
Evaluation of Muscle Synergy Flexibility Induced by a Muscle Nerve Block in Chronic Stroke Patients


Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Intervention
2.3. Motion Analysis
2.4. Kinematic Variables
2.5. EMG Variables
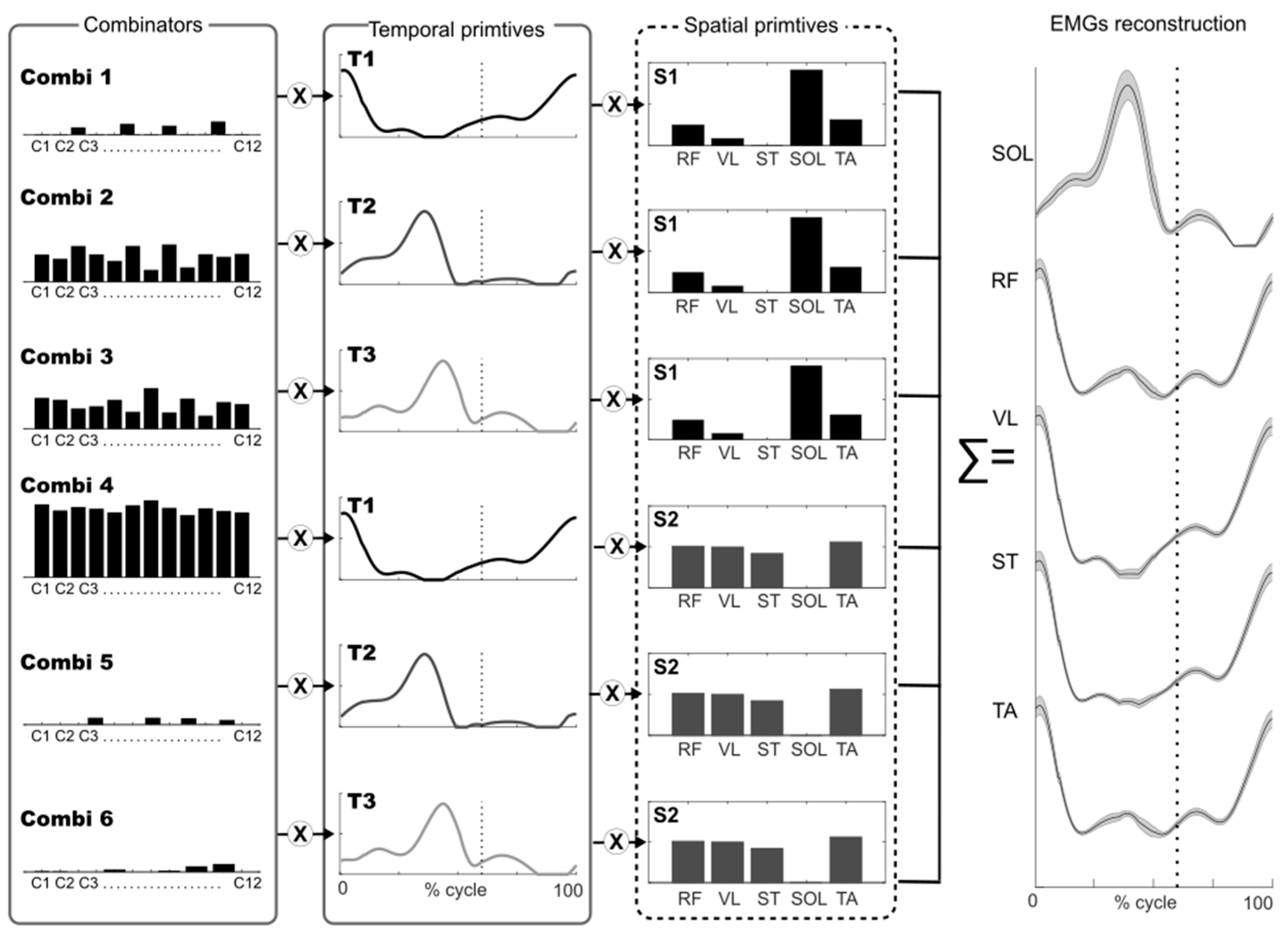
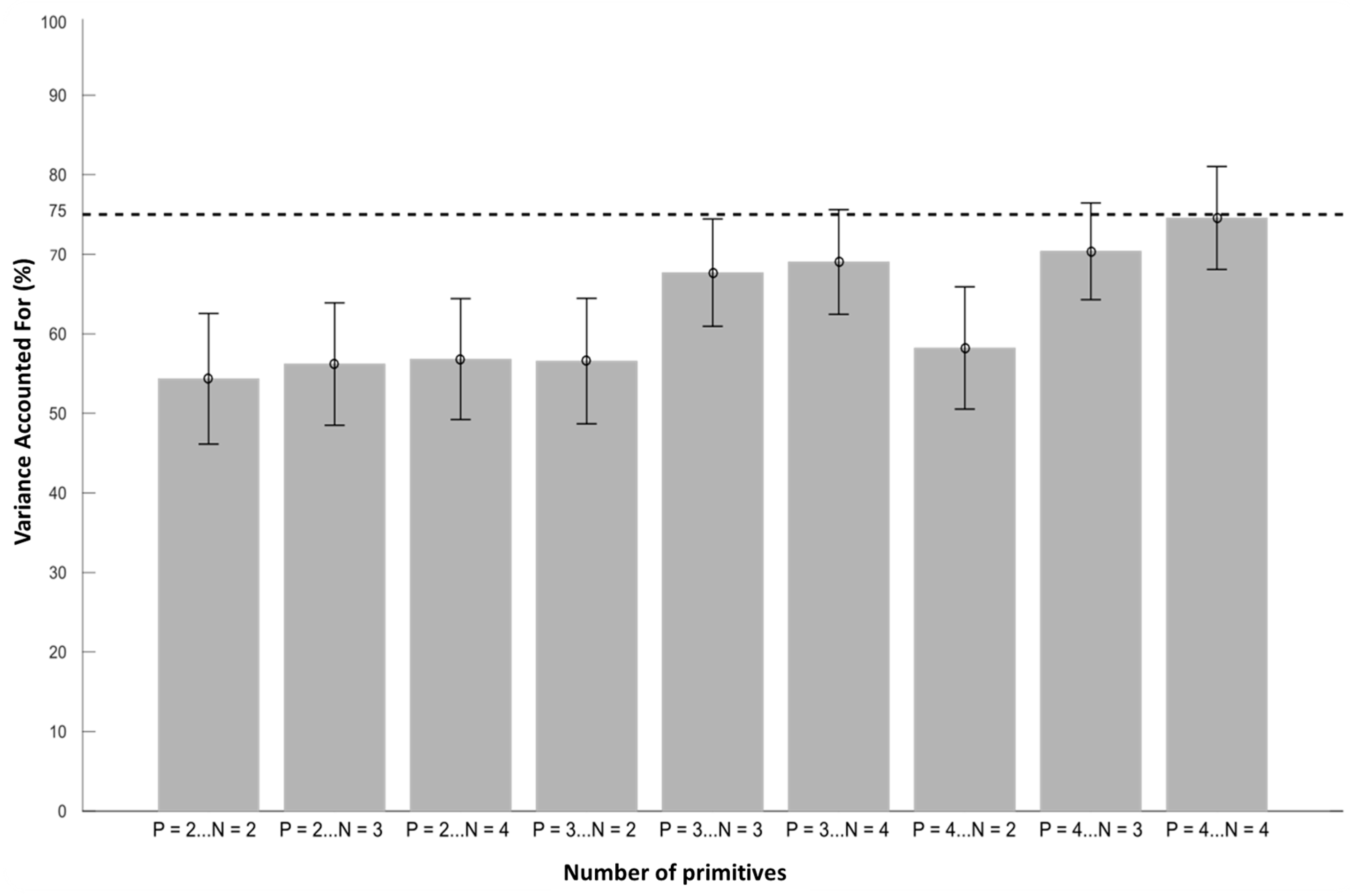
2.6. Muscle Synergy Extraction
2.7. Muscle Synergy Indexes
2.8. Statistical Analyses
3. Results
3.1. Gait Variables
3.2. Muscle Synergy Extraction
3.3. Muscle Synergy Indexes
4. Discussion
4.1. Gait Variables
4.2. Muscle Synergy Indexes
4.3. Clinical Implications
4.4. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hashiguchi, Y.; Ohata, K.; Kitatani, R.; Yamakami, N.; Sakuma, K.; Osako, S.; Aga, Y.; Watanabe, A.; Yamada, S. Merging and Fractionation of Muscle Synergy Indicate the Recovery Process in Patients with Hemiplegia: The First Study of Patients after Subacute Stroke. Neural Plast. 2016, 2016, 5282957. [Google Scholar] [CrossRef] [PubMed]
- Routson, R.L.; Clark, D.J.; Bowden, M.G.; Kautz, S.A.; Neptune, R.R. The influence of locomotor rehabilitation on module quality and post-stroke hemiparetic walking performance. Gait Posture 2013, 38, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Safavynia, S.; Torres-Oviedo, G.; Ting, L. Muscle Synergies: Implications for Clinical Evaluation and Rehabilitation of Movement. Top. Spinal Cord Inj. Rehabil. 2011, 17, 16–24. [Google Scholar] [CrossRef]
- Van Criekinge, T.; Vermeulen, J.; Wagemans, K.; Schröder, J.; Embrechts, E.; Truijen, S.; Hallemans, A.; Saeys, W. Lower limb muscle synergies during walking after stroke: A systematic review. Disabil Rehabil. 2020, 42, 2836–2845. [Google Scholar] [CrossRef]
- Booth, A.T.C.; van der Krogt, M.M.; Harlaar, J.; Dominici, N.; Buizer, A.I. Muscle synergies in response to biofeedback-driven gait adaptations in children with cerebral palsy. Front. Physiol. 2019, 10, 1208. [Google Scholar] [CrossRef]
- Mileti, I.; Zampogna, A.; Santuz, A.; Asci, F.; Del Prete, Z.; Arampatzis, A.; Palermo, E.; Suppa, A. Muscle synergies in parkinson’s disease. Sensors 2020, 20, 3209. [Google Scholar] [CrossRef]
