
Marine Apartectal (Chamberless) Mastogloiaceae (Diatomeae: Bacillariales): Paramastogloia cubana gen. nov., sp. nov., New Observations and Emended Diagnosis of Mastoneis, and Comparison with Mastogloiopsis


Abstract
1. Introduction
2. Materials and Methods
3. Results
- Mastogloiales D.G.Mann & Round
- Mastogloiaceae Mereschkowsky
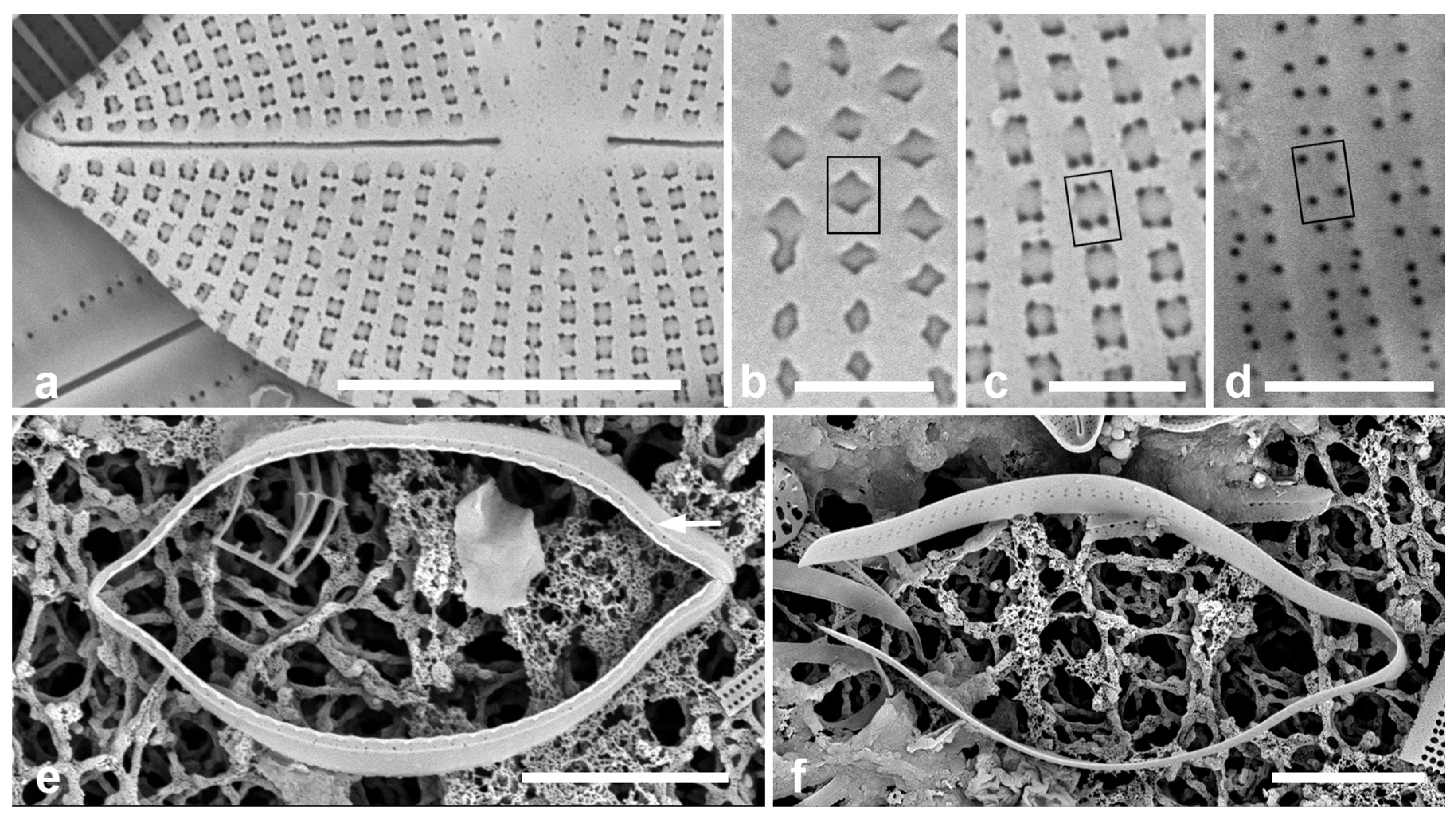
- Paramastogloia gen. nov.



