 100 μM, respectively). LOX activity significantly enhanced with extended exposure (503 and 1193%, on 2 and 5 day). | [83] |
| Arabidopsis thaliana | 100, 200 and 300 μM NaH2AsO4; ½ MS medium | 7 days | TBARS content increased by 16 and 38% in 200 and 300 μM As, respectively over control. | [103] |
| Brassica napus cv. Zheda 622 and ZS 758 | 200 μM NaAsO2; peat and soil mixture | 14 days | MDA content in roots enhanced by 101% in the cv. ZS 758 and by 178% in the cv. Zheda 622 over control. Superoxide radicals (O2•ˉ) and H2O2, approximately doubled over control in roots. | [104] |
| Nicotiana tabacum cv. Wisconsin and N. sylvestris | 10 and 30 µg L−1 Na2HAsO4.7H2O; perlite and sand (1:1); Hoagland solution | 7 weeks | Reactive oxygen species was increased in root and leaves only with 30 µg L−1 over control while two-fold increase observed in roots and leaves of sensitive N. sylvestris. MDA content in sensitive N. sylvestris was increased with increasing As level, while there are no significant differences recorded in N. tabacum cv. Wisconsin. | [41] |
| Plant Species | Plant Organ | Proteomic Technology | Induced Enzymes | Reference |
|---|---|---|---|---|
| Oryza sativa | Root | In-gel, MALDI-TOF MS | GST | [109] |
| Spinacia oleracea | Leaf | 2D-MS/MS | FeSOD, GST | [110] |
| O. sativa | Leaf | 2-DE/MALDI-TOF-TOF | APX, GST, cytochome c peroxidase | [111] |
| O. sativa | Leaf | 2-DE/MALDI-TOF-TOF | Cu/ZnSOD | [112] |
| Brassica napus | Leaf | RPLC-MS-MS/iTRAQ | GR, Cu/ZnSOD, CAT, FeSOD | [104] |
| B. napus | Leaf | RPLC-MS-MS/iTRAQ | CAT, POD, SOD, GST | [113] |
| Populus deltoides and Populus × euramericana | Root | 2-DE/MALDI-TOF-TOF-MS | Cu/ZnSOD, POD | [114] |
| Artemisia annua | Leaf | 2-DE/MALDI-TOF-TOF-MS | APX, DHAR | [115] |
| Zea mays | Root | 2-DE/MALDI-TOF-MS | GPX, Cu/ZnSOD | [116] |
| Plant Species | Plant Organs | Oxidative Stress Indicators * | Arsenic Levels | Contrasting Antioxidant Defense in As Tolerant Plants * | Reference | |
|---|---|---|---|---|---|---|
| Enzymes | Contents of Non-Enzymatic Antioxidants | |||||
| Zea mays | Leaf | Electrolytic leakage > MDA > H2O2 > | NaAsO2 (200 μM As), CdCl2 (100 μM Cd); 45, 60, 75, and 90 d | SOD↑ POD↑ CAT↑ APX↑ GPX↑ GR↑ | GSH↑ AsA↑ | [131] |
| Oryza sativa | Root and shoot | MDA↑ | AsIII (NaAsO2; 10 and 25 μM) and AsV (Na2HAsO4; 0, 10, and 50 μM); 1, 4, and 7 d | APX↑ GPOX↑ SOD (root)↑ | Overall stress responsive amino acid accumulation > | [132] |
| O. sativa | Leaf | MDA < | 50, 150, and 300 μM As(V); 24 and 96 h | APX > CAT > SOD > | Cysteine > Pro > | [133] |
| Nicotiana sylvestris and N. tabacum | Leaf and root | MDA = ROS < | Na2HAsO4·7H2O (10 and 30 µg L−1); 7 weeks | APX (leaf)↓ GST (leaf)↓ POD(leaf)↓ APX (root)↑ GST (root) = CAT (root)↓ CAT (leaf)↑ | AsA (root)↑ Anthocyanin (leaf) = GSH/GSSG↓ Carotenoids, Phenolic compounds (leaf) = Phenolic compounds (root)↑ Pro (leaf)↑ Total glutathione (root) > Carbohydrate status = | [41] |
| O. sativa | Root and shoot | MDALI H2O2LI | Na2HAsO4·7H2O; 18 d | SOD↑ CATLD | Pro↑ | [134] |
| O. sativa | Root and shoot | MDALI H2O2LI | 25, 50 and 75 μM Na2HAsO4·7H2O; 18 d | SODLI CATLD | Pro < | [135] |
| Z. mays | Leaf | MDA < H2O2 < | 2.5, 7.5, and 12.5 µg mL−1 NaHAsO4·7H2O in 30% HNS; 3 and 7 d | SOD > GR > APX < GPOX < CAT < | Carotenoids↑ GSH = | [136] |
| O. sativa | Leaf | TBARS < H2O2 < | 4 μg mL−1 As(III) (NaHAsO4·7H2O) and As(V) (NaAsO2); 14 d | SOD < APX > GR > GST > | GSH < | [137] |