- Shuman, B.R.; Schwartz, M.H.; Steele, K.M. Electromyography data processing impacts muscle synergies during gait for unimpaired children and children with cerebral palsy. Front. Comput. Neurosci. 2017, 11, 50. [Google Scholar] [CrossRef]
- Ambrosini, E.; Parati, M.; Peri, E.; De Marchis, C.; Nava, C.; Pedrocchi, A.; Ferriero, G.; Ferrante, S. Changes in leg cycling muscle synergies after training augmented by functional electrical stimulation in subacute stroke survivors: A pilot study. J. Neuroeng. Rehabil. 2020, 27, 17–35. [Google Scholar] [CrossRef]
- Pollock, C.; Eng, J.; Garland, S. Clinical measurement of walking balance in people post stroke: A systematic review. Clin. Rehabil. 2011, 25, 693–708. [Google Scholar] [CrossRef]
- Langhorne, P.; Coupar, F.; Pollock, A. Motor recovery after stroke: A systematic review. Lancet Neurol. 2009, 8, 741–754. [Google Scholar] [CrossRef]
- Weerdesteyn, V.; de Niet, M.; van Duijnhoven, H.J.; Geurts, A.C. Falls in individuals with stroke. J. Rehabil. Res. Dev. 2008, 45, 1195–1213. [Google Scholar] [CrossRef]
- Goldberg, S.R.; Anderson, F.C.; Pandy, M.G.; Delp, S.L. Muscles that influence knee flexion velocity in double support: Implications for stiff-knee gait. J. Biomech. 2004, 37, 1189–1196. [Google Scholar] [CrossRef]
- Kerrigan, D.C.; Bang, M.S.; Burke, D.T. An algorithm to assess stiff-legged gait in traumatic brain injury. J. Head Trauma Rehabil. 1999, 14, 136–145. [Google Scholar] [CrossRef]
- Piazza, S.J.; Delp, S.L. The influence of muscles on knee flexion during the swing phase of gait. J. Biomech. 1996, 29, 723–733. [Google Scholar] [CrossRef]
- Riley, P.O.; Kerrigan, D.C. Torque action of two-joint muscles in the swing period of stiff-legged gait: A forward dynamic model analysis. J. Biomech. 1998, 31, 835–840. [Google Scholar] [CrossRef]
- Roche, N.; Bonnyaud, C.; Geiger, M.; Bussel, B.; Bensmail, D. Relationship between hip flexion and ankle dorsiflexion during swing phase in chronic stroke patients. Clin. Biomech. 2015, 30, 219–225. [Google Scholar] [CrossRef]
