
A New Species of Proctoporus (Reptilia, Gymnophthalmidae, Cercosaurinae) from the Puna of the Otishi National Park in Peru †

 ,
,  ,
, 
Abstract
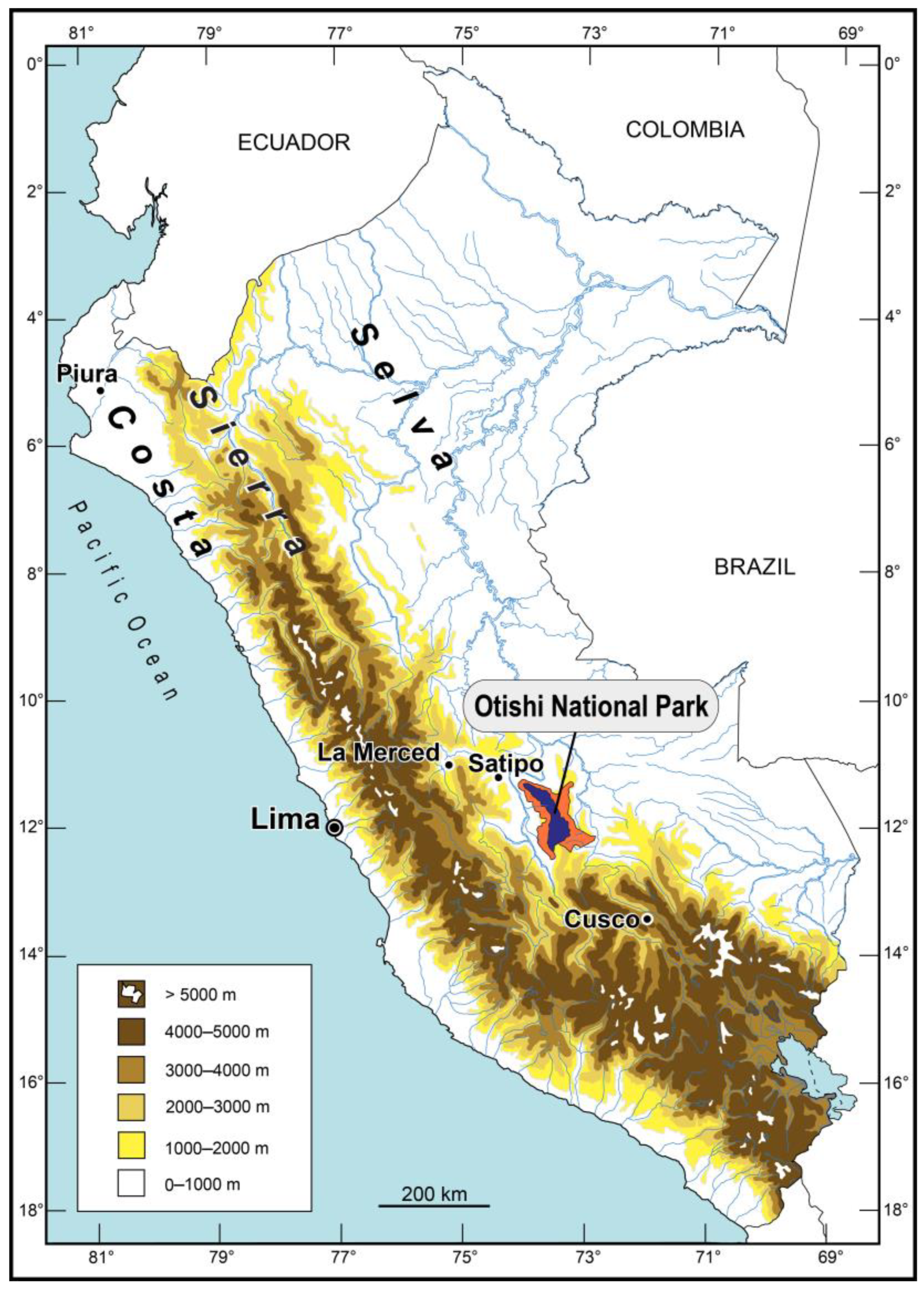
1. Introduction
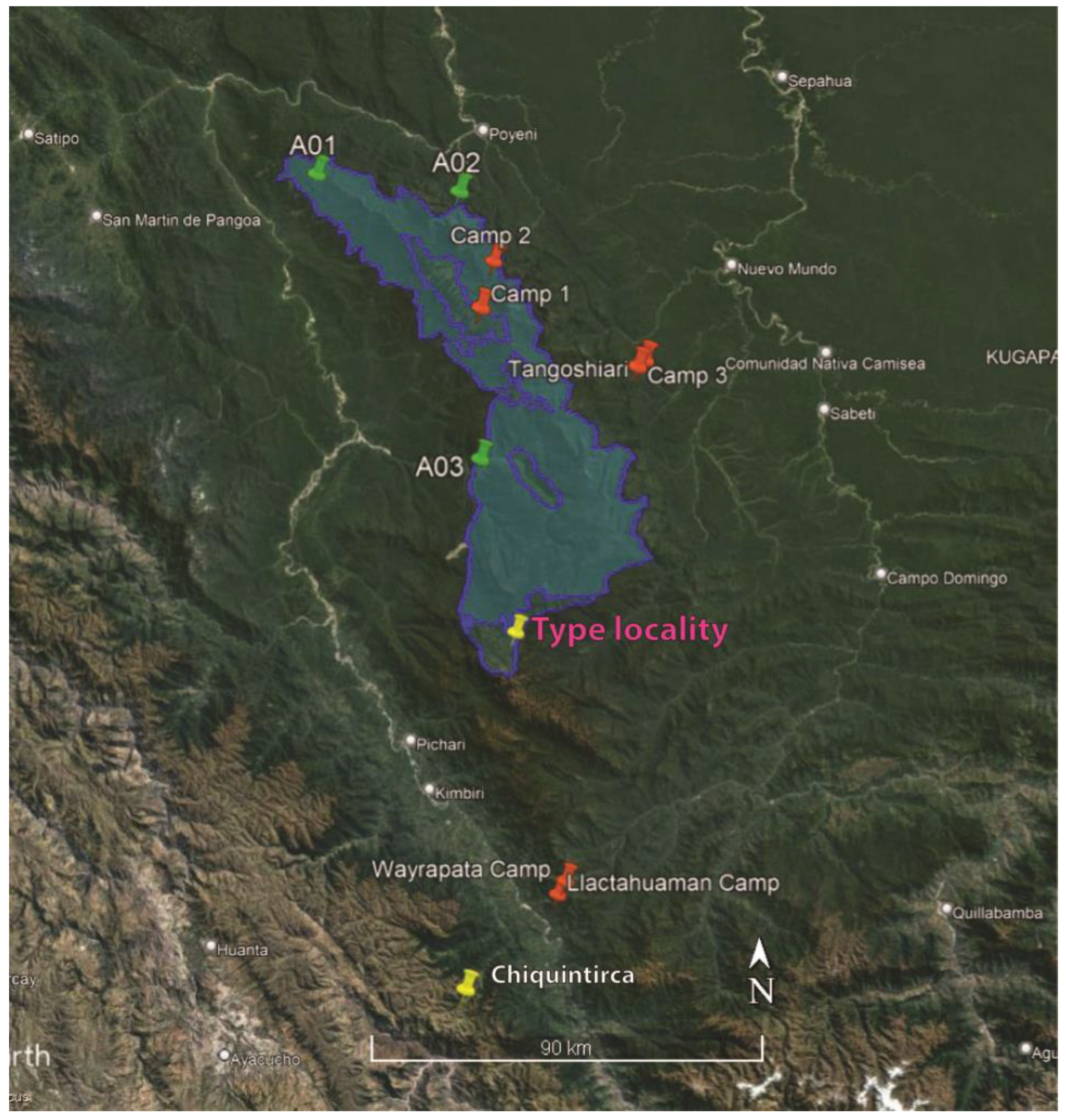
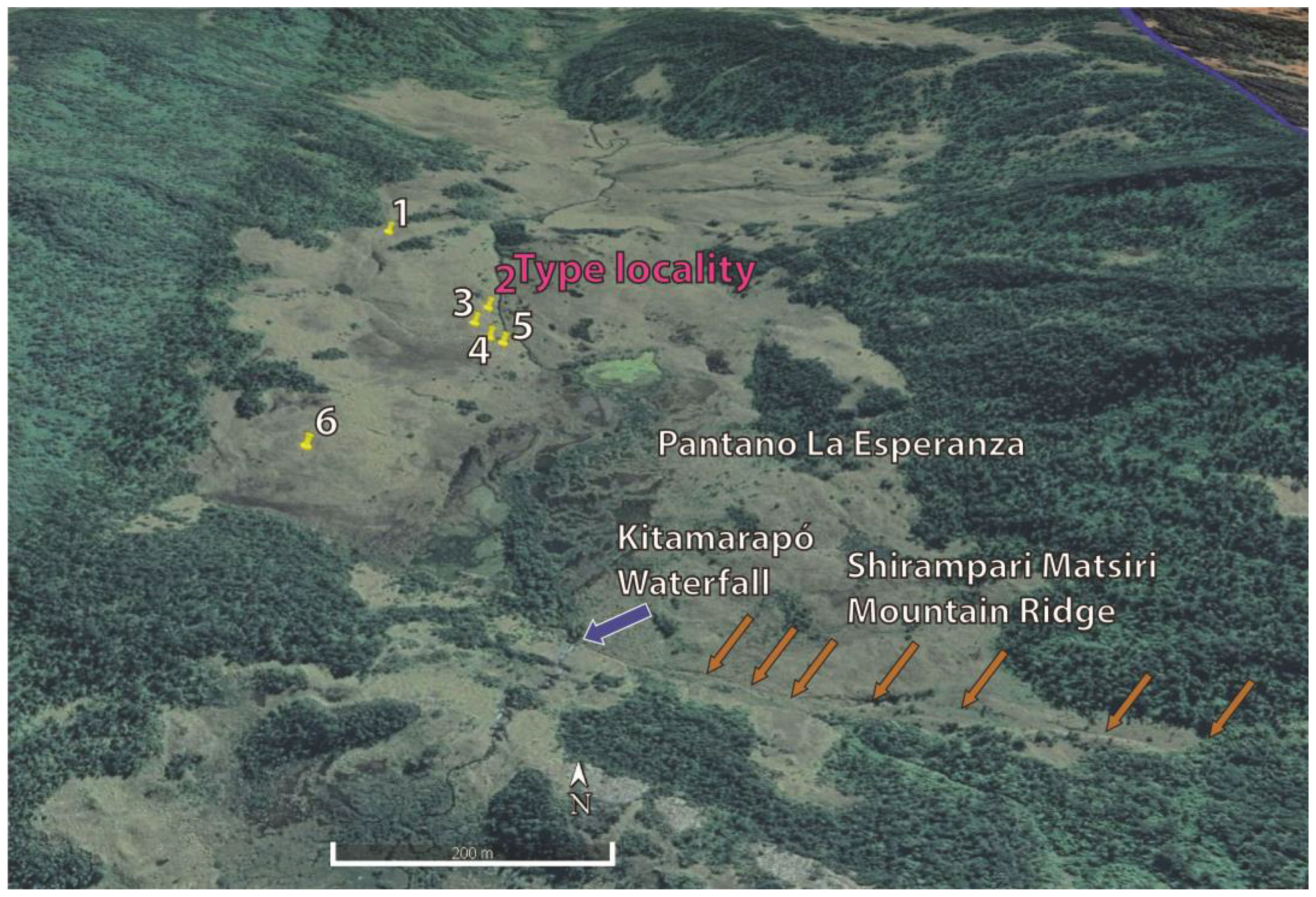
2. Materials and Methods
2.1. Molecular Genetics
2.2. Morphology
3. Results
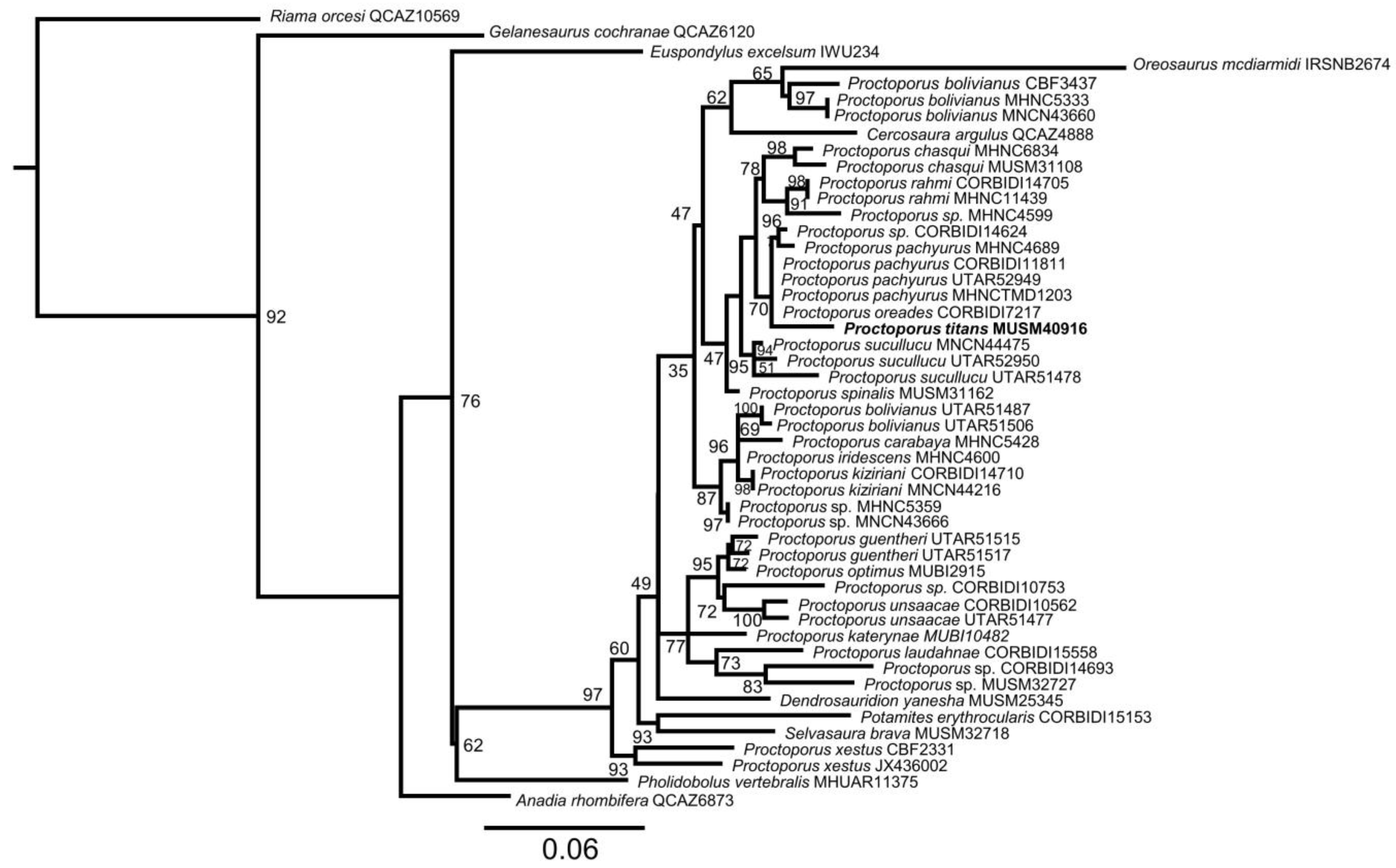
3.1. Molecular Genetics
3.2. Generic Assignment
3.3. Systematics
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Voucher | Accession Code (16S) |
|---|---|---|
| Proctoporus titans sp. nov. | MUSM 40915 | OQ148481 |
| Proctoporus titans sp. nov. | MUSM 40916 | OQ148482 |
| Proctoporus titans sp. nov. | MUSM 40922 | OQ148483 |
| Proctoporus titans sp. nov. | MUSM 40924 | OQ148484 |
Appendix B
Appendix C
| Characters | MUSM 40916 | MUSM 40921 | MUSM 40915 | MUSM 40922 | MUSM 40924 |
|---|---|---|---|---|---|
| Sex | m | m | f | f | f |
| SVL | 53.9 | 41.3 | 46.0 | 44.6 | 48.4 |
