Effects of Pseudomonas sp. OBA 2.4.1 on Growth and Tolerance to Cadmium Stress in Pisum sativum L.


Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation of Pseudomonas sp. OBA 2.4.1
2.2. Molecular Genetic Identification of Bacterial Strains
2.3. Biofilm Formation on Inert Surfaces
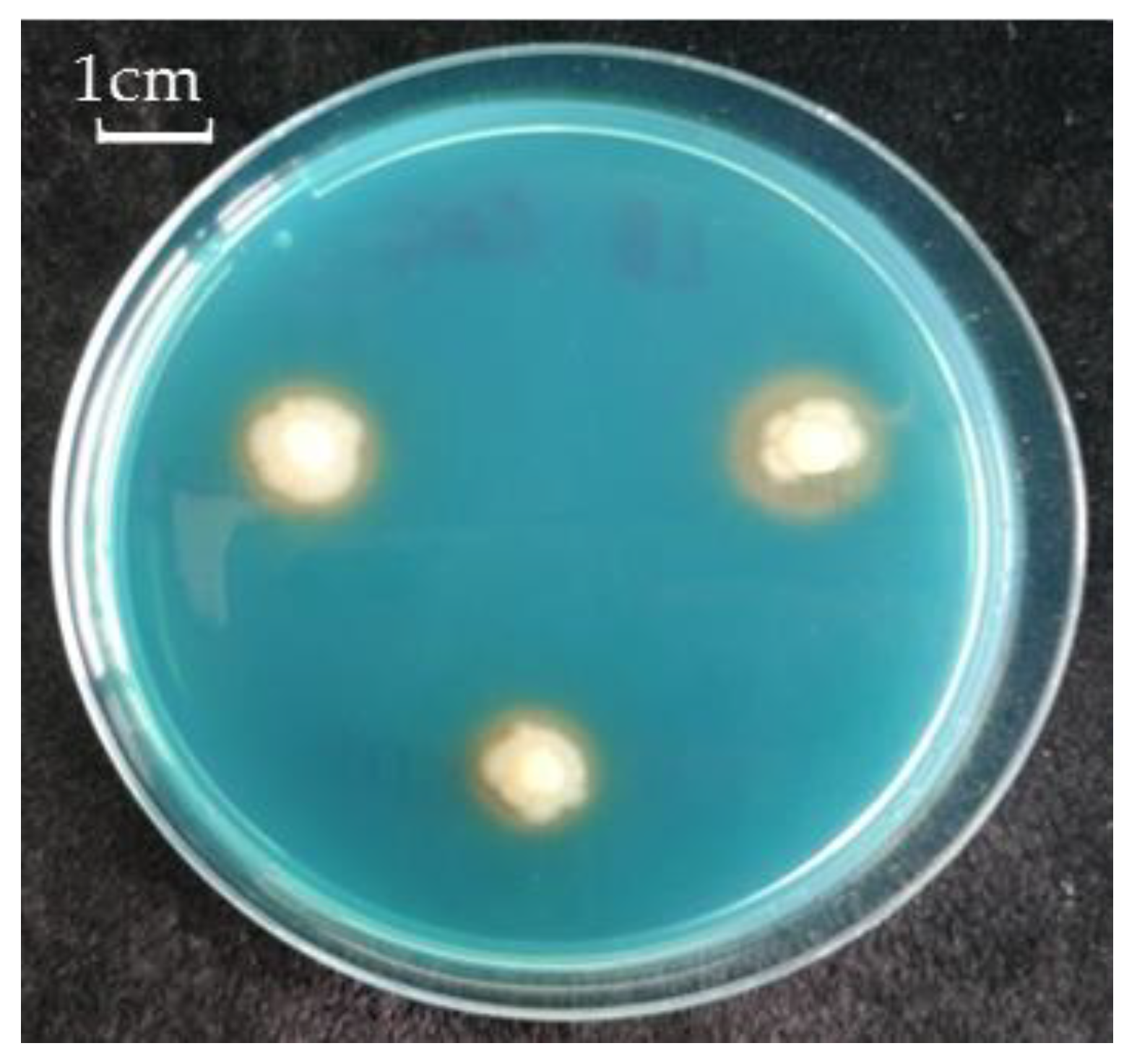
2.4. Phosphate Mobilization and Siderophore Activity of Pseudomonas sp. OBA 2.4.1
2.5. Determination of H2O2 Content
2.6. Determination of the Malondialdehyde (MDA) Content
2.7. Treatment of Plants with Pseudomonas sp. OBA 2.4.1
2.8. Statistical Analysis
3. Results
3.1. Identification of the Strain OBA 2.4.1 and Its Main PGP Traits