- Sung, D.H.; Bang, H.J. Motor branch block of the rectus femoris: Its effectiveness in stiff-legged gait in spastic paresis. Arch. Phys. Med. Rehabil. 2000, 81, 910–915. [Google Scholar] [CrossRef]
- Robertson, J.V.G.; Pradon, D.; Bensmail, D.; Fermanian, C.; Bussel, B.; Roche, N. Relevance of botulinum toxin injection and nerve block of rectus femoris to kinematic and functional parameters of stiff knee gait in hemiplegic adults. Gait Posture 2009, 29, 108–112. [Google Scholar] [CrossRef]
- Bohannon, R.W.; Smith, M.B. Interrater reliability of a modified Ashworth scale of muscle spasticity. Phys. Ther. 1987, 67, 206–207. [Google Scholar] [CrossRef]
- Merlo, A.; Campanini, I. Impact of instrumental analysis of stiff knee gait on treatment appropriateness and associated costs in stroke patients. Gait Posture 2019, 72, 195–201. [Google Scholar] [CrossRef]
- Kadaba, M.P.; Ramakrishnan, H.K.; Wootten, M.E. Measurement of lower extremity kinematics during level walking. J. Orthop. Res. 1990, 8, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Winter, D.A.; Yack, H.J. EMG profiles during normal human walking: Stride-to-stride and inter-subject variability. Electroencephalogr. Clin. Neurophysiol. 1987, 67, 402–411. [Google Scholar] [CrossRef]
- Devarajan, K.; Cheung, V.C.K. On non-negative matrix factorization algorithms for signal-dependent noise with application to electromyography data. Neural Comput. 2014, 26, 1128–1168. [Google Scholar] [CrossRef]
- Santuz, A.; Ekizos, A.; Janshen, L.; Baltzopoulos, V.; Arampatzis, A. The Influence of Footwear on the Modular Organization of Running. Front. Physiol. 2017, 8, 958. [Google Scholar] [CrossRef]
- Hebenstreit, F.; Leibold, A.; Krinner, S.; Welsch, G.; Lochmann, M.; Eskofier, B.M. Effect of walking speed on gait sub phase durations. Hum. Mov. Sci. 2015, 43, 118–124. [Google Scholar] [CrossRef]
- Perry, J.; Burnfield, J.M. Gait Analysis: Normal and Pathological Function, 2nd ed.; SLACK: Thorofare, NJ, USA, 1992. [Google Scholar]
