
Susceptibility of Tetranychus urticae to the Alkaloidal Extract of Zanthoxylum schreberi Bark: Phenotypic and Biochemical Insights for Biotechnological Exploitation



Abstract
1. Introduction
2. Materials and Methods
2.1. Mite Rearing
2.2. Preparation of Alkaloidal Extract from Z. schreberi Bark
2.3. Susceptibility of T. urticae to the Alkaloidal Extract of Z. schreberi
2.3.1. Preliminary Mortality Test with Unexposed Mites
2.3.2. Mortality and Repellency Tests for Z. schreberi Bark Extract Using Exposed Mites
2.3.3. Effects on Mortality, Repellency, and Oviposition of Z. schreberi Bark Extract Combined with Commercial Acaricides
2.4. LC-MS Characterization of Z. schreberi Bark Extract
2.5. Molecular Docking Studies
2.6. Purification of Selected Alkaloids from Z. schreberi Bark Extract by Semipreparative HPLC
2.7. Enzyme Activity and Inhibition
2.7.1. Preparation of the Protein Homogenate from T. uricae Adult Females
2.7.2. Acetylcholinesterase (AChE)
2.7.3. Glutathione S-Transferase (GST)
2.7.4. Esterase
2.7.5. Cytochrome P450 Monoxygenase (PMO)
2.7.6. Enzyme Inhibition
2.8. Experimental Design and Statistical Analysis
3. Results
3.1. Preliminary Test Using Unexposed Mite Population
3.2. Mortality and Repellency Test for Exposed Mite Population
3.3. Mortality, Repellency, and Oviposition Test for Extract Combined with Acaricides Using the Exposed Mite Population
3.4. Chemical Characterization of Z. schreberi Bark Extract
3.5. Enzyme Activities of Z. schreberi-Treated and Untreated Mites (Exposed Population)
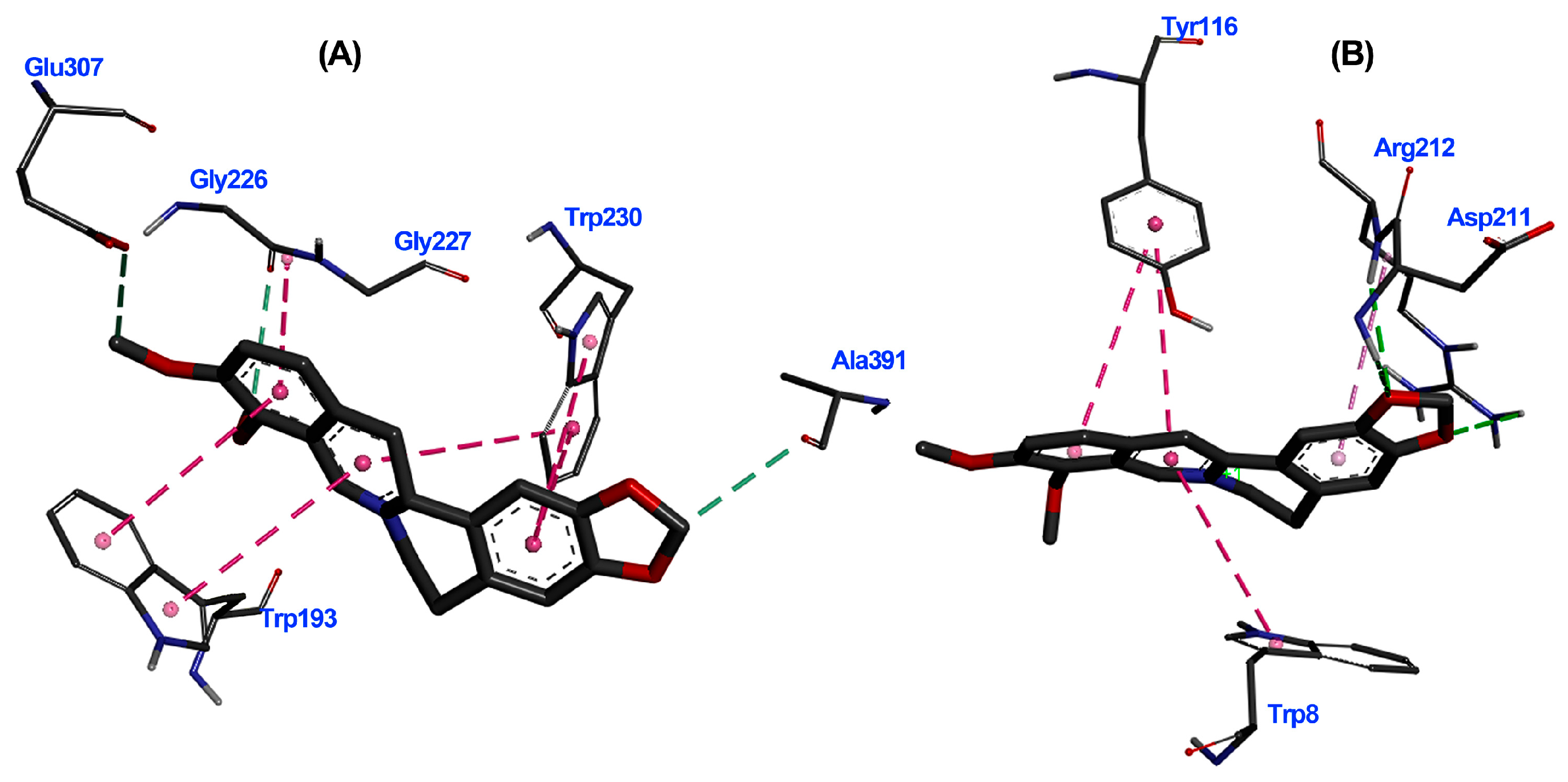
3.6. Molecular Docking Studies
Binding Mode of Top-Ranked Compounds within Active Sites of TuAChE and TuGST
3.7. Enzyme Inhibition of Z. schreberi Extract and Isolated Alkaloids
4. Discussion
5. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Attia, S.; Grissa, K.L.; Lognay, G.; Bitume, E.; Hance, T.; Mailleux, A.C. A Review of the Major Biological Approaches to Control the Worldwide Pest Tetranychus urticae (Acari: Tetranychidae) with Special Reference to Natural Pesticides. J. Pest Sci. 2013, 86, 361–386. [Google Scholar] [CrossRef]
- Rincón, R.A.; Rodríguez, D.; Coy-Barrera, E. Botanicals Against Tetranychus urticae Koch Under Laboratory Conditions: A Survey of Alternatives for Controlling Pest Mites. Plants 2019, 8, 272. [Google Scholar] [CrossRef] [PubMed]