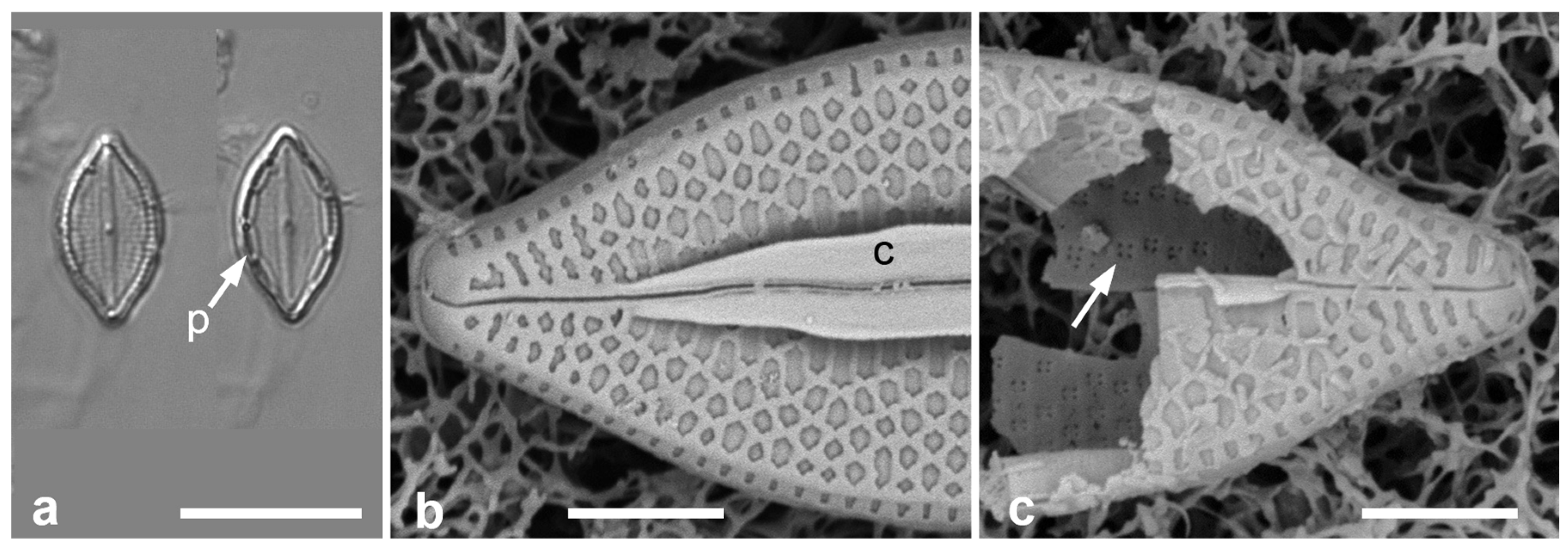
- Additional Observation of Mastogloiopsis biseriata. Figure 5a,b.