| TL | 56.8 r | 35.6 r | 60.8 | 63.8 | 48.8 b, r |
| HL | 10.7/10.9 | 8.9/8.9 | 9.0/9.6 | 8.8/9.6 | 9.2/8.4 |
| HW | 8.2 | 5.4 | 5.1 | 4.9 | 5.7 |
| HD | 5.7 | 4.7 | 3.9 | 3.8 | 4.4 |
| E-N | 3.8/3.6 | 2.8/2.8 | 2.9/2.7 | 3.1/2.9 | 2.9/2.9 |
| FLL | –/10.4 | 8.2/6.9 | 8.0/6.6 | 9.7/– | 9.8/– |
| HLL | 14.1/13.7 | 11.7/12.1 | –/12.9 | 11.3/10.8 | 13.9/10.6 |
| AGD | 28.0/26.7 | 21.7/21.5 | 21.5/22.4 | 23.5/23.2 | 26.9/25.2 |
| Supralabials | 5/5 | 6/6 | 6/6 | 6/6 | 6/6 |
| Infralabials | 5/5 | 6/6 | 5/5 | 5/5 | 5/6 |
| Anterior infralabials | 4/3 | 5/3 | 4/4 | 4/4 | 4/4 |
| Nasal scales | 1/1 inc | 1/1 inc | 1/1 und | 1/1 und | 1/1 inc |
| Pairs of genials in contact | 1 | 1 | 2 | 2 | 1 |
| Gular scale rows | 2 | 2 | 2 | 3 | 2 |
| Collar scale rows | 7 | 6 | 6 | 7 | 7 |
| Dorsal scales | 34 | 33 | 35 | 36 | 36 |
| Longitudinal dorsal scale rows | 21 | 20 | 21 | 20 | 21 |
| Lateral scales | 3/2 | 3/3 | 3/3 | 2/3 | 2/2 |
| Scales around midbody | 35 | 34 | 35 | 34 | 35 |
| Ventral scales | 21 | 21 | 21 | 20 | 23 |
