
City Living: Nest-Site Selection Preferences in Urban Herring Gulls, Larus argentatus



Abstract
:1. Introduction
1.1. Presence of Conspecifics
1.2. Availabiltiy of Food
1.3. Risk of Predation
1.4. Nest-Site Substrate
1.5. Research Questions
2. Materials and Methods
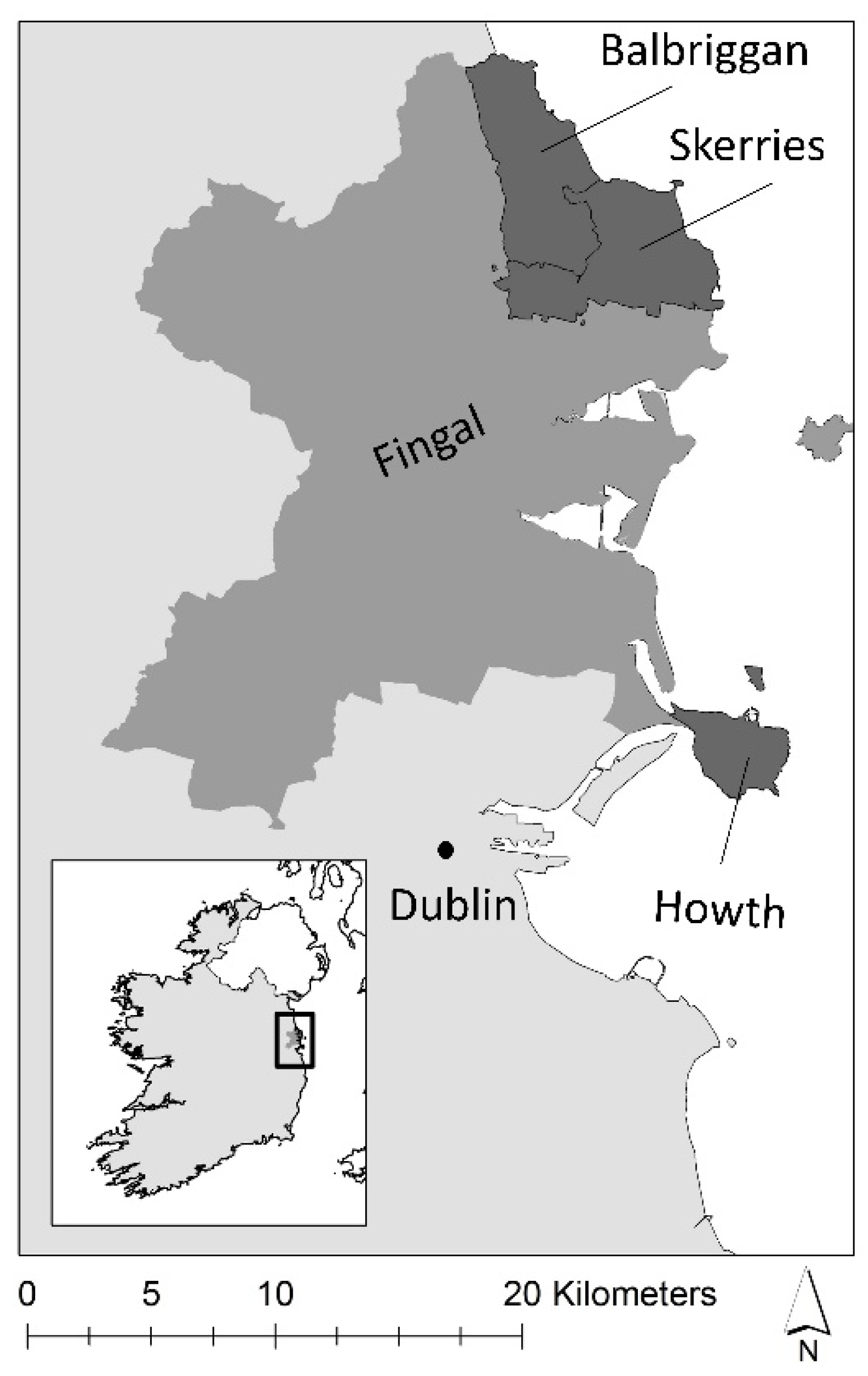
2.1. Study Area
2.2. Data Collection
2.3. Data Analysis
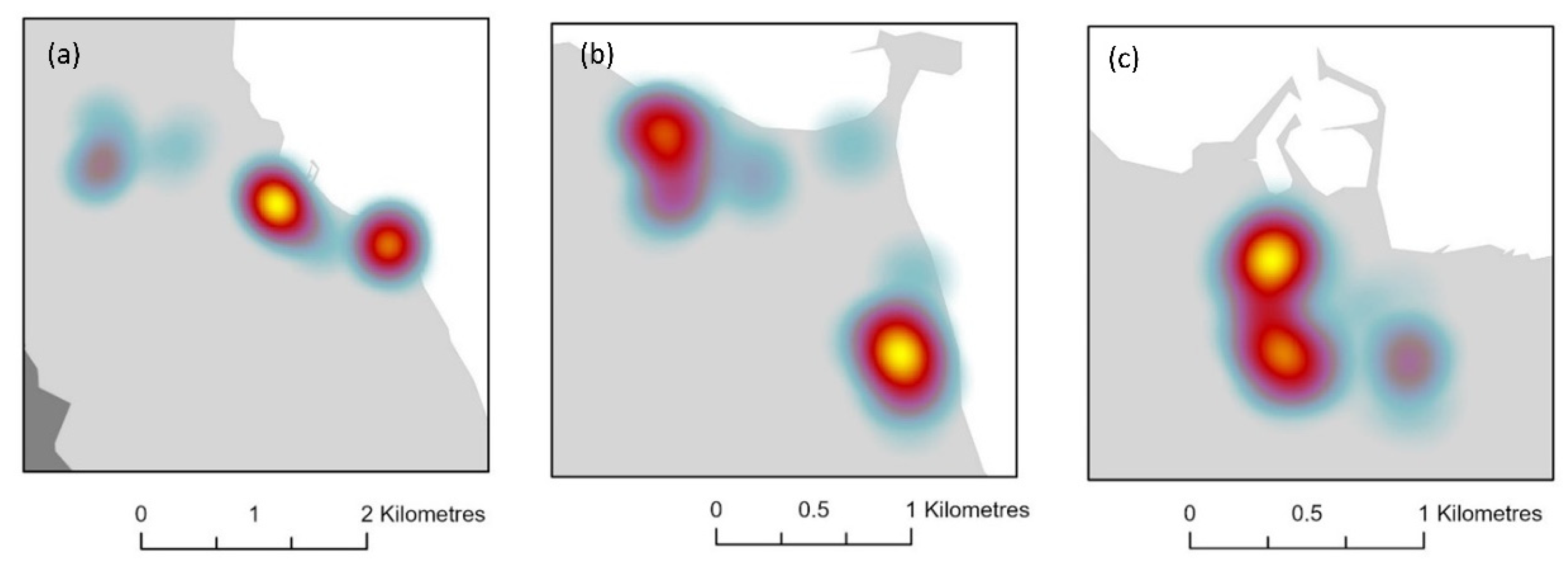
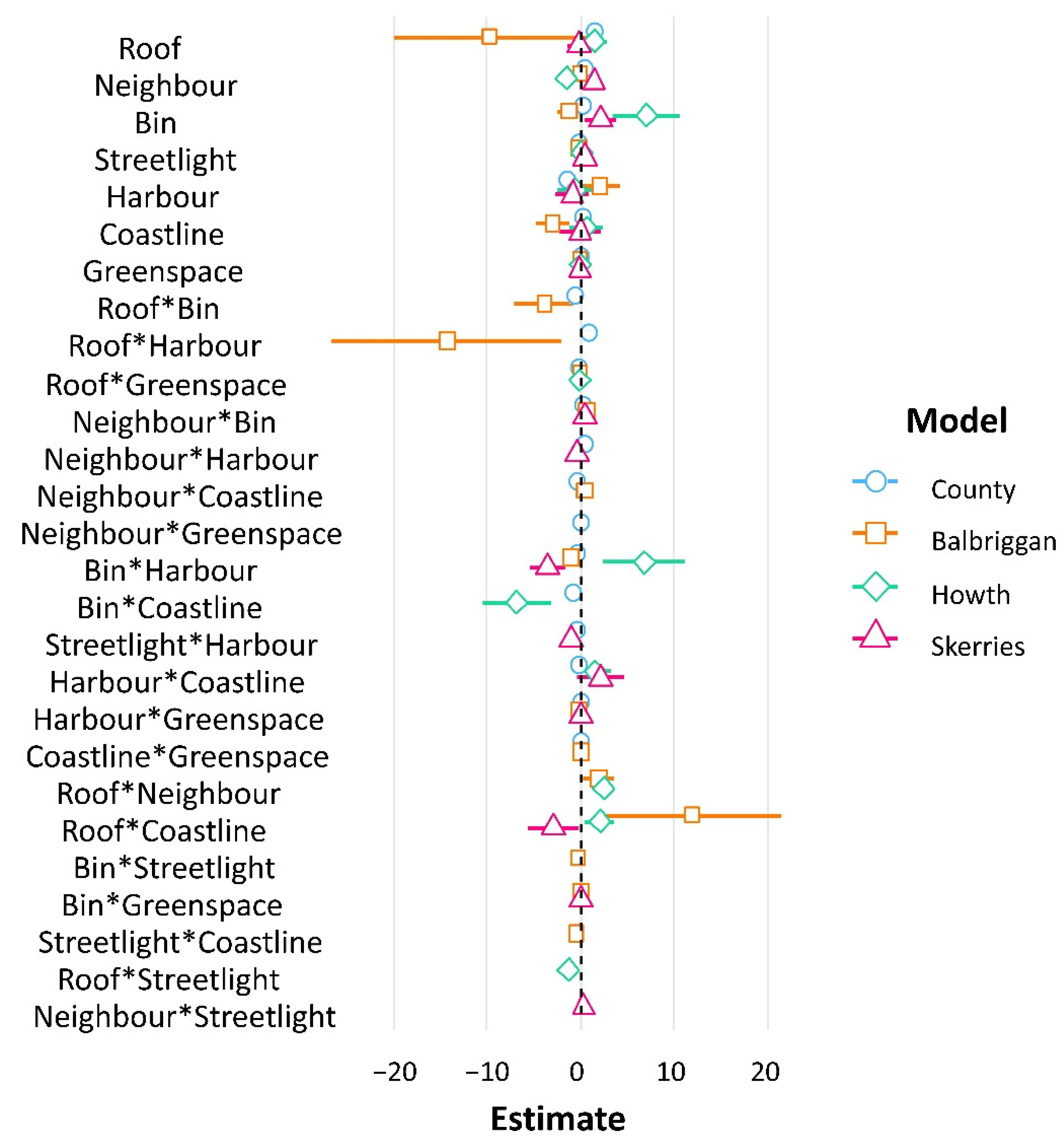
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Neumann, B.; Vafeidis, A.T.; Zimmermann, J.; Nicholls, R.J. Future Coastal Population Growth and Exposure to Sea-Level Rise and Coastal Flooding—A Global Assessment. PLoS ONE 2015, 10, e0131375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennie, J.; Duffy, J.P.; Davies, T.W.; Correa-Cano, M.E.; Gaston, K.J. Global Trends in Exposure to Light Pollution in Natural Terrestrial Ecosystems. Remote Sens. 2015, 7, 2715–2730. [Google Scholar] [CrossRef] [Green Version]
- Hastings, R.; Cummins, V.; Holloway, P. Assessing the impact of physical and anthropogenic environmental factors in determining the habitat suitability of seagrass ecosystems. Sustainability 2020, 12, 8302. [Google Scholar] [CrossRef]
- Holloway, P.; Field, R. Can rock-rubble groynes support similar intertidal ecological communities to natural rocky shores? Land 2020, 9, 131. [Google Scholar] [CrossRef]
- Belant, J.L. Gulls in urban environments: Landscape-level management to reduce conflict. Landsc. Urban Plan. 1997, 38, 245–258. [Google Scholar] [CrossRef]
- Lepczyk, C.A.; Warren, P.S. Urban Bird Ecology and Conservation; Studies in Avian Biology; University of California Press: Berkeley, CA, USA, 2012; Volume 45. [Google Scholar]