4. Discussion
4.1. Paramastogloia
4.2. Diagnosis of Mastoneis
5. Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smol, J.P.; Stoermer, E.F. (Eds.) The Diatoms: Applications for the Environmental and Earth Sciences, 2nd ed.; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Hustedt, F. Die Kieselalgen Deutschlands, Österreichs und der Schweiz. In Rabenhorst’s Kryptogamenflora; Band 7, Teil 2, Lief. 4; Akademischen Verlagsgesellschaft: Frankfurt am Main, Germany, 1933; Johnson Reprint, New York, NY, USA, 1962. [Google Scholar]
- Paddock, T.B.B.; Kemp, K.D. An illustrated survey of the morphological features of the diatom genus Mastogloia. Diatom Res. 1990, 5, 73–103. [Google Scholar] [CrossRef]
- Round, F.E.; Crawford, R.M.; Mann, D.G. The Diatoms: Biology and Morphology of the Genera; Cambridge University Press: Cambridge, UK, 1990. [Google Scholar]
- Lobban, C.S. Towards a biogeography of Mastogloia (Diatomeae: Bacillariophyceae): A preliminary checklist for Yap, Micronesia, with new ultrastructure images and a review of problems. Nova Hedw. 2025, 120, 313–417. [Google Scholar] [CrossRef]
- Ross, R.; Cox, E.J.; Karayeva, N.I.; Mann, D.G.; Paddock, T.B.B.; Simonsen, R.; Sims, P.A. An amended terminology for the siliceous components of the diatom cell. Nova Hedw. Beih. 1979, 64, 513–533. [Google Scholar]
- Sterrenburg, F.A.S.; Tiffany, M.A.; Meave del Castillo, M.E. Valve morphogenesis in the diatom genus Pleurosigma W. Smith (Bacillariophyceae): Nature’s alternative sandwich. J. Nanosci. Nanotech. 2005, 5, 140–145. [Google Scholar] [CrossRef]
- Sterrenburg, F.A.S.; Tiffany, M.A.; Hinz, F.; Herwig, W.E.; Hargraves, P.E. Seven new species expand the morphological spectrum of Haslea. A comparison with Gyrosigma and Pleurosigma (Bacillariophyta). Phytotaxa 2015, 207, 143–162. [Google Scholar] [CrossRef]
- Lobban, C.S.; Perez, C.O.; Ashworth, M.P. Non-blue Haslea species (Bacillariophyceae: Naviculaceae) in the benthic marine flora of Guam (Mariana Islands, Western Pacific Ocean). Diatom Res. 2020, 35, 163–183. [Google Scholar] [CrossRef]
- Mironov, A.; Glushchenko, A.; Kezlya, E.; Maltsev, Y.; Iurmanov, A.; Liu, Y.; Kulikovskiy, M. Decussiphycus sinensis sp. nov. (Bacillariophyceae, Mastogloiales)—A new species described from China, with comments on phylogenetic position of the genus. PhytoKeys 2025, 254, 1–19. [Google Scholar] [CrossRef]
- Cox, E.J. Diatoms, Diatomeae (Bacillariophyceae s.l., Bacillariophyta). In Engler’s Syllabus of Plant Families, 13th ed.; Frey, W., Ed.; Borntraeger Science Publishers: Stuttgart, Germany, 2015; Part 2/1; pp. 64–103. [Google Scholar]
- Guiry, M.D.; Guiry, G.M. AlgaeBase; World-wide Electronic Publication, National University of Ireland: Galway, Ireland. Available online: https://www.algaebase.org (accessed on 16 February 2025).
- Gluschenko, A.; Kulikovskiy, M.; Okhapkin, A.; Kociolek, J.P. Aneumastus laosica sp. nov. and A. genkalii sp. nov.—Two new diatom species from Laos (Southeast Asia) with comments on the biogeography of the genus. Cryptogam. Algol. 2017, 38, 183–199. [Google Scholar] [CrossRef]
- Spaulding, S.; Potapova, M.; Bishop, I.W.; Lee, S.S.; Gasperak, T.S.; Jovanoska, E.; Furey, P.C.; Edlund, M.B. Diatoms.org: Supporting taxonomists, connecting communities. Diatom Res. 2021, 36, 291–304. [Google Scholar] [CrossRef]
- Stelbrink, B.; Jovanovska, E.; Levkov, Z.; Ognjanova-Rumenova, N.; Wilke, T.; Albrech, C. Diatoms do radiate: Evidence for a freshwater species flock. J. Evol. Biol. 2018, 31, 1969–1975. [Google Scholar] [CrossRef]
- Edlund, M.B.; Brant, L.A.; Levkov, Z.; Nakov, T. An emended description of Decussata (Patrick) Lange-Bertalot & Metzeltin that includes protoplast organization and detailed valve and cingulum ultrastructure. Diatom Res. 2006, 21, 269–280. [Google Scholar]