- Isman, M.B. Botanical Insecticides: A Global Perspective. In Biopesticides: State of the Art and Future Opportunities; Gross, A.D., Coats, J.R., Duke, S.O., Seiber, J.N., Eds.; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 2014; Volume 1172, pp. 21–30. ISBN 978-0-8412-2999-0. [Google Scholar]
- Isman, M.B. Botanical Insecticides, Deterrents, and Repellents in Modern Agriculture and an Increasingly Regulated World. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef] [PubMed]
- Appelhans, M.S.; Reichelt, N.; Groppo, M.; Paetzold, C.; Wen, J. Phylogeny and Biogeography of the Pantropical Genus Zanthoxylum and Its Closest Relatives in the Proto-Rutaceae Group (Rutaceae). Mol. Phylogenet. Evol. 2018, 126, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Mutinda, E.S.; Kimutai, F.; Mkala, E.M.; Waswa, E.N.; Odago, W.O.; Nanjala, C.; Ndungu, C.N.; Gichua, M.K.; Njire, M.M.; Gituru, R.W.; et al. Ethnobotanical Uses, Phytochemistry and Pharmacology of Pantropical Genus Zanthoxylum L. (Rutaceae): An Update. J. Ethnopharmacol. 2023, 303, 115895. [Google Scholar] [CrossRef] [PubMed]
- Ombito, J.O. Phytochemistry and Pharmacology of the Genus Zanthoxylum (Rutaceae): A Review. Nat. Prod. J. 2021, 11, 21–43. [Google Scholar] [CrossRef]
- Verma, K.; Kumar, B.; Raj, H.; Sharma, A. A Review on Chemical Constituents, Traditional Uses, Pharmacological Studies of Zanthoxylum armatum (Rutaceae). J. Drug Deliv. Ther. 2021, 11, 136–142. [Google Scholar] [CrossRef]
- Tewary, D.K.; Bhardwaj, A.; Shanker, A. Pesticidal Activities in Five Medicinal Plants Collected from Mid Hills of Western Himalayas. Ind. Crops Prod. 2005, 22, 241–247. [Google Scholar] [CrossRef]
- Wang, C.-F.; Zhang, W.-J.; You, C.-X.; Guo, S.-S.; Geng, Z.-F.; Fan, L.; Du, S.-S.; Deng, Z.-W.; Wang, Y.-Y. Insecticidal Constituents of Essential Oil Derived from Zanthoxylum armatum against Two Stored-Product Insects. J. Oleo Sci. 2015, 64, 861–868. [Google Scholar] [CrossRef]
- Wang, C.F.; Yang, K.; Zhang, H.M.; Cao, J.; Fang, R.; Liu, Z.L.; Du, S.S.; Wang, Y.Y.; Deng, Z.W.; Zhou, L. Components and Insecticidal Activity against the Maize Weevils of Zanthoxylum schinifolium Fruits and Leaves. Molecules 2011, 16, 3077–3088. [Google Scholar] [CrossRef]