- Yoda, K.; Tomita, N.; Mizutani, Y.; Narita, A.; Niizuma, Y. Spatio-temporal responses of black-tailed gulls to natural and anthropogenic food resources. Mar. Ecol. Prog. Ser. 2012, 466, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Jones, J. Habitat Selection Studies in Avian Ecology: A Critical Review. Ornithology 2001, 118, 557–562. [Google Scholar] [CrossRef]
- Clucas, B.; Marzluff, J.M. Attitudes and Actions toward Birds in Urban Areas: Human Cultural Differences Influence Bird Behavior. Auk 2012, 129, 8–16. [Google Scholar]
- McCaffrey, R.E.; Mannan, R.W. How scale influences birds’ responses to habitat features in urban residential areas. Landsc. Urban Plan. 2012, 105, 274–280. [Google Scholar] [CrossRef]
- BirdLife International. European Red List of Birds; Office for Official Publications of the European Communities: Luxembourg, 2015. [Google Scholar]
- Madden, B.; Newton, S. Herring gull Larus argentatus. In Seabird Population of Britain and Ireland; A & C Black: London, UK, 2004; pp. 242–261. [Google Scholar]
- Monaghan, P.; Coulson, J.C.C. Status of Large Gulls Nesting on Buildings. Bird Study 1977, 24, 89–104. [Google Scholar] [CrossRef]
- Raven, S.J.; Coulson, J.C. The distribution and abundance of Larus gulls nesting on buildings in Britain and Ireland. Bird Study 1997, 44, 13–34. [Google Scholar] [CrossRef]
- Soldatini, C.; Albores-Barajas, Y.V.; Mainardi, D.; Monaghan, P. Roof nesting by gulls for better or worse? Ital. J. Zool. 2008, 75, 295–303. [Google Scholar] [CrossRef]
- Mainwaring, M.C. The use of man-made structures as nesting sites by birds: A review of the costs and benefits. J. Nat. Conserv. 2015, 25, 17–22. [Google Scholar] [CrossRef]