- Tresch, M.C.; Cheung, V.C.; D’Avella, A. Matrix factorization algorithms for the identification of muscle synergies: Evaluation on simulated and experimental data sets. J. Neurophysiol. 2006, 95, 2199–2212. [Google Scholar] [CrossRef]
- Delis, I.; Panzeri, S.; Pozzo, T.; Berret, B. A unifying model of concurrent spatial and temporal modularity in muscle activity. J. Neurophysiol. 2014, 111, 675–693. [Google Scholar] [CrossRef]
- Lee, D.D.; Seung, H.S. Algorithms for non-negative matrix factorization. In Advances in Neural Information Processing Systems 13—Proceedings of the 2000 Conference, Neural Information Processing Systems Foundation, Denver, CO, USA, 27–30 November 2000; MIT Press: Cambridge, MA, USA, 2000. [Google Scholar]
- Hayes, H.B.; Chvatal, S.A.; French, M.A.; Ting, L.H.; Trumbower, R.D. Neuromuscular constraints on muscle coordination during overground walking in persons with chronic incomplete spinal cord injury. Clin. Neurophysiol. 2014, 125, 2024–2035. [Google Scholar] [CrossRef]
- Hinnekens, E.; Berret, B.; Morard, E.; Do, M.C.; Barbu-Roth, M.; Teulier, C. Optimization of modularity during development to simplify walking control across multiple steps. Front. Neural Circuits 2024, 17, 1340298. [Google Scholar] [CrossRef]
- Hinnekens, E.; Barbu-Roth, M.; Do, M.C.; Berret, B.; Teulier, C. Generating variability from motor primitives during infant locomotor development. Elife 2023, 12, e87463. [Google Scholar] [CrossRef]
- Hoyer, P.O. Non-negative matrix factorization with sparseness constraints. J. Mach. Learn. Res. 2004, 5, 1457–1469. [Google Scholar]
- Oliveira, A.S.; Gizzi, L.; Farina, D.; Kersting, U.G. Motor modules of human locomotion: Influence of EMG averaging, concatenation, and number of step cycles. Front. Hum. Neurosci. 2014, 8, 335. [Google Scholar] [CrossRef] [PubMed]
- Clark, D.J.; Ting, L.H.; Zajac, F.E.; Neptune, R.R.; Kautz, S. Merging of Healthy Motor Modules Predicts Reduced Locomotor Performance and Muscle Coordination Complexity Post-Stroke. J. Neurophysiol. 2010, 103, 844–857. [Google Scholar] [CrossRef] [PubMed]