| O. sativa | Root and shoot | H2O2LI | 25 μM, 50 μM, and 100 μM Na2HAsO4.7H2O; 18 d | CATLD APXLI GRLI GST↑ | AsALI GSHLI ProLI | [130] |
| Pteris vittata and P. ensiformis | Frond | TBARS < H2O2 < | 133 and 267 μM of As as Na2HAsO4.7 H2O; 1, 5 and 10 d | NC | AsA > Carotenoid > GSH > | [13] |
| Ricinus communis | Leaf | MDA = H2O2 = | 100 and 200 μM Na2HAsO4·7H2O; 21 d | Mn-SOD = | Carotenoid (no contrasting differences) | [45] |
| R. communis | Leaf and root | NC | 100 and 200 μM As(V) | CAT = SOD↑ GPOX↑ | Pro = | [107] |
| Brassica juncea | Shoot | NC | As(V) (50 μM and 500 μM) and As(III) (25 μM and 250 μM); 7 d and 15 d | NC | Cysteine↑ GSH↑ Non-protein thiols↑ | [138] |
| B. juncea | Shoot | MDA = | As(V) (50 and 500 μM) and As(III) (25 and 250 μM); 7 or 15 d | DHAR↑ MDHAR↑ SOD > APX↑ GPOX↑ | GSH↑ GSH/GSSG↑ Pro↑ | [139] |
| O. sativa | Root and shoot | NO < NADPH oxidase < Ascorbate oxidase < | As(V) (NaAsO2; 5, 10, 25 μM); 15 d | GPX > | AsA > CarotenoidsLDPro > | [140] |
| P. cretica and Spinacia oleracea | Shoot | NC | 20 and 100 mg As kg−1 soil; NC | NC | CarotenoidsLD | [42] |
| Plant Species | Arsenic Levels | Exogenous Elicitors | Defense Responses | References |
|---|---|---|---|---|
| Oryza sativa L. cv. minakshi | 60 μM Na2AsO3; 7d | 10 μM Se (Na2SeO4); co-treatment | Reduced the generation of H2O2 and MDA by 23 and 35%, respectively. Decreased AsA content by about 15%. Reduced the activity of CAT, SOD and APX by 8, 23 and 9%, respectively. | [141] |
| Vicia faba L. cv. Tara | 5 μM Na2AsO3, 27 d | 30 mM CaCl2, as co-treatment | Suppressed ROS generation as O2•− and H2O2 contents and their histochemical detection. Reduced NADPH oxidase and glycolate oxidase (GOX) activities. Increased SOD, APX, MDHAR and DHAR activities. Strengthen membrane stability. | [73] |
| O. sativa L. var. Narendra | 50 μM NaAsO2, 15 d | Si, silicon (10 μM); co-treatment | Increased AsA content and AsA/DHA with higher activity of MDHAR, DHAR | [142] |
| Arabidopsis thaliana L. | 100 and 200 μM Na2HAsO4⋅7H2O, 24 h | 1 μM 24-Epibrassinolide (EBL) | Increased total antioxidant capacity with higher SOD and CAT activities. Reduced MDA content. | [143] |
| O. sativa L. cv. Swarna Sub1 | 50 μM NaAsO2, 240 h | 10 μM of ABA, 24 h as pretreatment | Reduced O2•− and H2O2 by 50 and 38%, respectively. Reduced lipid peroxidation (TBARS) by 48%. Reduced membrane damage as decreased EL. Significantly increased GSH/GSSG with higher GR activity. Upregulated antioxidants activity like SOD, CAT, APX and GPX. | [125] |
| Zea mays L. cv. “DK5783” | 0.1 mM Na2HAs4O·7H2O, 28 d | 0.5 mM salicylic acid (SA as 2-hydroxybenzoic acid), pretreatment, 7 d. | Reduced H2O2, MDA production and EL. Decreased SOD activity with higher activity of CAT and POD. Improved AsA/DHA and GSH/GSSG by improving AsA and GSH content with reduction of DHA and GSSG level. Elevated the activity of enzymatic components of AsA-GSH pathway like APX, GR, MDHAR and DHAR. | [144] |
| O. sativa cv. Sarjoo52 | 25 μM NaAsO2, 7 d | Sodium nitroprusside (SNP) (30 μM; NO donor | Decreased the production of H2O2 and TBARS. Reduced O2•− generation detected by histochemical staining. Lowered SOD, CAT, GPX, APX activities. Improved GR activity. | [145] |
| Lemna valdiviana | 4.0 mg L−1 Na2HAsO4·7H2O, 24 h | 100 μM JA, co-treatment | Reduced O2•−, H2O2 and TBARS level. Elevated SOD, CAT, APX, GR activities. | [74] |