- Overgaard, H.J.; Sirisopa, P.; Mikolo, B.; Malterud, K.E.; Wangensteen, H.; Zou, Y.-F.; Paulsen, B.S.; Massamba, D.; Duchon, S.; Corbel, V.; et al. Insecticidal Activities of Bark, Leaf and Seed Extracts of Zanthoxylum heitzii against the African Malaria Vector Anopheles gambiae. Molecules 2014, 19, 21276–21290. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.-S.; Li, Y.-Q. Studies on Acaricidal Activities and Mechanism of Zanthoxylum bungeanum Maxim. Against Tetranychus viennensis Zacher. J. Shanxi Agric. Sci. 2008, 36, 38–41. [Google Scholar]
- Tennyson, S.; Ravindran, K.J.; Arivoli, S. Screening of Twenty Five Plant Extracts for Larvicidal Activity against Culex quinquefasciatus Say (Diptera: Culicidae). Asian Pac. J. Trop. Biomed. 2012, 2, S1130–S1134. [Google Scholar] [CrossRef]
- Van Leeuwen, T.; Vontas, J.; Tsagkarakou, A.; Dermauw, W.; Tirry, L. Acaricide Resistance Mechanisms in the Two-Spotted Spider Mite Tetranychus urticae and Other Important Acari: A Review. Insect Biochem. Mol. Biol. 2010, 40, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Santamaria, M.E.; Arnaiz, A.; Rosa-Diaz, I.; González-Melendi, P.; Romero-Hernandez, G.; Ojeda-Martinez, D.A.; Garcia, A.; Contreras, E.; Martinez, M.; Diaz, I. Plant Defenses Against Tetranychus urticae: Mind the Gaps. Plants 2020, 9, 464. [Google Scholar] [CrossRef]
- Adesanya, A.W.; Lavine, M.D.; Moural, T.W.; Lavine, L.C.; Zhu, F.; Walsh, D.B. Mechanisms and Management of Acaricide Resistance for Tetranychus urticae in Agroecosystems. J. Pest Sci. 2021, 94, 639–663. [Google Scholar] [CrossRef]
- Dermauw, W.; Wybouw, N.; Rombauts, S.; Menten, B.; Vontas, J.; Grbić, M.; Clark, R.M.; Feyereisen, R.; Van Leeuwen, T. A Link between Host Plant Adaptation and Pesticide Resistance in the Polyphagous Spider Mite Tetranychus urticae. Proc. Natl. Acad. Sci. USA 2013, 110, E113–E122. [Google Scholar] [CrossRef]
- Shivanandappa, T.; Rajashekar, Y. Mode of Action of Plant-Derived Natural Insecticides. In Advances in Plant Biopesticides; Singh, D., Ed.; Springer: New Delhi, India, 2014; pp. 323–345. ISBN 978-81-322-2006-0. [Google Scholar]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World Map of the Köppen-Geiger Climate Classification Updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- Sparks, T.C.; Nauen, R. IRAC: Mode of Action Classification and Insecticide Resistance Management. Pestic. Biochem. Physiol. 2015, 121, 122–128. [Google Scholar] [CrossRef]
- Insecticide Resistance Action Committee, IRAC. The IRAC Mode of Action Classification. Available online: https://irac-online.org/mode-of-action/ (accessed on 18 January 2024).