- Guiry, M.D.; Ghandi, K. Decussiphycus gen. nov.: A validation of “Decussata” (R.M.Patrick) Lange-Bertalot (Mastogloiaceae, Bacillariophyta). Not. Alg. 2019, 94, 2. [Google Scholar]
- Cleve, P.T. Synopsis of the naviculoid diatoms, Part I. Kongliga Sven.-Vetensk. Akad. Handl. 1894, 26, 1–194. [Google Scholar]
- Lobban, C.S.; Navarro, J.N. Mastogloiopsis biseriata, gen. et sp. nov., a diatom without partecta, is very similar to the Marginulatae group of Mastogloia. Nova Hedw. 2012, 94, 251–263. [Google Scholar] [CrossRef]
- Grunow, A. Über einige neue und ungenügend bekannte Arten und Gattungen von Diatomaceen. Verhandl. Kaiser. König. Zool. Bot. Ges. Wien 1863, 13, 137–162, 2 pls. [Google Scholar]
- Siqueiros Beltrones, D.A.; Echevarría Herrera, E.; López-Fuerte, F.O.; Martínez, Y.J. Species diversity of benthic marine diatoms from a Natural Protected Area in Cuba. Diversity 2025, 17, 181. [Google Scholar] [CrossRef]
- Torgan, L.C.; Donadel, L.; Gonçalves da Silva, J. A transferência de Navicula sovereignae Hustedt para o gênero Placoneis Mereschkowsky (Bacillariophyta). Iheringia Ser. Bot. 2010, 65, 107–114. [Google Scholar]
- Siqueiros Beltrones, D.A.; Echevarría Herrera, E.; López-Fuerte, F.O. Additions to the marine Mastogloia (Bacillariophyceae) from Cuban coasts; remarks on misidentified taxa. Diversity 2024, 16, 747. [Google Scholar] [CrossRef]
- Turland, N.J.; Wiersema, J.H.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.-H.; Li, D.-Z.; Marhold, K.; et al. (Eds.) International Code of Nomenclature for algae, fungi, and plants. (Shenzhen Code) adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017. Regnum Veg. 2018, 159, [i]–xxxviii, 1–253. Available online: https://www.iapt-taxon.org/historic/2018.htm (accessed on 6 March 2025).
- von Stosch, H.A. An amended terminology of the diatom girdle. Nova Hedw. Beih. 1975, 53, 1–28. [Google Scholar]
- Mann, A. Marine diatoms of the Philippine Islands. Bull. United States Natl. Mus. 1925, 100 Pt 1, 1–182. [Google Scholar]
- Hendey, N.I. Some littoral diatoms of Kuwait. Nova Hedw. Beih. 1970, 31, 101–167. [Google Scholar] [CrossRef]
- Witkowski, A.; Lange-Bertalot, H.; Metzeltin, D. Diatom Flora of Marine Coasts; I.A.R.G. Gantner Verlag: Ruggell, Germany, 2000. [Google Scholar]
- Hein, M.K.; Winsborough, B.M.; Sullivan, M.J. Bacillariophyta (Diatoms) of the Bahamas; Iconographia Diatomologica; Gantner Verlag: Ruggell, Germany, 2008. [Google Scholar]
- Güttinger, W. Collection of SEM-Micrographs of Diatoms. Series 2. 50 Plates; Koeltz Scientific Books: Koenigstein, Germany, 1988. [Google Scholar]
- Paddock, T.B.B.; Kemp, K.-D. A description of some new species of the genus Mastogloia with further observations on M. elegans and M. goessii. Diatom Res. 1988, 3, 109–121. [Google Scholar] [CrossRef]
- Voigt, M. Contribution to the knowledge of the diatom genus Mastogloia. J. Roy. Microsc. Soc. Ser. 3 1942, 62, 1–20. [Google Scholar] [CrossRef]
- Wallich, G.C. On the siliceous organisms found in the digestive cavities of the Salpae, and their relation to the flint nodules of the Chalk Formation. Trans. Microsc. Soc. Lond. N. Ser. 1860, 8, 36–55, pl. II. [Google Scholar]
- Kemp, K.-D.; Paddock, T.B.B. Observations on the diatom genus Stigmaphora G. C. Wallich and two species of Mastogloia Thwaites ex Wm Smith. In Proceedings of the 9th International Diatom Symposium, Bristol, UK, 24–30 August 1986; Round, F.E., Ed.; Koenigstein & Biopress Ltd.: Bristol, UK, 1988; pp. 405–416. [Google Scholar]
- Molinari Novoa, E.A. In Guiry, M.D. & Guiry, G.M. 2024. AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. Available online: https://www.algaebase.org/search/genus/detail/?genus_id=47079 (accessed on 20 February 2025).