- Beasley, E.R. Foraging Habits, Population Changes, and Gull-Human Interactions in an Urban Population of Herring Gulls (Larus argentatus) and Lesser Black-backed (Larus fuscus). Master’s Thesis, Middlesex University, London, UK, 2017. [Google Scholar]
- Burger, J.; Shisler, J. Nest Site Selection and Competitive Interactions of Herring and Laughing Gulls in New Jersey. Auk 1978, 95, 252–266. [Google Scholar]
- Mainwaring, M.C.; Hartley, I.R.; Lambrechts, M.M.; Deeming, D.C. The design and function of birds’ nests. Ecol. Evol. 2014, 4, 3909–3928. [Google Scholar] [CrossRef]
- Isaac, B.; Cooke, R.; Simmons, D.; Hogan, F. Predictive mapping of powerful owl (Ninox strenua) breeding sites using Geographical Information Systems (GIS) in urban Melbourne, Australia. Landsc. Urban Plan. 2008, 84, 212–218. [Google Scholar] [CrossRef]
- Reynolds, S.J.; Ibáñez-Álamo, J.D.; Sumasgutner, P.; Mainwaring, M.C. Urbanisation and nest building in birds: A review of threats and opportunities. J. Ornithol. 2019, 160, 841–860. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.A.; Holloway, P. Incorporating movement in species distribution models. Prog. Phys. Geogr. 2015, 39, 837–849. [Google Scholar] [CrossRef]
- Heppner, J.J.; Ouyang, J.Q. Incubation behavior differences in urban and rural house wrens, Troglodytes aedon. Front. Ecol. Evol. 2021, 9, 89. [Google Scholar] [CrossRef]
- Thériault, S.; Villard, M.A.; Haché, S. Habitat selection in site-faithful ovenbirds and recruits in the absence of experimental attraction. Behav. Ecol. 2012, 23, 1289–1295. [Google Scholar] [CrossRef] [Green Version]
- Mariette, M.M.; Griffith, S.C. Conspecific attraction and nest site selection in a nomadic species, the Zebra finch. Oikos 2012, 121, 823–834. [Google Scholar] [CrossRef]
