
Molecular Characterization of Spirometra decipiens Complex (Eucestoda: Diphyllobothriidea) from Uruguay

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Ethics Concerning Wild Animal Samples
2.2. Sampling
2.3. DNA Extraction, PCR and Sequencing
2.4. Phylogenetic Analyses
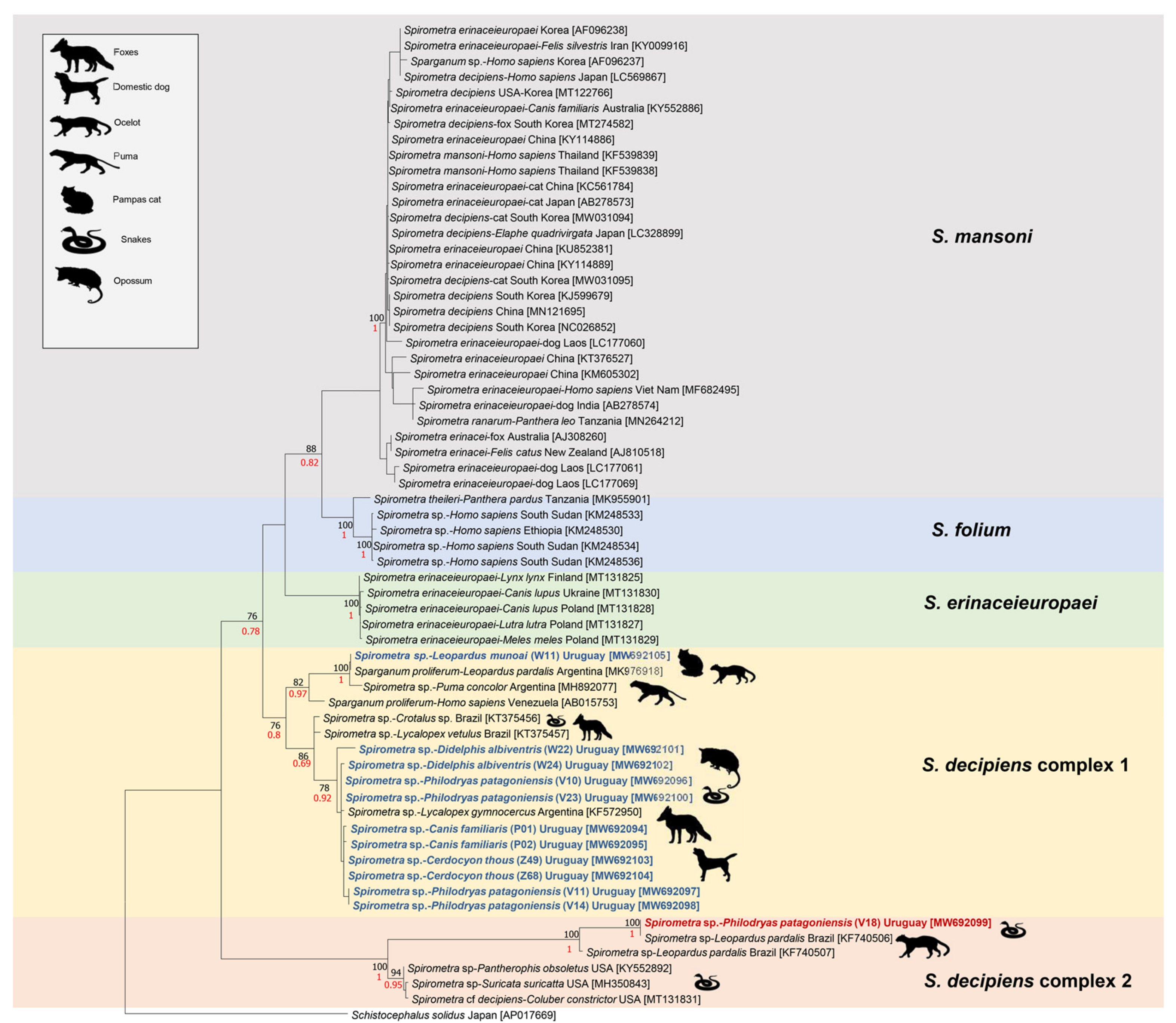
3. Results
3.1. Samples
3.2. PCR and Sequencing
3.3. Phylogenetic Analyses
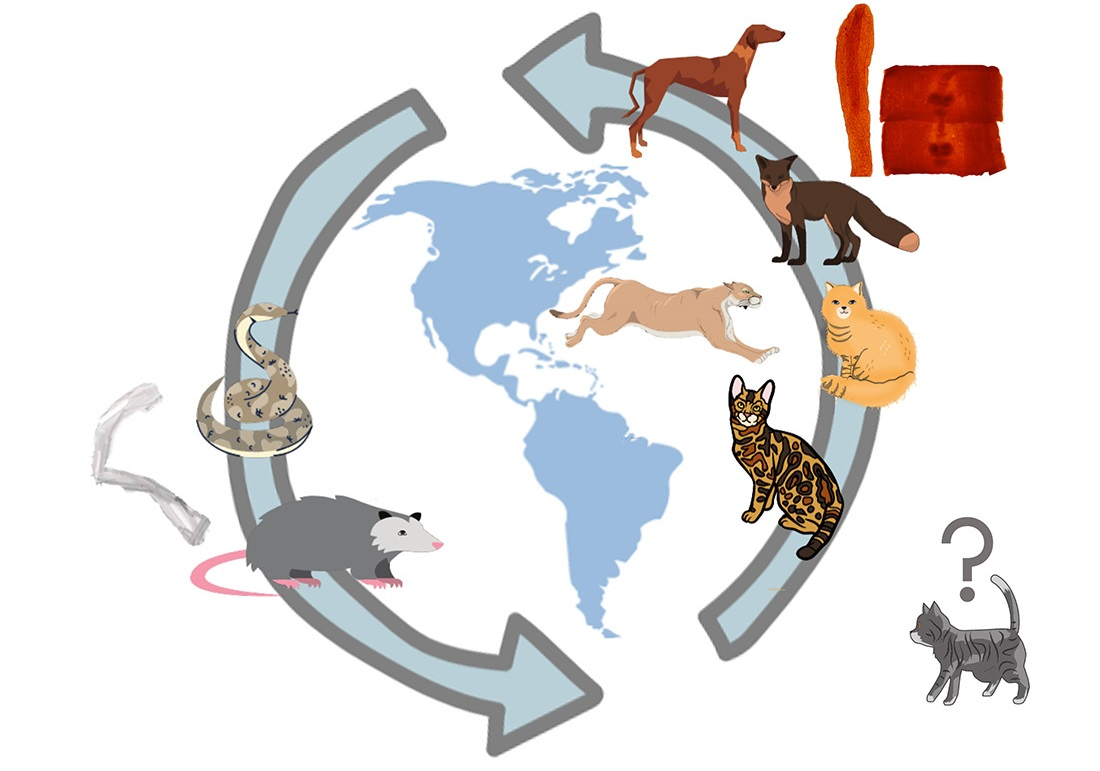
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Waeschenbach, A.; Brabec, J.; Scholz, T.; Littlewood, D.T.J.; Kuchta, R. The catholic taste of broad tapeworms–multiple routes to human infection. Int. J. Parasitol. 2017, 47, 831–843. [Google Scholar] [CrossRef]
- Kuchta, R.; Scholz, T.; Brabec, J.; Narduzzi-Wicht, B. Diphyllobothrium, Diplogonoporus and Spirometra. In Biology of Foodborne Parasites. Section III Important Foodborne Parasites; Xiao, L., Ryan, U., Feng, Y., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 299–326. [Google Scholar]
- Scholz, T.; Kuchta, R.; Brabec, J. Broad tapeworms (Diphyllobothriidae), parasites of wildlife and humans: Recent progress and future challenges. Int. J. Parasitol. Parasites Wildl. 2019, 9, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, T.; Gutierrez, C.; Rodriguez, A.; Sauto, S. Testicular sparganosis in a child from Uruguay. Acta Trop. 2003, 88, 83–86. [Google Scholar] [CrossRef]
- Kuchta, R.; Kołodziej-Sobocińska, M.; Brabec, J.; Młocicki, D.; Sałamatin, R.; Scholz, T. Sparganosis (Spirometra). In Europe in the molecular era. Clin. Infect. Dis. 2021, 72, 882–890. [Google Scholar] [CrossRef] [PubMed]
- Oda, F.H.; Borteiro, C.; da Graça, R.J.; Tavares, L.E.R.; Crampet, A.; Guerra, V.; Lima, F.S.; Bellay, S.; Karling, L.C.; Castro, O.; et al. Parasitism by larval tapeworms genus Spirometra in South American amphibians and reptiles: New records from Brazil and Uruguay, and a review of current knowledge in the region. Acta Trop. 2016, 164, 150–164. [Google Scholar] [CrossRef] [PubMed]