| Longitudinal ventral scale rows | 10 | 10 | 10 | 10 | 10 |
| Anterior preanal plate scales | 2 | 2 | 2 | 2 | 2 |
| Posterior cloacal plate scales | 4 | 4 | 4 | 6 | 5 |
| Lamellae under Finger IV | –/13 | 11/10 | 12/12 | 10/– | 10/– |
| Lamellae under Toe IV | 19/19 | 17/17 | –/17 | 15/15 | 17/17 |
| Femoral pores | 5/5 | 6/6 | – | – | – |
Appendix D
| Characters | MUSM 35638 | MUSM 37754 | MUSM 37755 | MUSM 37757 | MUSM 35637 | MUSM 35639 | MUSM 37756 | MUSM 37758 | MUSM 37789 |
|---|---|---|---|---|---|---|---|---|---|
| Sex | m | m | m | m | f | f | f | f | f |
| SVL | 43.2 | 44.7 | 46.3 | 51.0 | 47.5 | 50.1 | 52.2 | 43.6 | 52.6 |
| TL | 62.2 | 55.2 r | 44.5 r | 65.3 r | 36.5 r, b | 61.4 | 36.6 r | 45.4 r | 52.2 i |
| HL | 11.5/11.7 | 11.0/9.9 | 12.0/12.4 | 12.1/12.7 | 9.8/10.7 | 10.4/11.0 | 10.5/11.3 | 10.9/10.0 | 10.8/11.0 |
| HW | 6.4 | 6.5 | 7.6 | 7.4 | 6.8 | 6.3 | 7.0 | 6.0 | 5.8 |
| HD | 4.7 | 5.4 | 4.9 | 5.3 | 4.4 | 4.9 | 5.0 | 4.5 | 4.7 |