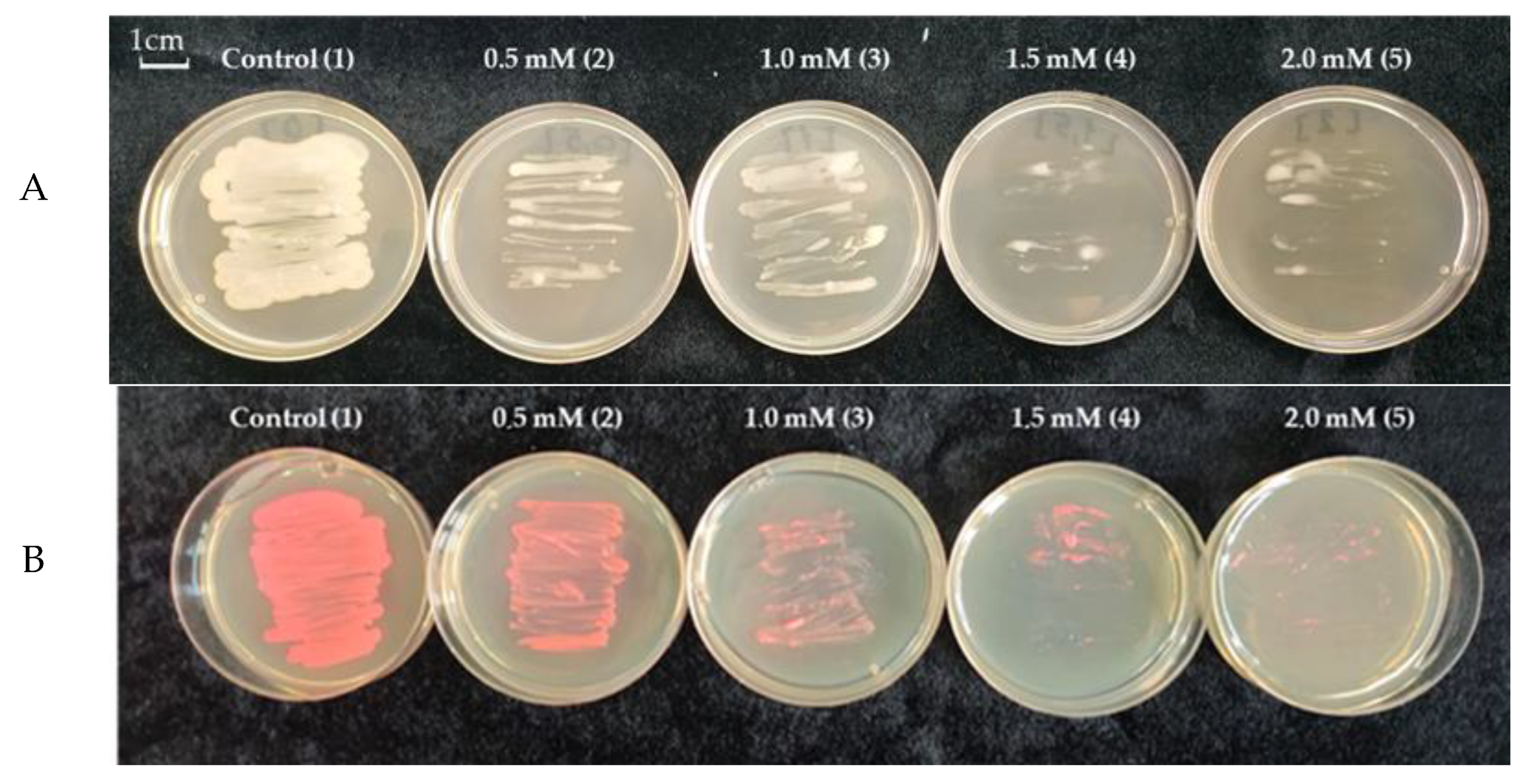
3.2. Growth Analysis of Pseudomonas sp. OBA 2.4.1 under Cadmium Stress (0.5, 1, 1.5, 2 mM CdCl2)
3.3. Effect of Pseudomonas sp. OBA 2.4.1 Strain on Pea Growth under Cd Stress
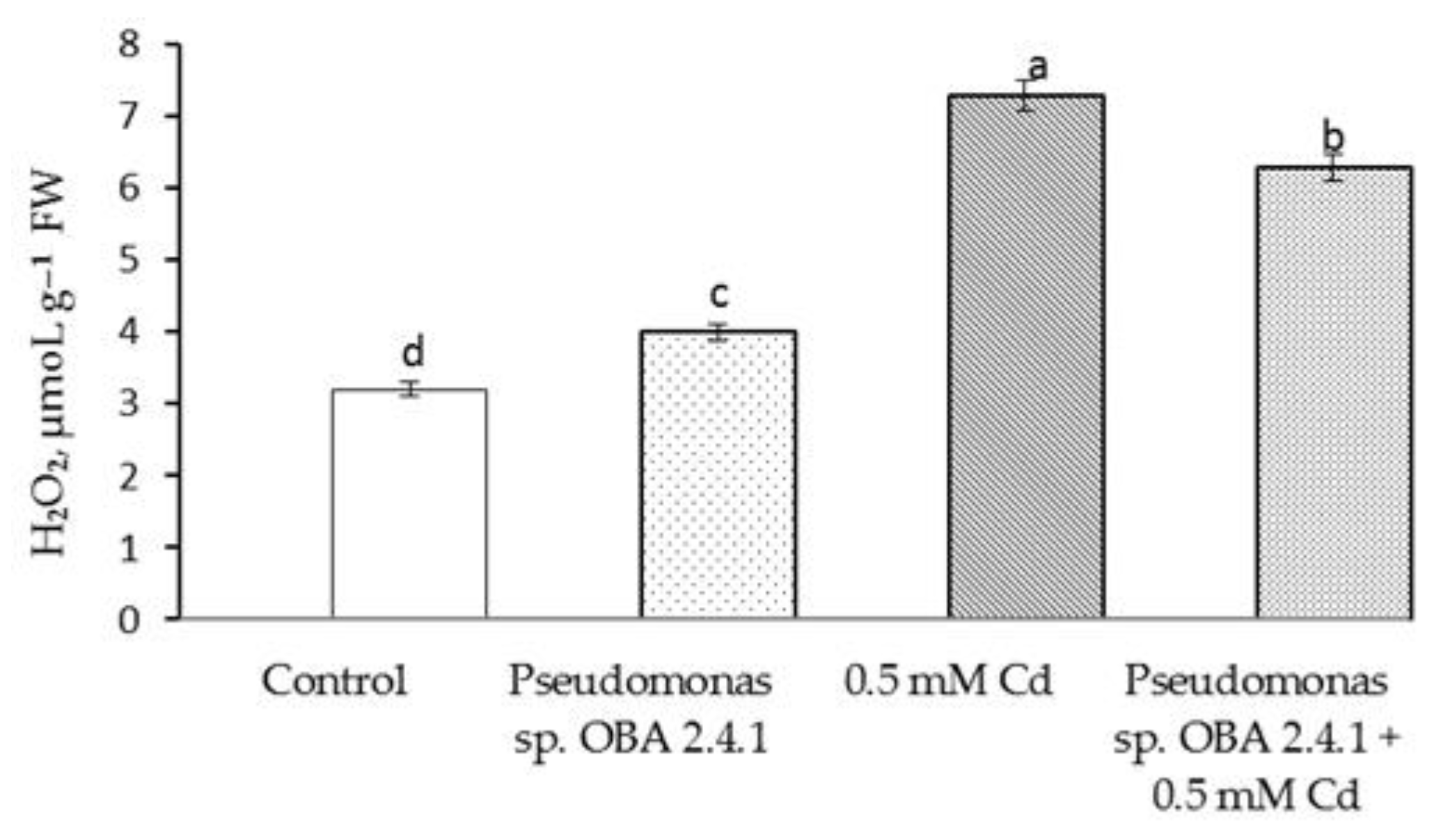
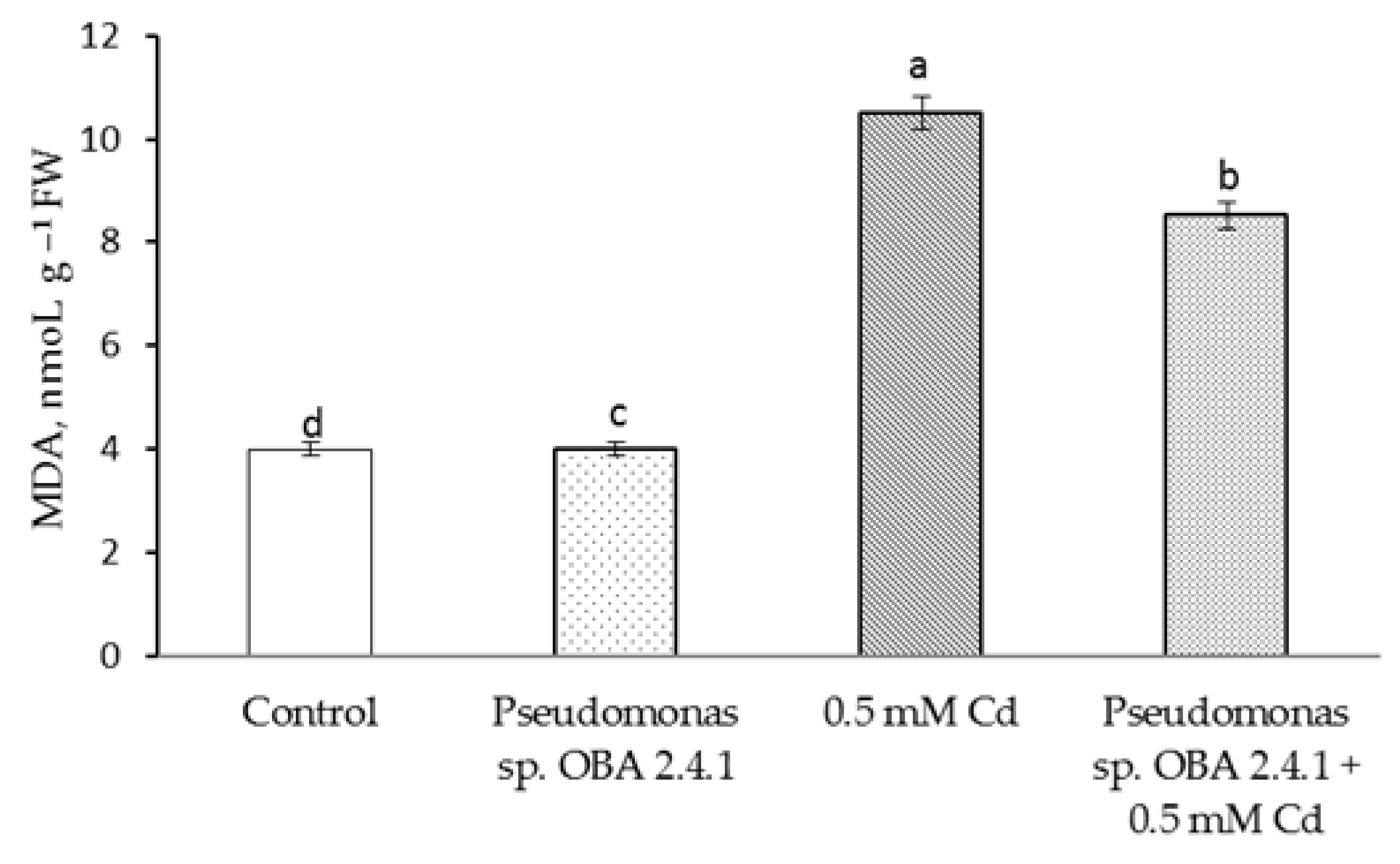
3.4. Effects of Strain Pseudomonas sp. OBA 2.4.1 on the Content H2O2 and MDA in Pea Seedlings under Cd Stress
3.5. Bacterial Colonization of Pisum sativum L. Plant Roots
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhu, T.; Li, L.; Duan, Q.; Liu, X.; Chen, M. Progress in our understanding of plant responses to the stress of heavy metal cadmium. Plant Signal. Behav. 2021, 16, 1836884. [Google Scholar] [CrossRef] [PubMed]