- Vermeer, K.; Power, D.; Smith, G.E.J. Habitat Selection and Nesting Biology of Roof-Nesting Glaucous-Winged Gulls. Colon. Waterbirds 1988, 11, 189–201. [Google Scholar] [CrossRef]
- Cunningham, J.A.; Kesler, D.C.; Lanctot, R.B. Habitat and social factors influence nest-site selection in Arctic-breeding shorebirds. Auk 2016, 133, 364–377. [Google Scholar] [CrossRef]
- O’Hanlon, N.J. Spatial Variation in Herring Gull Traits and Their Potential as Monitors of the Coastal Environment. Ph.D. Thesis, University of Glasgow, Glasgow, UK, 2016. [Google Scholar]
- Coogan, S.C.P.; Raubenheimer, D.; Zantis, S.P.; Machovsky-Capuska, G.E. Multidimensional nutritional ecology and urban birds. Ecosphere 2018, 9, e02177. [Google Scholar] [CrossRef] [Green Version]
- Soh, M.C.K.; Sodhi, N.S.; Seoh, R.K.H.; Brook, B.W. Nest site selection of the house crow (Corvus splendens), an urban invasive bird species in Singapore and implications for its management. Landsc. Urban Plan. 2002, 59, 217–226. [Google Scholar] [CrossRef]
- Kumar, N.; Gupta, U.; Jhala, Y.V.; Qureshi, Q.; Gosler, A.G.; Sergio, F. Habitat selection by an avian top predator in the tropical megacity of Delhi: Human activities and socio-religious practices as prey-facilitating tools. Urban Ecosyst. 2018, 21, 339–349. [Google Scholar] [CrossRef] [Green Version]
- Spelt, A.; Williamson, C.; Shamoun-Baranes, J.; Shepard, E.; Rock, P.; Windsor, S. Habitat use of urban-nesting lesser black-backed gulls during the breeding season. Sci. Rep. 2019, 9, 10527. [Google Scholar] [CrossRef]