| Brassica napus L. cvs. Zheda 622 and ZS 758 | 200 μM NaAsO2, 14 d | 1.0 μM MeJA | Decreased MDA content with the higher activity of SOD, POD, CAT, APX. Elevated the GSH and GSSG level with the increasing GR activity. Increased AsA content. | [146] |
| Pisum sativum L. | 20 μM NaAsO2, 31 d | 200 μM H2S (NaHS), co-treatment | Suppressed H2O2, MDA and EL by 63, 57, and 44%, respectively. Increased the activity of SOD, CAT, APX, MDHAR, DHAR, GR and GST by 32, 71, 15, 43, 71, 19, and 27%, respectively. Augmented the content of AsA, GSH and GSSG by 67, 25 and 61%, respectively. Reduced the generation of MG by 35% with 18 and 44% higher activity of Gly I, and Gly II, respectively. | [99] |
| O. sativa L. var. Pusa Basmati | 150 μM NaAsO2; 48 h | 100 μM NO SNP; 24 h as pretreatment | Lowered cysteine content in both shoot and root (by 7 and 18%). Reduced H2O2 and MDA content by 7 and 19%, respectively in shoot and by 7 and 16% in roots. Decreased the activity of CAT, SOD, APX and GR in both shoot and root. | [147] |
| B. juncea var. Pusa Jagannath | 150 μM NaAsO2, 48 h NaAsO2 | 100 μM SNP, pre-incubation started before 24 h of stress | Reduced H2O2 and MDA in leaves (13 and 28%, respectively) and roots (18 and 20%, respectively). Decreased cysteine and Pro content significantly. Lowered down the level of GSH with reduction in the activities of SOD, CAT, APX and GR. Reduced the thiol components including both total thiol and non-protein thiol. | [148] |
| Solanum lycopersicum L. cv. Pusa ruby | 10 µM Na2HAsO4, 7 d | 250 µM citric acid (CA) | Reduced MDA and H2O2 content. Elevated the non-enzymatic antioxidants content with higher (62%) CAT activity. Decease the activity of APX and GR (89%). | [149] |
| S. lycopersicum L. cv. Pusa ruby | 10 µM Na2HAsO4, 7 d | 250 µM GSH | Reduced oxidative stress by lowering MDA and H2O2 contents. Elevated the CAT activity by 96% with lower down of APX and GR (123%) activities. | [149] |
| Hydrilla verticillata | 3 mg L−1 as As2O3 and Na2HAsO4·7H2O; 10 d | 200–2000 µg L−1 oxalic acid (OA) | Decreased TBRAS content with higher activity of SOD, POD and CAT | [150] |
| O. sativa L. | 150 μM Na3AsO, 2 d | 20 μM melatonin, co-treatment | Reduced protease activity. Lowered MDA content by 30% and LOX activity by 21%. Reduced MG content (14%) with higher activity of Gly I and Gly II by 20 and 12%, respectively. Decreased NADP oxidase (NOX) activity by 31%. The activity of ascorbic acid oxidase (AAO) and nitrate reductase (NR) were reduced and increased, respectively. Increased anthocyanin, flavonoid, xanthophylls, and total phenolic content by 92, 40, 90, and 20%. Increased total antioxidant capacity with 48% higher AsA content. Increased phenylalanine ammonia lyase (PAL) activity by 29%. | [151] |
| V. faba L. cv. Tara | 5 μM Na2AsO, 27 d | 50 μM melatonin as co-treatment | Reduced ROS including O2•− and H2O2 contents and the NADPH oxidase and GOX activity. Decreased membrane damage as indicated by lower MDA content and EL. Strengthened antioxidants defense mechanism by increasing SOD, AsA-GSH pathways. | [73] |
| Vigna radiata | 23 mg kg−1 Na2HAsO4·7H2O, 7 d | Acinetobacter lwoffi RJB-2 | Reduced MDA content and EL. | [152] |