- Kargo, W.J.; Nitz, D.A. Early skill learning is expressed through selection and tuning of cortically represented muscle synergies. J. Neurosci. 2003, 23, 11255–11269. [Google Scholar] [CrossRef]
- Torres-Oviedo, G.; Bastian, A.J. Natural error patterns enable transfer of motor learning to novel contexts. J. Neurophysiol. 2012, 107, 346–356. [Google Scholar] [CrossRef]
- Frère, J.; Hug, F. Between-subject variability of muscle synergies during a complex motor skill. Front. Comput. Neurosci. 2012, 6, 99. [Google Scholar] [CrossRef]
- Ririe, D.G.; Walker, F.O.; James, R.L.; Butterworth, J. Effect of alkalinization of lidocaine on median nerve block. Br. J. Anaesth. 2000, 84, 163–168. [Google Scholar] [CrossRef]
- Santuz, A.; Ekizos, A.; Eckardt, N.; Kibele, A.; Arampatzis, A. Challenging human locomotion: Stability and modular organisation in unsteady conditions. Sci. Rep. 2018, 8, 2740. [Google Scholar] [CrossRef]
- Oliveira, A.S.C.; Gizzi, L.; Kersting, U.G.; Farina, D. Modular organization of balance control following perturbations during walking. J. Neurophysiol. 2012, 108, 1895–1906. [Google Scholar] [CrossRef]
- Den Otter, A.R.; Geurts, A.C.H.; De Haart, M.; Mulder, T.; Duysens, J. Step characteristics during obstacle avoidance in hemiplegic stroke. Exp. Brain Res. 2005, 161, 180–192. [Google Scholar] [CrossRef]
- van Swigchem, R.; van Duijnhoven, H.J.R.; den Boer, J.; Geurts, A.C.; Weerdesteyn, V. Deficits in motor response to avoid sudden obstacles during gait in functional walkers poststroke. Neurorehabil. Neural Repair. 2013, 27, 230–239. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Participants | Sex | Age | Height | Weight | Pathology |
|---|---|---|---|---|---|
| P 1 | M | 27 | 180 | 74 | Left Hemiparesis |
| P 2 | M | 40 | 188 | 73 | Left Hemiparesis |
| P 3 | F | 52 | 162 | 68 | Right Hemiparesis |
| P 4 | M | 33 | 180 | 85 | Left Hemiparesis |
| P 5 | M | 37 | 182 | 82 | Right Hemiparesis |
| P 6 | M | 40 | 173 | 76 | Left Hemiparesis |
| P 7 | M | 40 | 163 | 51 | Left Hemiparesis |