- Maciusik, B.; Lenda, M.; Skórka, P. Corridors, local food resources, and climatic conditions affect the utilization of the urban environment by the Black-headed Gull Larus ridibundus in winter. Ecol. Res. 2010, 25, 263–272. [Google Scholar] [CrossRef]
- Garthe, S.; Schwemmer, P.; Paiva, V.H.; Corman, A.M.; Fock, H.O.; Voigt, C.C.; Adler, S. Terrestrial and Marine Foraging Strategies of an Opportunistic Seabird Species Breeding in the Wadden Sea. PLoS ONE 2016, 11, e0159630. [Google Scholar] [CrossRef] [Green Version]
- Lambert, L.; Cawkwell, F.; Holloway, P. The Importance of Connected and Interspersed Urban Green and Blue Space for Biodiversity: A Case Study in Cork City, Ireland. Geographies 2021, 1, 217–237. [Google Scholar] [CrossRef]
- Parsons, J. Cannibalism in Herring Gulls. Br. Birds 1971, 64, 524–537. [Google Scholar]
- Bergin, T.M. Habitat Selection by the Western Kingbird in Western Nebraska: A Hierarchical Analysis. Condor 1992, 94, 903–911. [Google Scholar] [CrossRef]
- Mateo-Tomás, P.; Olea, P.P. Combining scales in habitat models to improve conservation planning in an endangered vulture. Acta Oecol. 2009, 35, 489–498. [Google Scholar] [CrossRef]
- Fischer, J.H.; Debski, I.; Taylor, G.A.; Wittmer, H.U. Nest site selection of South Georgia Diving-petrels Pelecanoides georgicus on Codfish Island, New Zealand: Implications for conservation management. Bird Conserv. Int. 2017, 28, 216–227. [Google Scholar] [CrossRef]
- Bakken, L.E.; Bakken, G.S. American Redstart Feeding by Artificial Light. Ornithology 1977, 94, 373–374. [Google Scholar]
- Sohi, G.K.; Kler, T.K. Adaptations in avian nesting behavior in relation to indigenous trees and housing structures in Punjab. J. Entomol. Zool. Stud. 2017, 5, 1045–1051. [Google Scholar]