| Camellia sinensis L. | 25 μM Na2HAsO4⋅7H2O, 30 d | 100 μM melatonin for 24 h as pretreatment | Reduced O2•− and H2O2 with lower MDA level. Elevated APX, SOD, CAT and POD activities with higher total antioxidants capacity. | [153] |
| Salvinia natans L. | 500 μM NaAsO2, 7 d | 500 μM of 2,4-dichlorophenoxyacetic acid (2,4-D), 3 d as pretreatment | Reduced NOX activity by 39%. Decreased TBARS by 47%. Reduced O2•− and H2O2 production by 54 and 48%, respectively led to lower 37% of EL. Elevated SOD and APX activity with reduction of CAT an GPX. Increased AsA content and GSH/GSSG with the elevated response of enzymatic components like MDHAR, DHAR and GR activity. | [154] |
| Glycine max var. JS 20–29 | 500 μM Na2HAsO4·7H2O, 7 d | 1 μM H2O2, 24 h as pretreatment | Reduced O2•−, H2O2 and MDA content. Elevated the activity of APX, MDHAR, DHAR and GR. Increased AsA, DHA, GSH, and GSSG contents resulted higher AsA/DHA and GSH/GSSG status. | [155] |
| Gene Transformed | Product | Donor Species | Recipient Species | Redox and Metabolite Changes | Reference |
|---|---|---|---|---|---|
| TaNIP2;1 | Silicon channel type transporter | Triticum aestivum | Arabidopsis thaliana | Increase in POD and CAT activities and lipid peroxidation | [216] |
| WNK1 | Serine/threonine protein kinase | O. sativa | A. thaliana | Increase in SOD, POD, and CAT activities | [217] |
| PRX38 | Class III peroxidase | O. sativa | A. thaliana | Increase in SOD, PRX and GST activity and low H2O2 content | [218] |
| GrxC2.1; GrxC7 | Glutaredoxin | O. sativa | A. thaliana | Rise in GSH content and depletion in GSSG content | [219] |
| PCS1 | Phytochelatin synthase | Arabidopsis thaliana | Nicotiana tabacum | Increase in GSH content | [220] |
| MYB40 | Transcription factor in As resistance | A. thaliana | A. thaliana | Increase in thiol-peptide accumulation | [221] |
| GSTU5 | Glutathione S-transferase | O. sativa | O. sativa | Increase in SOD and PRX activity | [222] |
| SULTR1;2 | Sulfate transporters | A. thaliana | A. thaliana | Reduction in GSH content in shoots and roots | [223] |
| CLT1 | CRT-like transporter | O. sativa | O. sativa | Decrease in GSH, GSSG and γ-glutamylcysteine levels | [92] |
| VPT1 | Vacuolar Phosphate Transporter 1 | A. thaliana | A. thaliana | Increase in anthocyanin level | [224] |
| Cyc07 | 40S ribosomal protein S3Ae | Nicotiana tabacum | N. tabacum | Augmentation in SOD, CAT and GR activities | [225] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nahar, K.; Rhaman, M.S.; Parvin, K.; Bardhan, K.; Marques, D.N.; García-Caparrós, P.; Hasanuzzaman, M. Arsenic-Induced Oxidative Stress and Antioxidant Defense in Plants. Stresses 2022, 2, 179-209. https://doi.org/10.3390/stresses2020013
Nahar K, Rhaman MS, Parvin K, Bardhan K, Marques DN, García-Caparrós P, Hasanuzzaman M. Arsenic-Induced Oxidative Stress and Antioxidant Defense in Plants. Stresses. 2022; 2(2):179-209. https://doi.org/10.3390/stresses2020013
Chicago/Turabian StyleNahar, Kamrun, Mohammad Saidur Rhaman, Khursheda Parvin, Kirti Bardhan, Deyvid Novaes Marques, Pedro García-Caparrós, and Mirza Hasanuzzaman. 2022. "Arsenic-Induced Oxidative Stress and Antioxidant Defense in Plants" Stresses 2, no. 2: 179-209. https://doi.org/10.3390/stresses2020013
APA StyleNahar, K., Rhaman, M. S., Parvin, K., Bardhan, K., Marques, D. N., García-Caparrós, P., & Hasanuzzaman, M. (2022). Arsenic-Induced Oxidative Stress and Antioxidant Defense in Plants. Stresses, 2(2), 179-209. https://doi.org/10.3390/stresses2020013