| E-N | 3.3/3.8 | 3.5/3.4 | 3.8/3.5 | 4.0/4.0 | 3.4/3.2 | 3.8/3.7 | 3.2/3.9 | 3.4/3.4 | 3.3/3.8 |
| FLL | 12.4/10.0 | 9.6/11.8 | 11.0/12.7 | 11.3/10.6 | 11.6/10.3 | 10.5/9.9 | 11.2/10.2 | 7.7/8.7 | 11.9/11.6 |
| HLL | 14.0/14.5 | 14.3/16.1 | 17.7/13.4 | 13.6/16.7 | 16.0/15.2 | 14.0/12.9 | 15.6/13.9 | 12.6/15.1 | 16.1/15.1 |
| AGD | 23.6/24.1 | 22.0/24.1 | 23.0/21.6 | 23.3/26.3 | 25.4/21.9 | 25.8/27.9 | 29.5/27.8 | 23.4/22.3 | 28.5/28.4 |
| Supralabials | 7/7 | 7/7 | 5/5 | 7/7 | 6/6 | 6/6 | 6/6 | 6/6 | 6/6 |
| Infralabials | 4/4 | 4/4 | 6/6 | 4/4 | 4/4 | 4/4 | 4/4 | 4/4 | 4/4 |
| Anterior infralabials | 3/3 | 3/3 | 3/3 | 3/3 | 4/3 | 3/3 | 3/3 | 3/3 | 4/4 |
| Nasal scales | 1/1 inc | 1/1 inc | 1/1 inc | 1/1 inc | 1/1 div | 1/1 inc | 1/1 inc | 1/1 div | 1/1 div |
| Pairs of genials in contact | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 3 |
| Gular scale rows | 4 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 2 |
| Collar scale rows | 6 | 7 | 7 | 7 | 7 | 7 | 7 | 6 | 7 |
| Dorsal scales | 38 | 36 | 38 | 37 | 35 | 37 | 39 | 39 | 41 |