- Holveck, M.J.; Grégoire, A.; Doutrelant, C.; Lambrechts, M.M. Nest height is affected by lamppost lighting proximity in addition to nestbox size in urban great tits. J. Avian Biol. 2018, e01798. [Google Scholar] [CrossRef] [Green Version]
- Ben-Aharon, N.; Kapota, D.; Saltz, D. Roads and road-posts as an ecological trap for cavity nesting desert birds. Front. Conserv. Sci. 2020, 1, 614899. [Google Scholar] [CrossRef]
- Yorio, P.; Bertellotti, M.; Quintana, F. Preference for Covered Nest Sites and Breeding Success in Kelp Gulls Larus dominicanus. Mar. Ornithol. 1995, 23, 121–128. [Google Scholar]
- Kim, S.Y.; Monaghan, P. Effects of vegetation on nest microclimate and breeding performance of lesser black-backed gulls (Larus fuscus). J. Ornithol. 2005, 146, 176–183. [Google Scholar] [CrossRef]
- Monaghan, P. Aspects of the breeding biology of Herring Gulls Larus argentatus in urban colonies. Ibis 1979, 121, 475–481. [Google Scholar] [CrossRef]
- Roughan and O’Donovan Consulting Engineers. Nesting Gulls Populations in Balbriggan; Skerries and Howth, Co. Dublin Final Report to Fingal County Council: Dublin, Ireland, 2018. [Google Scholar]
- Johnson, D.H. The comparison of usage and availability measurements for evaluating resource preference. Ecology 1980, 61, 65–71. [Google Scholar] [CrossRef]
- Hutto, R.L. Habitat selection by nonbreeding migratory land birds. In Habitat Selection in Birds; Harcourt Brace Jovanovich: New York, NY, USA, 1985; pp. 455–476. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002; ISBN 0-387-95457-0. [Google Scholar]
- Long, J.A. jtools: Analysis and Presentation of Social Scientific Data. R Package Version 2.1.0. 2020. Available online: https://cran.r-project.org/package=jtools (accessed on 1 June 2021).