| P 8 | F | 58 | 160 | 69 | Right Hemiparesis |
| Maximum Knee Flexion (degrees) | Gait Speed (m·s−1) | Step Length (cm) | ||||
|---|---|---|---|---|---|---|
| PRE | POST | PRE | POST | PRE | POST | |
| P 1 | 35.09 ± 2.47 | 43.13 ± 3.62 | 0.54 ± 0.11 | 0.66 ± 0.05 | 30.5 ± 2.61 | 36.23 ± 2.77 |
| P 2 | 24.07 ± 1.35 | 29.55 ± 1.59 | 0.57 ± 0.07 | 0.67 ± 0.04 | 44.70 ± 5.27 | 44.49 ± 5.52 |
| P 3 | 18.02 ± 2.06 | 23.05 ± 3.37 | 0.57 ± 0.04 | 0.54 ± 0.05 | 45.19 ± 2.98 | 45.64 ± 3.17 |
| P 4 | 28.83 ± 3.04 | 33.90 ± 4.74 | 0.47 ± 0.04 | 0.58 ± 0.10 | 40.87 ± 3.09 | 43.81 ± 7.46 |
| P 5 | 24.77 ± 2.98 | 43.63 ± 6.91 | 0.42 ± 0.05 | 0.30 ± 0.02 | 36.99 ± 4.40 | 25.35 ± 2.25 |
| P 6 | 24.79 ± 2.12 | 38.00 ± 3.91 | 0.82 ± 0.04 | 0.90 ± 0.05 | 56.10 ± 2.78 | 55.06 ± 3.83 |
| P 7 | 26.47 ± 3.62 | 40.20 ± 5.14 | 0.50 ± 0.04 | 0.64 ± 0.07 | 44.99 ± 3.45 | 45.21 ± 4.18 |
| P 8 | 31.42 ± 1.87 | 33.35 ± 2.90 | 0.44 ± 0.03 | 0.46 ± 0.06 | 36.10 ± 2.61 | 37.04 ± 2.99 |
| Mean | 26.68 ± 5.16 | 35.60 ± 7.07 ** | 0.54 ± 0.12 | 0.59 ± 0.17 | 41.93 ± 7.73 | 41.61 ± 8.76 |
| Effect size | 1.18 | 0.33 | 0.04 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Supiot, A.; Roche, N.; Berret, B.; Pradon, D. Evaluation of Muscle Synergy Flexibility Induced by a Muscle Nerve Block in Chronic Stroke Patients. Biomechanics 2025, 5, 27. https://doi.org/10.3390/biomechanics5020027
Supiot A, Roche N, Berret B, Pradon D. Evaluation of Muscle Synergy Flexibility Induced by a Muscle Nerve Block in Chronic Stroke Patients. Biomechanics. 2025; 5(2):27. https://doi.org/10.3390/biomechanics5020027
Chicago/Turabian StyleSupiot, Anthony, Nicolas Roche, Bastien Berret, and Didier Pradon. 2025. "Evaluation of Muscle Synergy Flexibility Induced by a Muscle Nerve Block in Chronic Stroke Patients" Biomechanics 5, no. 2: 27. https://doi.org/10.3390/biomechanics5020027
APA StyleSupiot, A., Roche, N., Berret, B., & Pradon, D. (2025). Evaluation of Muscle Synergy Flexibility Induced by a Muscle Nerve Block in Chronic Stroke Patients. Biomechanics, 5(2), 27. https://doi.org/10.3390/biomechanics5020027