- Miyadera, H.; Kokaze, A.; Kuramochi, T.; Kita, K.; Machinami, R.; Noya, O.; de Noya, B.A.; Okamoto, M.; Kojima, S. Phylogenetic identification of Sparganum proliferum as a pseudophyllidean cestode by the sequence analyses on mitochondrial COI and nuclear sdhB genes. Parasitol. Int. 2001, 50, 93–104. [Google Scholar] [CrossRef]
- Almeida, G.G.; Coscarelli, D.; Melo, M.N.; Melo, A.L.; Pinto, H.A. Molecular identification of Spirometra spp. (Cestoda: Diphyllobothriidae) in some wild animals from Brazil. Parasitol. Int. 2016, 65, 428–431. [Google Scholar] [CrossRef]
- McHale, B.; Callahan, R.T.; Paras, K.L.; Weber, M.; Kimbrell, L.; Velázquez-Jiménez, Y.; McManamon, R.; Howerth, E.W.; Verocai, G.G. Sparganosis due to Spirometra sp. (cestoda; Diphyllobothriidae) in captive meerkats (Suricata suricatta). Int. J. Parasitol. Parasites Wildl. 2020, 13, 186–190. [Google Scholar] [CrossRef]
- Yamasaki, H.; Sanpool, O.; Rodpai, R.; Sadaow, L.; Laummaunwai, P.; Un, M.; Thanchomnang, T.; Laymanivong, S.; Pa Pa Aung, W.; Intapan, P.M.; et al. Spirometra species from Asia: Genetic diversity and taxonomic challenges. Parasitol. Int. 2021, 80, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Holcman-Spector, B.; Olagüe, G.; Couto, A. Helmintiasis del perro vagabundo (Canis familiaris) en la ciudad de Montevideo. Revista Uruguaya Patología Clínica 1985, 21, 67–73. [Google Scholar]
- Sampaio, I.; Castro, E.; Chifflet, L.; Areosa, O. Hallazgo de Diphyllobothrium sp. en Canis familiaris. Veterinaria (Montev.) 1992, 28, 22–23. [Google Scholar]
- Osimani, J.J.; Dei-Cas, E.V. Sobre la obtención experimental de Spirometra sp. (Cestoda-Dibothriocephalidae). Neotropica 1974, 20, 57–63. [Google Scholar]
- Castro, O.; Venzal, J.M.; Felix, M.L. Two new records of helminth parasites of domestic cat from Uruguay: Alaria alata (Goeze, 1782) (Digenea, Diplostomidae) and Lagochilascaris major Leiper, 1910 (Nematoda, Ascarididae). Vet. Parasitol. 2009, 160, 344–347. [Google Scholar] [CrossRef]
- Castro, O.; Venzal, J.M.; Morgades, D.; Katz, H.; Gagliardi, F.; Benítez, G. Algunos helmintos nuevos para Uruguay registrados en la Colección de Helmintos Parásitos de Fauna Silvestre del Departamento de Parasitología, Facultad de Veterinaria, UdelaR. In Proceedings of the V Jornadas Técnicas de la Facultad de Veterinaria, Universidad de la República, Montevideo, Uruguay, 21–23 November 2007; V063 JORr. p. 37. [Google Scholar]