- Enners, L.; Schwemmer, P.; Corman, A.M.; Voigt, C.C.; Garthe, S. Intercolony variations in movement patterns and foraging behaviors among herring gulls (Larus argentatus) breeding in the eastern Wadden Sea. Ecol. Evol. 2018, 8, 7529–7542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tryjanowski, P.; Sparks, T.H.; Jerzak, L.; Rosin, Z.M.; Skórka, P. A Paradox for Conservation: Electricity Pylons May Benefit Avian Diversity in Intensive Farmland. Conserv. Lett. 2013, 7, 34–40. [Google Scholar] [CrossRef] [Green Version]
- Vogel, H.F.; McCarron, V.E.A.; Zocche, J.J. Use of artificial perches by birds in ecological restoration areas of the Cerrado and Atlantic Forest biomes in Brazil. Neotrop. Biol. Conserv. 2018, 13, 24–36. [Google Scholar]
- Calladine, J. A comparison of Herring Gull Larus argentatus and Lesser Black-backed Gull Larus fuscus nest sites: Their characteristics and relationships with breeding success. Bird Study 1997, 44, 318–326. [Google Scholar] [CrossRef]
- O’Hanlon, N.J.; Nager, R.G. What Herring Gulls Eat and How Diet Affects Their Breeding Performance. In Encyclopedia of Animal Behavior, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Van Donk, S.; Shamoun-Baranes, J.; Bouten, W.; Van Der Meer, J.; Camphuysen, K.C. Individual differences in foraging site fidelity are not related to time-activity budgets in Herring Gulls. Ibis 2019, 162, 429–445. [Google Scholar] [CrossRef]
- O’Hanlon, N.J.; Nager, R.G. Identifying habitat-driven spatial variation in colony size of Herring Gulls Larus argentatus. Bird Study 2018, 65, 306–316. [Google Scholar] [CrossRef] [Green Version]
- Goumas, M.; Boogert, N.J.; Kelley, L.A. Urban herring gulls use human behavioural cues to locate food. R. Soc. Open Sci. 2020, 7, 191959. [Google Scholar] [CrossRef] [Green Version]
- Rock, P. Urban gulls: Problems and solutions. Br. Birds 2005, 98, 338–355. [Google Scholar]
- Clark, D.E. Roosting, Site Fidelity, and Food Sources of Urban Gulls in Massachusetts: Implications for Protecting Public Water Supplies. Ph.D. Thesis, University of Massachusetts, Amherst, MA, USA, 2014. [Google Scholar]
- Pais de Faria, J.; Paiva, V.H.; Veríssimo, S.; Gonçalves, A.M.; Ramos, J.A. Seasonal variation in habitat use, daily routines and interactions with humans by urban-dwelling gulls. Urban Ecosyst. 2021, 24, 1101–1115. [Google Scholar] [CrossRef]
- Salas, R.; Muller, W.; Vercuijsse, H.; Lens, L.; Steinen, L. Forced nest site relocations negatively affect reproductive investment in a colonial seabird species. Biol. Conserv. 2020, 246, 108550. [Google Scholar] [CrossRef]
- Doubleday, K. Nonlinear liminality: Human-animal relations on preserving the world’s most famous tigress. Geoforum 2017, 81, 32–44. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| County | Howth | Balbriggan | Skerries | Bal 1 | Bal 2 | Bal 3 | Ske 1 | Ske 2 | Ske 3 | |
|---|---|---|---|---|---|---|---|---|---|---|
| (Intercept) | −2.69 ** | −1.95 ** | −2.20 ** | −1.37 | 140.89 ** | −35.89 ** | −9.27 ** | 193.80 ** | 599.17 | −28,730.00 |
| Roof | 1.49 ** | 1.49 ** | −9.67 * | −0.09 | −55.15 | 3.99 ** | −574.53 ** | 28,740.00 | ||