| Longitudinal dorsal scale rows | 22 | 22 | 24 | 25 | 22 | 20 | 21 | 22 | 23 |
| Lateral scales | 2/2 | 3/2 | 2/2 | 2/2 | 3/2 | 3/3 | 2/2 | 2/2 | 4/3 |
| Scales around midbody | 38 | 39 | 38 | 40 | 38 | 35 | 37 | 35 | 39 |
| Ventral scales | 19 | 21 | 20 | 20 | 20 | 21 | 20 | 21 | 22 |
| Longitudinal ventral scale rows | 12 | 12 | 10 | 12 | 11 | 10 | 10 | 10 | 10 |
| Anterior preanal plate scales | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| Posterior cloacal plate scales | 4 | 5 | 3 | 2 | 6 | 5 | 6 | 5 | 4 |
| Lamellae under Finger IV | 12/13 | 12/12 | 13/12 | 13/13 | 13/13 | 13/13 | 12/12 | 13/13 | 13/14 |
| Lamellae under Toe IV | 18/18 | 18/18 | 18/19 | 19/18 | 18/19 | 18/19 | 18/18 | 19/19 | 20/19 |
| Femoral pores | 6/6 | 5/6 | 7/7 | 7/7 | – | – | – | – | – |
References
- SERNANP. Guía Oficial De Áreas Naturales Protegidas Del Perú; Ministerio del Ambiente: Lima, Peru, 2010; pp. 1–208.
- Baekeland, G.B. By parachute into Peru’s lost world. Natl. Geogr. 1964, 126, 268–296. [Google Scholar]