- Numa, S.; Rodríguez, L.; Rodríguez, D.; Coy-Barrera, E. Susceptibility of Tetranychus urticae Koch to an Ethanol Extract of Cnidoscolus aconitifolius Leaves under Laboratory Conditions. SpringerPlus 2015, 4, 338. [Google Scholar] [CrossRef]
- Numa, S.; Rodríguez, L.; Rodríguez, D.; Coy-Barrera, E. Examination of the Acaricidal Effect of a Set of Colombian Native Plants-Derived Extracts against Tetranychus urticae Koch under Laboratory Conditions. J. Biopestic. 2018, 11, 30–37. [Google Scholar] [CrossRef]
- Kousar, T.; Sahito, H.A.; Jatoi, F.A.; Hussain, Z.; Shah, Z.H.; Mangrio, W.M. Resistant Insecticides of Cotton Mealybug, Phenacoccus solenopsis Tinsley) under Laboratory Conditions. J. Entomol. Zool. Stud. 2016, 4, 355–359. [Google Scholar]
- Schymanski, E.L.; Jeon, J.; Gulde, R.; Fenner, K.; Ruff, M.; Singer, H.P.; Hollender, J. Identifying Small Molecules via High Resolution Mass Spectrometry: Communicating Confidence. Environ. Sci. Technol. 2014, 48, 2097–2098. [Google Scholar] [CrossRef] [PubMed]
- Hanwell, M.D.; Curtis, D.E.; Lonie, D.C.; Vandermeersch, T.; Zurek, E.; Hutchison, G.R. Avogadro: An Advanced Semantic Chemical Editor, Visualization, and Analysis Platform. J. Cheminform. 2012, 4, 17. [Google Scholar] [CrossRef] [PubMed]
- The UniProt Consortium UniProt: The Universal Protein Knowledgebase in 2023. Nucleic Acids Res. 2023, 51, D523–D531. [CrossRef]
- Land, H.; Humble, M.S. YASARA: A Tool to Obtain Structural Guidance in Biocatalytic Investigations. In Protein Engineering: Methods and Protocols; Bornscheuer, U.T., Höhne, M., Eds.; Springer: New York, NY, USA, 2018; pp. 43–67. ISBN 978-1-4939-7366-8. [Google Scholar]
- Seeliger, D.; de Groot, B.L. Ligand Docking and Binding Site Analysis with PyMOL and Autodock/Vina. J. Comput. Aided Mol. Des. 2010, 24, 417–422. [Google Scholar] [CrossRef]
- Bitencourt-Ferreira, G.; de Azevedo, W.F. Molegro Virtual Docker for Docking. In Docking Screens for Drug Discovery; de Azevedo, W.F., Jr., Ed.; Springer: New York, NY, USA, 2019; pp. 149–167. ISBN 978-1-4939-9752-7. [Google Scholar]
- Jones, G.; Willett, P.; Glen, R.C.; Leach, A.R.; Taylor, R. Development and Validation of a Genetic Algorithm for Flexible Docking. J. Mol. Biol. 1997, 267, 727–748. [Google Scholar] [CrossRef]
- Palacio-Rodríguez, K.; Lans, I.; Cavasotto, C.N.; Cossio, P. Exponential Consensus Ranking Improves the Outcome in Docking and Receptor Ensemble Docking. Sci. Rep. 2019, 9, 5142. [Google Scholar] [CrossRef]
- Biovia 2016. Discovery Studio Modeling Environment, (version 16.1.0.). Windows; Biovia: San Diego, CA, USA, 2016. [Google Scholar]
- Jeon, Y.W.; Jung, J.W.; Kang, M.R.; Chung, I.K.; Lee, W.T. NMR Studies on Antitumor Drug Candidates, Berberine and Berberrubine. Bull. Korean Chem. Soc. 2002, 23, 391–394. [Google Scholar] [CrossRef]
- Miao, F.; Yang, X.-J.; Zhou, L.; Hu, H.-J.; Zheng, F.; Ding, X.-D.; Sun, D.-M.; Zhou, C.-D.; Sun, W. Structural Modification of Sanguinarine and Chelerythrine and Their Antibacterial Activity. Nat. Prod. Res. 2011, 25, 863–875. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, T.; Suzuki, M. Revision of the Structure of Fagaridine Based on the Comparison of UV and NMR Data of Synthetic Compounds. J. Nat. Prod. 1998, 61, 1263–1267. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.N.; Chauhan, L.; Thankachan, P.P.; Barthwal, R. Quantum Chemical and Nuclear Magnetic Resonance Spectral Studies on Molecular Properties and Electronic Structure of Berberine and Berberrubine. Magn. Reson. Chem. 2007, 45, 647–655. [Google Scholar] [CrossRef] [PubMed]