| Neighbour | 0.53 ** | −1.46 ** | −0.01 | 1.47 ** | −99.43 ** | 0.95 ** | 4.69 | 1.54 * | 361.26 | 1.07 * |
| Bin | 0.31 ** | 7.07 ** | −1.21 * | 2.18 ** | −49.88 ** | −7.18 * | 28.58 ** | 166.52 ** | −25,080.00 | |
| Streetlight | −0.17 ** | 0.19 | −0.14 | 0.46 | 55.65 ** | −17.28 ** | −0.39 | 1.54 * | 2472.00 | |
| Harbour | −1.39 ** | −0.52 | 2.10 * | −0.84 | −141.49 ** | −39.25 ** | −50.28 ** | −560.12 ** | −5.46 | |
| Coastline | 0.36 | 0.65 | −2.96 ** | 0.01 | −80.10 ** | 32.77 ** | 36.70 ** | 193.96 ** | 3.43 | |
| Greenspace | 0.01 ** | 0.00 | 0.00 | −0.03 ** | 2.03 ** | 0.22 * | −0.20 ** | −2.78 ** | −38.69 | −0.18 ** |
| Roof*Bin | −0.51 * | −3.78 ** | −536.19 ** | 25,080.00 | ||||||
| Roof*Harbour | 0.88 ** | −14.28 ** | −165.90 | 1665.78 ** | ||||||
| Roof*Greenspace | −0.01 ** | −0.01 ** | −0.04 ** | −0.05 ** | ||||||
| Roof*Neighbour | 2.57 ** | 2.01 ** | 9.04 ** | |||||||
| Roof*Streetlight | −1.23 ** | 0.94 * | 6.89 ** | −2479.00 | ||||||
| Roof*Coastline | 2.15 ** | 11.97 ** | −2.88 ** | 112.93 | −430.79 ** | |||||
| Neighbour*Bin | 0.38 ** | 0.77 ** | 0.58 ** | 37.29 ** | 10.05 * | |||||
| Neighbour*Harbour | 0.51 ** | −0.39* | 19.66 ** | |||||||
| Neighbour*Coastline | −0.23 ** | 0.53 ** | 55.70 ** | 14.36 * | ||||||
| Neighbour*Greenspace | −0.01 ** | −0.11 ** | −0.09 ** | −0.11 ** | ||||||
| Neighbour*Streetlight | 0.40 | 4.66 * | −2.70 ** | 7.75 ** | ||||||
| Bin*Harbour | −0.31 | 6.89 | −0.98 ** | −3.47 ** | ||||||
| Bin*Coastline | −0.68 ** | −6.81 ** | 12.30 ** | |||||||
| Bin*Streetlight | −0.22 * | −32.45 ** | −5.04 ** | −7.24 ** | ||||||
| Bin*Greenspace | 0.02 ** | 0.02 ** | −0.57 ** | 0.07 * | 0.08 | −2.42 ** | -0.06 | |||
| Streetlight*Harbour | −0.32 ** | −0.96 ** | −50.99 ** | −19.71 ** | ||||||
| Streetlight*Coastline | −0.40 ** | 12.63 ** | −6.33 ** | −8.14 ** | ||||||
| Streetlight*Greenspace | 0.02 * | 0.04 ** | −0.02 * | |||||||
| Harbour*Coastline | −0.20 ** | 1.67 ** | 2.20 * | 59.45 ** | ||||||
| Harbour*Greenspace | 0.00 ** | −0.04 ** | 0.02 ** | 0.24 * | 0.34 ** | 8.17 ** | 0.06 | |||
| Coastline*Greenspace | 0.00 * | 0.03 ** | −0.82 ** | −0.21 * | −2.76 ** | −0.08 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dalla Pria, C.; Cawkwell, F.; Newton, S.; Holloway, P. City Living: Nest-Site Selection Preferences in Urban Herring Gulls, Larus argentatus. Geographies 2022, 2, 161-172. https://doi.org/10.3390/geographies2020011
Dalla Pria C, Cawkwell F, Newton S, Holloway P. City Living: Nest-Site Selection Preferences in Urban Herring Gulls, Larus argentatus. Geographies. 2022; 2(2):161-172. https://doi.org/10.3390/geographies2020011
Chicago/Turabian StyleDalla Pria, Caitlin, Fiona Cawkwell, Stephen Newton, and Paul Holloway. 2022. "City Living: Nest-Site Selection Preferences in Urban Herring Gulls, Larus argentatus" Geographies 2, no. 2: 161-172. https://doi.org/10.3390/geographies2020011
APA StyleDalla Pria, C., Cawkwell, F., Newton, S., & Holloway, P. (2022). City Living: Nest-Site Selection Preferences in Urban Herring Gulls, Larus argentatus. Geographies, 2(2), 161-172. https://doi.org/10.3390/geographies2020011