- Schuster, T.M.; Hamilton, P.B.; Haring, V.; Edlund, M.B.; Van de Vijver, B. An introduction to the catalogue of Albert Grunow’s 19th century diatom collection at W including a palaeographic aid. Ann. Nat. Hist. Mus. Vienna B 2023, 125, 101–122. [Google Scholar]
- Lobban, C.S. Disymmetria reticulata, gen. nov., sp. nov. (Mediophyceae: Thalassiosirales), a new genus in Lauderiaceae emend., and transfer of Lauderia excentrica. Diatom 2023, 39, 25–30. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Character | Paramastogloia | Mastoneis | Mastogloiopsis | Mastogloia | Aneumastus | Decussiphycus |
|---|---|---|---|---|---|---|
| Plastids | Unknown | Two, fore-and-aft, with pyrenoid | Two, fore-and-aft, with pyrenoid | Two, fore-and-aft, with pyrenoid | Two, H-shaped, fore-and-aft, girdle-appressed, with pyrenoid | Fore-and-aft, complex lobed, no pyrenoid |
| Structured mucilage | Unknown | None; motile in biofilm on sand | Polar stalk, attached to filamentous seaweed | When present, mostly as threads from partecta, some spp. with polar or central stalks from valves, some in mucilage bubbles | Absent | Absent |
| Valve wall | Pseudoloculate | Pseudoloculate | Laminar | Some spp. pseudoloculate, some laminate; still others loculate or alveolate | Pseudoloculate | Laminar |
| Areolae | External broad, irregular foramen internal cluster of 4 pores | External small pore, internal cluster of 4 pores | Small pores directly through the wall | Diverse | External variable, internal cribra | External pores, internal occulsions, internal hymenes |
| Internal transapical costae | Absent | Present in outer zone, every 2nd or 3rd virga | Present across entire valve, every virga | Some spp. (outer zone or entire), every virga | Absent | Absent |
| Valvocopula pores (pars exterior) | None | None | Densely porate | Most spp. none | Irregular row of poroids | Two rows of areolae |
| Partecta on valvocopula/1st pleura | No | No | Porate hollow flange on valvocopula | Yes | Series of tiny chambers | No |
| References | This paper | This paper | Lobban & Navarro [19] | Paddock & Kemp [3], Lobban [5] | Gluschenko et al. [13] | Edlund et al. [16]; Mironov et al. [10] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lobban, C.S. Marine Apartectal (Chamberless) Mastogloiaceae (Diatomeae: Bacillariales): Paramastogloia cubana gen. nov., sp. nov., New Observations and Emended Diagnosis of Mastoneis, and Comparison with Mastogloiopsis. Taxonomy 2025, 5, 24. https://doi.org/10.3390/taxonomy5020024
Lobban CS. Marine Apartectal (Chamberless) Mastogloiaceae (Diatomeae: Bacillariales): Paramastogloia cubana gen. nov., sp. nov., New Observations and Emended Diagnosis of Mastoneis, and Comparison with Mastogloiopsis. Taxonomy. 2025; 5(2):24. https://doi.org/10.3390/taxonomy5020024
Chicago/Turabian StyleLobban, Christopher S. 2025. "Marine Apartectal (Chamberless) Mastogloiaceae (Diatomeae: Bacillariales): Paramastogloia cubana gen. nov., sp. nov., New Observations and Emended Diagnosis of Mastoneis, and Comparison with Mastogloiopsis" Taxonomy 5, no. 2: 24. https://doi.org/10.3390/taxonomy5020024
APA StyleLobban, C. S. (2025). Marine Apartectal (Chamberless) Mastogloiaceae (Diatomeae: Bacillariales): Paramastogloia cubana gen. nov., sp. nov., New Observations and Emended Diagnosis of Mastoneis, and Comparison with Mastogloiopsis. Taxonomy, 5(2), 24. https://doi.org/10.3390/taxonomy5020024