- de Moura, N.F.; Ribeiro, H.B.; Machado, E.C.S.; Ethur, E.M.; Zanatta, N.; Morel, A.F. Benzophenanthridine Alkaloids from Zanthoxylum rhoifolium. Phytochemistry 1997, 46, 1443–1446. [Google Scholar] [CrossRef]
- Ernst, O.; Zor, T. Linearization of the Bradford Protein Assay. J. Vis. Exp. 2010, 38, e1918. [Google Scholar] [CrossRef]
- Fisher, T.C.; Crane, M.; Callaghan, A. An Optimized Microtiterplate Assay to Detect Acetylcholinesterase Activity in Individual Chironomus riparius Meigen. Environ. Toxicol. Chem. 2000, 19, 1749–1752. [Google Scholar] [CrossRef]
- Vontas, J.G.; Enayati, A.A.; Small, G.J.; Hemingway, J. A Simple Biochemical Assay for Glutathione S-Transferase Activity and Its Possible Field Application for Screening Glutathione S-Transferase-Based Insecticide Resistance. Pestic. Biochem. Physiol. 2000, 68, 184–192. [Google Scholar] [CrossRef]
- Zhu, K.Y.; He, F. Elevated Esterases Exhibiting Arylesterase-like Characteristics in an Organophosphate-Resistant Clone of the Greenbug, Schizaphis graminum (Homoptera: Aphididae). Pestic. Biochem. Physiol. 2000, 67, 155–167. [Google Scholar] [CrossRef]
- Ding, T.-B.; Niu, J.-Z.; Yang, L.-H.; Zhang, K.; Dou, W.; Wang, J.-J. Transcription Profiling of Two Cytochrome P450 Genes Potentially Involved in Acaricide Metabolism in Citrus Red Mite Panonychus citri. Pestic. Biochem. Physiol. 2013, 106, 28–37. [Google Scholar] [CrossRef]
- R Foundation for Statistical Computing. R: A Language and Environment for Statistical Computing, (version 4.3.1.). Windows; R Foundation for Statistical Computing: Vienna, Austria, 2023. [Google Scholar]
- Nicastro, R.L.; Sato, M.E.; Arthur, V.; da Silva, M.Z. Chlorfenapyr Resistance in the Spider Mite Tetranychus urticae: Stability, Cross-Resistance and Monitoring of Resistance. Phytoparasitica 2013, 41, 503–513. [Google Scholar] [CrossRef]
- Khajehali, J.; Van Nieuwenhuyse, P.; Demaeght, P.; Tirry, L.; Van Leeuwen, T. Acaricide Resistance and Resistance Mechanisms in Tetranychus urticae Populations from Rose Greenhouses in the Netherlands. Pest Manag. Sci. 2011, 67, 1424–1433. [Google Scholar] [CrossRef]
- Stumpf, N.; Nauen, R. Biochemical Markers Linked to Abamectin Resistance in Tetranychus urticae (Acari: Tetranychidae). Pestic. Biochem. Physiol. 2002, 72, 111–121. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Lee, S.-H.; Lee, S.-W.; Ahn, Y.-J. Fenpyroximate Resistance in Tetranychus urticae (Acari: Tetranychidae): Cross-Resistance and Biochemical Resistance Mechanisms. Pest Manag. Sci. 2004, 60, 1001–1006. [Google Scholar] [CrossRef]
- Roh, H.S.; Lim, E.G.; Kim, J.; Park, C.G. Acaricidal and Oviposition Deterring Effects of Santalol Identified in Sandalwood Oil against Two-Spotted Spider Mite, Tetranychus urticae Koch (Acari: Tetranychidae). J. Pest Sci. 2011, 84, 495–501. [Google Scholar] [CrossRef]
- Attia, S.; Grissa, K.L.; Ghrabi, Z.G.; Mailleux, A.C.; Lognay, G.; Hance, T. Acaricidal Activity of 31 Essential Oils Extracted from Plants Collected in Tunisia. J. Essent. Oil Res. 2012, 24, 279–288. [Google Scholar] [CrossRef]