- Vogelsang, E.G. La presencia de un sparganum en las ranas (Leptodactylus ocellatus) del Uruguay. Rev. Med. Vet. 1925, 2, 301. [Google Scholar]
- Wolffhügel, K.; Vogelsang, E.G. Dibothriocephalus decipiens (Diesing) y su larva Sparganum reptans en el Uruguay. Rev. Med. Vet. 1926, 2, 433–434. [Google Scholar]
- Dei-Cas, E.; Rodríguez, N.; Botto, C.; Osimani, J.J. Larvas plerocercoides de Spirometra (Dibothriocephalidae) en el hombre y en animales silvestres de Uruguay. Rev. Inst. Med. Trop. Sao Paulo 1976, 18, 165–172. [Google Scholar]
- Borteiro, C.; Castro, O.; Sabalsagaray, M.J.; Kolenc, F.; Martínez Debat, C.; Ubilla, M. Spargana in the Neotropical frog Hypsiboas pulchellus (Hylidae) from Uruguay. North -West J. Zool. 2015, 11, 171–173. [Google Scholar]
- Osimani, J.J.; Peyrallo, R. Segundo caso de esparganosis encontrado en América del Sur. Primer caso descrito en el Uruguay. Arch. Urug. Med. Cir. Espec. 1954, 44, 139–148. [Google Scholar] [PubMed]
- Bowles, J.; Blair, D.; McManus, D.P. Genetic variants within the genus Echinococcus identified by mitochondrial DNA sequencing. Mol. Biochem. Parasitol. 1992, 54, 165–173. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrigh, R.S.; Scioscia, N.P.; Denegri, G.M.; Fugassa, M.H. Cox-1 gene sequence of Spirometra in Pampas foxes from Argentina. Helminthologia 2015, 52, 355–359. [Google Scholar] [CrossRef]
- Ruas, J.L.; Muller, G.; Farias, N.A.R.; Gallina, T.; Lucas, A.S.; Pappen, F.G.; Sinkoc, A.L.; Brum, J.G.W. Helminths of Pampas fox Pseudalopex gymnocercus (Fischer, 1814) and of Crab-eating fox Cerdocyon thous (Linnaeus, 1766) in the Southern of the State of Rio Grande do Sul, Brazil. Rev. Bras. Parasitol. Vet. 2008, 17, 87–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, E.M.; Martínez-Lanfranco, J.A. Mamíferos de Uruguay. Guía de Campo e Introducción a su Estudio y Conservación; Vida Silvestre, Museo Nacional de Historia Natural, Ediciones de la Banda Oriental: Montevideo, Uruguay, 2010; p. 463. [Google Scholar]