- Alonso, L.E.; Alonso, A.; Schulenberg, T.S.; Dallmeier, F. (Eds.) Biological and Social Assessments of the Cordillera de Vilcabamba, Peru. RAP Working Papers 12 and SI/MAB Series 6; Conservation International: Washington, DC, USA, 2001; pp. 1–296. [Google Scholar]
- Lehr, E. New eleutherodactyline frogs (Leptodactylidae: Pristimantis, Phrynopus) from Peru. Bull. Mus. comp. Zool. 2007, 159, 145–178. [Google Scholar] [CrossRef]
- Mendoza, W. Especie nueva de Polylepis (Rosaceae) de la cordillera Vilcabamba (Cusco, Perú). Rev. Peru. Biol. 2005, 12, 103–106. [Google Scholar] [CrossRef][Green Version]
- Mamani, L.; Rodríguez, L. A new species of Andean lizard, Proctoporus (Gymnophthalmidae: Cercosaurinae), from the highland of Parque Nacional Otishi in Peru. Zootaxa 2022, 5213, 75–85. [Google Scholar] [CrossRef]
- INRENA. Parque Nacional Otishi. Plan Maestro 2005–2010; Instituto Nacional de Recursos Naturales—INRENA: Lima, Peru, 2003; pp. 1–118.