- Khambay, B.P.S.; Batty, D.; Cahill, M.; Denholm, I.; Mead-Briggs, M.; Vinall, S.; Niemeyer, H.M.; Simmonds, M.S.J. Isolation, Characterization, and Biological Activity of Naphthoquinones from Calceolaria Andina L. J. Agric. Food Chem. 1999, 47, 770–775. [Google Scholar] [CrossRef]
- Latif, Z.; Craven, L.; Hartley, T.G.; Kemp, B.R.; Potter, J.; Rice, M.J.; Waigh, R.D.; Waterman, P.G. An Insecticidal Quassinoid from the New Australian Species Quassia sp. aff. Bidwillii. Biochem. Syst. Ecol. 2000, 28, 183–184. [Google Scholar] [CrossRef]
- Antonious, G.F.; Meyer, J.E.; Snyder, J.C. Toxicity and Repellency of Hot Pepper Extracts to Spider Mite, Tetranychus urticae Koch. J. Environ. Sci. Health Part B 2006, 41, 1383–1391. [Google Scholar] [CrossRef] [PubMed]
- Hiremath, I.; Ahn, Y.; Kim, S.; Choi, B.; Cho, J. Insecticidal and Acaricidal Activities of African Plant Extracts against the Brown Planthopper and Two-Spotted Spider Mite. Korean J. Appl. Entomol. 1995, 34, 200–205. [Google Scholar]
- Chermenskaya, T.D.; Stepanycheva, E.A.; Shchenikova, A.V.; Chakaeva, A.S. Insectoacaricidal and Deterrent Activities of Extracts of Kyrgyzstan Plants against Three Agricultural Pests. Ind. Crops Prod. 2010, 32, 157–163. [Google Scholar] [CrossRef]
- da Camara, C.A.G.; Akhtar, Y.; Isman, M.B.; Seffrin, R.C.; Born, F.S. Repellent Activity of Essential Oils from Two Species of Citrus against Tetranychus urticae in the Laboratory and Greenhouse. Crop Prot. 2015, 74, 110–115. [Google Scholar] [CrossRef]
- Marčić, D.; Međo, I. Effects of Azadirachtin-A (NeemAzal-T/S) on Tetranychus urticae (Acari: Tetranychidae). Syst. Appl. Acarol. 2015, 20, 25–38. [Google Scholar] [CrossRef]
- Van Leeuwen, T.; Van Pottelberge, S.; Tirry, L. Biochemical Analysis of a Chlorfenapyr-Selected Resistant Strain of Tetranychus urticae Koch. Pest Manag. Sci. 2006, 62, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhao, S.; Shi, L.; Xu, Z.; He, L. Resistance Selection and Biochemical Mechanism of Resistance against Cyflumetofen in Tetranychus cinnabarinus (Boisduval). Pestic. Biochem. Physiol. 2014, 111, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, N.; Sasama, Y.; Takahashi, N.; Ikemi, N. Cyflumetofen, a Novel Acaricide—Its Mode of Action and Selectivity. Pest Manag. Sci. 2013, 69, 1080–1084. [Google Scholar] [CrossRef] [PubMed]
- Soetopo, D.; Alouw, J.C. Biopesticide Development & Registration: Challenges & Strategies. IOP Conf. Ser. Earth Environ. Sci. 2023, 1179, 012003. [Google Scholar] [CrossRef]
- Damalas, C.A.; Koutroubas, S.D. Current Status and Recent Developments in Biopesticide Use. Agriculture 2018, 8, 13. [Google Scholar] [CrossRef]
- Saha, S.; Thosar, R.; Kabade, S.; Pawar, K.; Banerjee, K. Biopesticides. In Trajectory of 75 Years of Indian Agriculture after Independence; Ghosh, P.K., Das, A., Saxena, R., Banerjee, K., Kar, G., Vijay, D., Eds.; Springer Nature: Singapore, 2023; pp. 675–703. ISBN 978-981-19799-7-2. [Google Scholar]
- Reynel, C. Zanthoxylum (Rutaceae); Flora Neotropica Series; New York Botanical Garden Press: Bronx, NY, USA, 2017; Volume 117, 264p. [Google Scholar]
- Koul, O. Chapter 1—Biopesticides: Commercial Opportunities and Challenges. In Development and Commercialization of Biopesticides; Koul, O., Ed.; Academic Press: Cambridge, MA, USA, 2023; pp. 1–23. ISBN 978-0-323-95290-3. [Google Scholar]