- Arrabal, J.P.; Pérez, M.G.; Arce, L.F.; Kamenetzky, L. First identification and molecular phylogeny of Sparganum proliferum from endangered felid (Panthera onca) and other wild definitive hosts in one of the regions with highest worldwide biodiversity. Int. J. Parasitol.: Parasites Wildl. 2020, 13, 142–149. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Sample Code | Host Species | Date | Site—Department | Latitude, Longitude | Diphyllobothriidean Recovered | PCR | Accession Number | Sequence Length (bp) |
|---|---|---|---|---|---|---|---|---|
| P01 | Canis familiaris | 6 June 2017 | Montevideo | * | +a | + | MW692094 | 381 |
| P02 | Canis familiaris | 30 October 2016 | Campo experimental 1, Migues—Canelones | 34°22′22″ S, 55°36′10″ W | +a | + | MW692095 | 372 |
| Z49 | Cerdocyon thous | 8 August 2016 | Route 1, km 67—San José | 34°33′21″ S, 56°44′45″ W | +a | + | MW692103 | 373 |
| Z59 | Cerdocyon thous | 24 October 2016 | Route 3, km 460—Paysandú | 31°35′54″ S, 57°52′20″ W | - | - | - | |
| Z64 | Cerdocyon thous | 1 April 2017 | Route 3, km 427—Paysandú | 31°52′48″ S, 57°51′43″ W | - | - | - | |
| Z68 | Cerdocyon thous | 11 May 2017 | Route 31, km 34—Salto | 31°23′46″ S, 57°35′20″ W | +a | - | - | |
| Z69 | Cerdocyon thous | 11 May 2017 | Route 31, km 30—Salto | 31°23′36″ S, 57°37′48″ W | +a | + | MW692104 | 368 |
| Z71 | Cerdocyon thous | 1 July 2017 | Route 5, km 427—Rivera | 31°31′03″ S, 55°40′54″ W | +a | - | - | |
| Z75 | Cerdocyon thous | 4 July 2017 | Route 2, km 269—Soriano | 33°19′53″ S, 57°58′40″ W | - | - | - | |
| W06 | Didelphis albiventris | 29 March 2016 | Route 31, km 115—Salto | 31°22′49″ S, 56°50′12″ W | - | - | - | |
| W09 | Didelphis albiventris | 30 March 2016 | Tambores—Paysandú | 31°51′34″ S, 56°15′02″ W | - | - | - | |
| W22 | Didelphis albiventris | 18 September 2016 | Route 2, km 287—Río Negro | 33°12′01″ S, 58°04′15″ W | +p | + | MW692101 | 370 |
| W24 | Didelphis albiventris | 26 October 2016 | Route 27, km 61—Rivera | 31°23′56″ S, 55°14′45″ W | +p | + | MW692102 | 338 |