- Rodríguez, L.O.; Young, K.R. Biological diversity of Peru: Determining priority areas for conservation. AMBIO A J. Hum. Environ. 2000, 29, 329–337. [Google Scholar] [CrossRef]
- Fajardo, J.; Lessmann, J.; Bonaccorso, E.; Devenish, C.; Muñoz, J. Combined use of systematic conservation planning, species distribution modeling, and connectivity analysis reveals severe conservation gaps in a megadiverse country (Peru). PLoS ONE 2014, 9, e114367. [Google Scholar] [CrossRef]
- Barboza Sánchez, C.A.N. Composición y Distribución Altitudinal de Los Anfibios del Parque Nacional Otishi, Cordillera Central del Perú. Tesis Para Obtar el Titulo de Biologo; Universidad Nacional de Trujillo: Trujillo, Peru, 2014; pp. 1–75. [Google Scholar]
- von Tschudi, J.J. Reptilium conspectum quae in republica Peruana reperiuntur et pleraque observata vel collecta sunt in itenere. Arch. Für Nat. 1845, 11, 150–170. [Google Scholar]
- Mamani, L.; Cruz, R.; Mallqui, S.; Catenazzi, A. Molecular phylogenetics and comparative examination of voucher museums reveal two new species of gymnophthalmid lizards (Squamata, Gymnophthalmidae) from the Peruvian Andes, with comments on Proctoporus guentheri (Boettger, 1891). Diversity 2022, 14, 215. [Google Scholar] [CrossRef]
- Uetz, P.; Freed, P.; Aguilar, R.; Hošek, J. (Eds.) The Reptile Database. Available online: http://www.reptile-database.org (accessed on 11 December 2022).
- Werner, F. Über neue oder seltene Reptilien des Naturhistorischen Museums in Hamburg. II. Eidechsen. Jahrb. Hamburg. Wiss. Anst. 1910, 27 (Suppl. S2), 1–46. [Google Scholar]
- Goicoechea, N.; Padial, J.M.; Chaparro, J.C.; Castroviejo-Fisher, S.; De la Riva, I. A taxonomic revision of Proctoporus bolivianus Werner (Squamata: Gymnophthalmidae) with the description of three new species and resurrection of Proctoporus lacertus Stejneger. Am. Mus. Nov. 2013, 3786, 1–32. [Google Scholar] [CrossRef][Green Version]
- Chávez, G.; Siu-Ting, K.; Duran, V.; Venegas, P.J. Two new species of Andean gymnophthalmid lizards of the genus Euspondylus (Reptilia, Squamata) from central and southern Peru. ZooKeys 2011, 109, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Boettger, O. Reptilien und Batrachier aus Bolivien. Zool. Anz. 1891, 14, 343–347. [Google Scholar]
- Stejneger, L. Results of the Yale Peruvian Expedition of 1911. Batrachians and reptiles. Proc. United States Natl. Mus. 1913, 45, 541–547. [Google Scholar] [CrossRef][Green Version]
- Köhler, G.; Lehr, E. Comments on Euspondylus and Proctoporus (Squamata: Gymnophthalmidae) from Peru, with the description of three new species and a key to the Peruvian species. Herpetologica 2004, 60, 501–518. [Google Scholar] [CrossRef]
- Mamani, L.; Goicoechea, N.; Chaparro, J.C. A new species of Andean lizard Proctoporus (Squamata: Gymnophthalmidae) from montane forest of the Historic Sanctuary of Machu Picchu, Peru. Amphib. Reptile Conserv. 2015, 9, e96. [Google Scholar]
- De Grijs, P. Prionodactylus rahmi, eine neue Eidechse aus den Anden. Zool. Anz. 1936, 116, 27–30. [Google Scholar]
- Boulenger, G.A. Descriptions of new reptiles from the Andes of South America, preserved in the British Museum. Ann. Mag. Nat. Hist. 1911, 7, 19–25. [Google Scholar] [CrossRef][Green Version]
- Doan, T.M.; Castoe, T.A. Using morphological and molecular evidence to infer species boundaries within Proctoporus bolivianus Werner (Squamata: Gymnopthalmidae). Herpetologica 2003, 59, 432–449. [Google Scholar] [CrossRef]
- Uzzell, T. A new genus and species of teiid lizard from Bolivia. Postilla 1969, 129, 1–15. [Google Scholar] [CrossRef]
- Goicoechea, N.; Padial, J.M.; Chaparro, J.C.; Castroviejo-Fisher, S.; De la Riva, I. Data from: Molecular phylogenetics, species diversity, and biogeography of the Andean lizards Proctoporus (Squamata: Gymnophthalmidae). Dryad Dataset 2013. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Oftedal, O.T. A revision of the genus Anadia (Sauria, Teiidae). Arq. Zool. 1974, 25, 203–265. [Google Scholar] [CrossRef]