- Nooreen, Z.; Tandon, S.; Yadav, N.P.; Kumar, P.; Xuan, T.D.; Ahmad, A. Zanthoxylum: A Review of Its Traditional Uses, Naturally Occurring Constituents and Pharmacological Properties. Curr. Org. Chem. 2019, 23, 1307–1341. [Google Scholar] [CrossRef]
- Wei, W.-J.; Chen, X.-H.; Guo, T.; Liu, X.-Q.; Zhao, Y.; Wang, L.-L.; Lan, J.-X.; Li, H.-W.; Si, Y.-P.; Wang, Z.-M. A Review on Classification and Biological Activities of Alkaloids from the Genus Zanthoxylum Species. Mini Rev. Med. Chem. 2021, 21, 336–361. [Google Scholar] [CrossRef]
- Nechepurenko, I.V.; Salakhutdinov, N.F.; Tolstikov, G.A. Berberine: Chemistry and Biological Activity. Chem. Sustain. Dev. 2010, 18, 1–23. [Google Scholar]
- Och, A.; Podgórski, R.; Nowak, R. Biological Activity of Berberine—A Summary Update. Toxins 2020, 12, 713. [Google Scholar] [CrossRef]
- Ma, D.-L.; Chan, D.S.-H.; Leung, C.-H. Molecular Docking for Virtual Screening of Natural Product Databases. Chem. Sci. 2011, 2, 1656–1665. [Google Scholar] [CrossRef]
- Senol Deniz, S.F.; Ekhteiari Salmas, R.; Emerce, E.; Sener, B.; Orhan, E.I. Cholinesterase Inhibitory and In Silico Toxicity Assessment of Thirty-Four Isoquinoline Alkaloids—Berberine as the Lead Compound. Former. Curr. Drug Targets—CNS Neurol. Disord. 2023, 23, 1–11. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Commercial Product | Effect | Active Ingredients | Mechanism of Action a |
|---|---|---|---|
| Quorum | Fungicide | chlorothalonil/dimethomorph | Lipid biosynthesis inhibition |
| Connect Duo | Insecticide | imidacloprid/β-cyfluthrin | Competitive modulator of acetylcholine receptors/sodium channel modulator |
| Abamectin | Acaricide | abamectin | Allosteric modulator of chlorine channels |
| Preza | Insecticide | cyantraniliprole | Modulator of ryanodine receptors |
| Copper oxychloride | Fungicide | copper oxychloride | Fixation of Cu2+ ions in enzymes |
| Fitotripen | Fungicide | Trichoderma sp. | Antagonism and mycoparasitism |
| Mesurol | Insecticide | methiocarb | Acetylcholinesterase inhibitor |
| Topas | Fungicide | penconazole | Inhibition of sterol synthesis |
| Athrin Brio | Insecticide | λ-cyhalothrin | Sodium channel modulator |
| Plaguicontrol | Insecticide | garlic and chili extracts | Unknown |
| No a | tR b | [M+H]+ (m/z) | am c (m/z) | Error d | Annotation e |
|---|---|---|---|---|---|
| 1 | 16.7 | 342.1 | 342.1718 | −3.71 | tetrahydrocolumbamine |
| 2 | 22.2 | 354.2 | 354.1721 | −4.43 | N-methylcanadine |
| 3 | 25.3 | 322.1 | 322.1088 | −2.70 | berberrubine |
| 4 | 25.9 | 348.1 | 348.1225 | 3.10 | chelerythrine |
| 5 | 28.1 | 334.1 | 334.1066 | 3.98 | fagaridine |
| 6 | 29.3 | 336.1 | 336.1247 | −3.33 | berberine |
| 7 | 34.2 | 230.1 | 230.0809 | 3.56 | γ-fagarine |
| 8 | 38.8 | 386.2 | 386.1981 | −3.50 | muramine |
| 9 | 40.6 | 320.1 | 320.0933 | −3.19 | zanthoxyline |
| 10 | 41.4 | 350.2 | 350.1383 | 2.66 | dihydrochelerythrine |
| 11 | 44.1 | 334.1 | 334.1071 | 2.48 | norchelerythrine |
| Enzymes a | ||||
|---|---|---|---|---|
| GST b | PMO c | Est b | AChE b | |
| Untreated mites | 2118 ± 56 A | 463 ± 34 B | 438 ± 26 A | 1257 ± 48 A |
| Z. schreberi-treated mites | 235 ± 13 B | 1345 ± 41 A | 445 ± 31 A | 578 ± 41 B |