| W36 | Leopardus geoffroyi | 20 September 2017 | Route 31, km 187—Tacuarembó | 31°38′35″ S, 56°17′47″ W | - | - | - | |
| W11 | Leopardus munoai | 12 May 2016 | Route 3, km 210.5—Flores | 33°23′58″ S, 57°01′50″ W | +a | + | MW692105 | 413 |
| Z73 | Lycalopex gymnocercus | 3 July 2017 | Route 11, km 123—Canelones | 34°51′32″ S, 56°03′17″ W | - | - | - | |
| V10 | Philodryas patagoniensis | 11 July 2015 | El Relincho farm—Paysandú | 32°04′41″ S, 57°18′13″ W | +p | + | MW692096 | 374 |
| V11 | Philodryas patagoniensis | 11 July 2015 | El Relincho farm—Paysandú | 32°04′41″ S, 57°18′13″ W | +p | + | MW692097 | 376 |
| V14 | Philodryas patagoniensis | 2 October 2016 | Route 3, km 536.9—Salto | 31°00′87″ S, 57°42′12″ W | +p | + | MW692098 | 370 |
| V18 | Philodryas patagoniensis | 17 October 2016 | Barrio Artigas—Salto | 31°22′59″ S, 57°53′15″ W | +p | + | MW692099 | 352 |
| V23 | Philodryas patagoniensis | 11 February 2016 | Route 3, km 512—Salto | 31°13′27″ S, 57°45′37″ W | +p | + | MW692100 | 354 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Armúa-Fernández, M.T.; Burutarán, M.; Bazzano, V.; Félix, M.L.; Castro, O.; Venzal, J.M. Molecular Characterization of Spirometra decipiens Complex (Eucestoda: Diphyllobothriidea) from Uruguay. Taxonomy 2021, 1, 270-277. https://doi.org/10.3390/taxonomy1030021
Armúa-Fernández MT, Burutarán M, Bazzano V, Félix ML, Castro O, Venzal JM. Molecular Characterization of Spirometra decipiens Complex (Eucestoda: Diphyllobothriidea) from Uruguay. Taxonomy. 2021; 1(3):270-277. https://doi.org/10.3390/taxonomy1030021
Chicago/Turabian StyleArmúa-Fernández, María Teresa, Mauricio Burutarán, Valentin Bazzano, María Laura Félix, Oscar Castro, and José Manuel Venzal. 2021. "Molecular Characterization of Spirometra decipiens Complex (Eucestoda: Diphyllobothriidea) from Uruguay" Taxonomy 1, no. 3: 270-277. https://doi.org/10.3390/taxonomy1030021
APA StyleArmúa-Fernández, M. T., Burutarán, M., Bazzano, V., Félix, M. L., Castro, O., & Venzal, J. M. (2021). Molecular Characterization of Spirometra decipiens Complex (Eucestoda: Diphyllobothriidea) from Uruguay. Taxonomy, 1(3), 270-277. https://doi.org/10.3390/taxonomy1030021