- Chávez, G.; Catenazzi, A.; Venegas, P.J. A new species of arboreal microteiid lizard of the genus Euspondylus (Gymnophthalmidae: Cercosaurinae) from the Andean slopes of central Peru with comments on Peruvian Euspondylus. Zootaxa 2017, 4350, 301–316. [Google Scholar] [CrossRef]
- Sánchez-Pacheco, S.J.; Nunes, P.M.S.; Marques-Souza, S.; Rodrigues, M.T.; Murphy, R.W. Formal recognition of the species of Oreosaurus (Reptilia, Squamata, Gymnophthalmidae) from the Sierra Nevada de Santa Marta, Colombia. ZooKeys 2017, 691, 149–162. [Google Scholar] [CrossRef]
- Sánchez-Pacheco, S.J.; Torres-Carvajal, O.; Aguirre-Peñafiel, V.; Nunes, P.M.; Verrastro, L.; Rivas, G.A.; Rodrigues, M.T.; Grant, T.; Murphy, R.W. Phylogeny of Riama (Squamata: Gymnophthalmidae), impact of phenotypic evidence on molecular datasets, and the origin of the Sierra Nevada de Santa Marta endemic fauna. Cladistics 2017, 34, 260–291. [Google Scholar] [CrossRef]
- Moravec, J.; Šmíd, J.; Štundl, J.; Lehr, E. Systematics of Neotropical microteiid lizards (Gymnophthalmidae, Cercosaurinae), with the description of a new genus and species from the Andean montane forests. ZooKeys 2018, 774, 105–139. [Google Scholar] [CrossRef]
- IUCN. IUCN Standards and Petitions Subcommittee. Guidelines for Using the IUCN Red List Categories and Criteria. Version 10.1. Prepared by the Standards and Petitions Subcommittee. 2013. Available online: http://www.iucnredlist.org/documents/RedListGuidelines.pdf (accessed on 15 September 2022).
- Castoe, T.A.; Doan, T.M.; Parkinson, C.L. Data partitions and complex models in Bayesian analyses: The phylogeny of gymnophthalmid lizards. Syst. Biol. 2004, 53, 448–469. [Google Scholar] [CrossRef]
- Fitzinger, L. Neue Classification der Reptilien Nach Ihren Natürlichen Verwandtschaften Nebst Einer Verwandschafts-Tafel Und Einem Verzeichnisse der Reptilien-Sammlung des K. K. Zoologischen Museums zu Wien; Im Verlage von J.G. Heubner: Wien, Austria, 1826; pp. 1–66. [Google Scholar]
- Gray, J.E. Catalogue of the slender-tongued saurians, with descriptions of many new genera and species. Part 1. Ann. Mag. Nat. Hist. London 1838, 1, 274–283. [Google Scholar] [CrossRef]
- Chávez, G.; Chávez-Arribasplata, J.C. Distribution and natural history notes on the Peruvian lizard Proctoporus laudahnae (Squamata: Gymnophthalmidae). Phyllomedusa 2016, 15, 147–154. [Google Scholar] [CrossRef]
- Holdridge, L.R. Life Zone Ecology; Tropical Science Center: San Jose, Costa Rica, 1967; pp. 1–206. [Google Scholar]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lehr, E.; Cusi, J.C.; Fernandez, M.I.; Vera, R.J.; Catenazzi, A. A New Species of Proctoporus (Reptilia, Gymnophthalmidae, Cercosaurinae) from the Puna of the Otishi National Park in Peru. Taxonomy 2023, 3, 10-28. https://doi.org/10.3390/taxonomy3010002
Lehr E, Cusi JC, Fernandez MI, Vera RJ, Catenazzi A. A New Species of Proctoporus (Reptilia, Gymnophthalmidae, Cercosaurinae) from the Puna of the Otishi National Park in Peru. Taxonomy. 2023; 3(1):10-28. https://doi.org/10.3390/taxonomy3010002
Chicago/Turabian StyleLehr, Edgar, Juan C. Cusi, Maura I. Fernandez, Ricardo J. Vera, and Alessandro Catenazzi. 2023. "A New Species of Proctoporus (Reptilia, Gymnophthalmidae, Cercosaurinae) from the Puna of the Otishi National Park in Peru" Taxonomy 3, no. 1: 10-28. https://doi.org/10.3390/taxonomy3010002
APA StyleLehr, E., Cusi, J. C., Fernandez, M. I., Vera, R. J., & Catenazzi, A. (2023). A New Species of Proctoporus (Reptilia, Gymnophthalmidae, Cercosaurinae) from the Puna of the Otishi National Park in Peru. Taxonomy, 3(1), 10-28. https://doi.org/10.3390/taxonomy3010002