| TuAChE a | TuGST a | TuPMO c | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No b | V c | M c | G c | ES d | ECR e | No b | V c | M c | G c | ES d | ECR e | No b | V c | M c | G c | ES d | ECR e |
| 12 f | −8.70 | −121.7 | 70.2 | 0.239 | 1 | 13 g | −8.60 | −138.2 | 52.7 | 0.233 | 1 | 14 h | −8.80 | −120.9 | 71.8 | 0.263 | 1 |
| 6 | −6.90 | −124.8 | 72.4 | 0.239 | 2 | 6 | −8.70 | −100.3 | 50.4 | 0.230 | 2 | 7 | −3.30 | −89.7 | 54.4 | 0.254 | 2 |
| 9 | −7.10 | −122.4 | 63.9 | 0.209 | 3 | 2 | −8.60 | −106.4 | 49.2 | 0.223 | 3 | 9 | −1.50 | −32.4 | 42.7 | 0.189 | 3 |
| 4 | −7.20 | −113.1 | 68.8 | 0.201 | 4 | 3 | −8.90 | −99.2 | 48.9 | 0.218 | 4 | 5 | −1.40 | −22.7 | 43.5 | 0.179 | 4 |
| 2 | −4.30 | −126.7 | 45.3 | 0.173 | 5 | 1 | −7.20 | −98.2 | 54.4 | 0.184 | 5 | 11 | −1.20 | −6.5 | 50.4 | 0.174 | 5 |
| 5 | −4.10 | −115.3 | 66.6 | 0.163 | 6 | 11 | −8.50 | −96.7 | 48.4 | 0.159 | 6 | 3 | −1.30 | −10.1 | 48.8 | 0.171 | 6 |
| 8 | −5.90 | −119.5 | 39.4 | 0.151 | 7 | 9 | −8.60 | −85.5 | 43.6 | 0.137 | 7 | 4 | −1.10 | −16.7 | 46.5 | 0.149 | 7 |
| 3 | −4.10 | −114.7 | 52.9 | 0.144 | 8 | 10 | −8.10 | −95.4 | 44.6 | 0.136 | 8 | 10 | −1.10 | −17.5 | 42.2 | 0.147 | 8 |
| 7 | −5.60 | −102.2 | 51.2 | 0.130 | 9 | 4 | −7.90 | −92.3 | 46.5 | 0.131 | 9 | 2 | −0.90 | 3.0 | 49.2 | 0.141 | 9 |
| 1 | −3.10 | −103.6 | 54.8 | 0.121 | 10 | 5 | −8.00 | −88.4 | 44.8 | 0.126 | 10 | 8 | −0.50 | 57.9 | 47.7 | 0.115 | 10 |
| 11 | −3.10 | −110.6 | 50.8 | 0.111 | 11 | 8 | −7.30 | −85.5 | 48.8 | 0.125 | 11 | 6 | −1.00 | 3.1 | 9.0 | 0.111 | 11 |
| 10 | −3.60 | −109.6 | 49.5 | 0.110 | 12 | 7 | −6.50 | −73.6 | 37.0 | 0.090 | 12 | 1 | −0.90 | 5.1 | 2.4 | 0.100 | 12 |
| Enzymes a | ||||
|---|---|---|---|---|
| Subs. b | GST | PMO | Est | AChE |
| extract | 8.65 (8.44–8.78) | 125 (119–130) | >1000 | 15.8 (15.1–16.7) |
| 3 | 5.61 (5.49–5.77) | 62.7 (60.3–64.8) | >300 | 12.4 (11.8–13.5) |
| 4 | 11.71 (11.53–11.89) | 89.6 (88.8–90.8) | >300 | 1.86 (1.69–1.98) |
| 5 | 112 (99.3–121) | 20.9 (20.1–22.3) | >300 | 14.6 (13.9–15.7) |
| 6 | 1.24 (1.05–1.45) | 386 (371–399) | >300 | 0.325 (0.306–0.351) |
| 9 | 21.5 (20.9–22.3) | 8.37 (8.23–8.49) | 289 (263–299) | 3.53 (3.21–3.76) |
| donepezil | - | - | - | 0.0135 (0.0118–0.0154) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rincón, R.A.; Rodríguez, D.; Coy-Barrera, E. Susceptibility of Tetranychus urticae to the Alkaloidal Extract of Zanthoxylum schreberi Bark: Phenotypic and Biochemical Insights for Biotechnological Exploitation. BioTech 2024, 13, 5. https://doi.org/10.3390/biotech13010005
Rincón RA, Rodríguez D, Coy-Barrera E. Susceptibility of Tetranychus urticae to the Alkaloidal Extract of Zanthoxylum schreberi Bark: Phenotypic and Biochemical Insights for Biotechnological Exploitation. BioTech. 2024; 13(1):5. https://doi.org/10.3390/biotech13010005
Chicago/Turabian StyleRincón, Ricardo A., Daniel Rodríguez, and Ericsson Coy-Barrera. 2024. "Susceptibility of Tetranychus urticae to the Alkaloidal Extract of Zanthoxylum schreberi Bark: Phenotypic and Biochemical Insights for Biotechnological Exploitation" BioTech 13, no. 1: 5. https://doi.org/10.3390/biotech13010005
APA StyleRincón, R. A., Rodríguez, D., & Coy-Barrera, E. (2024). Susceptibility of Tetranychus urticae to the Alkaloidal Extract of Zanthoxylum schreberi Bark: Phenotypic and Biochemical Insights for Biotechnological Exploitation. BioTech, 13(1), 5. https://doi.org/10.3390/biotech13010005