- Vicas, S.I.; Laslo, V.; Timar, A.V.; Balta, C.; Herman, H.; Ciceu, A.; Gharbia, S.; Rosu, M.; Mladin, B.; Chiana, L.; et al. Nano selenium—Enriched probiotics as functional food products against cadmium liver toxicity. Materials 2021, 14, 2257. [Google Scholar] [CrossRef]
- Rai, R.; Agrawal, M.; Agrawal, S.B. Impact of heavy metals on physiological processes of plants: With special reference to photosynthetic system. In Plant Responses to Xenobiotics; Springer: Singapore, Singapore, 2016; pp. 127–140. [Google Scholar] [CrossRef]
- Choi, Y.E.; Harada, E.; Wada, M.; Tsuboi, H.; Morita, Y.; Kusano, T.; Sano, H. Detoxification of cadmium in tobacco plants: Formation and active excretion of crystals containing cadmium and calcium through trichomes. Planta 2001, 213, 45–50. [Google Scholar] [CrossRef]
- Asgher, M.; Khan, M.I.; Anjum, N.A.; Khan, N.A. Minimising toxicity of cadmium in plants—Role of plant growth regulators. Protoplasma 2015, 252, 399–413. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.S.; Dietz, K.J. The relationship between metal toxicity and cellular redox imbalance. Trends Plant Sci. 2009, 14, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Sandalio, L.M.; Dalurzo, H.C.; Gomez, M.; Romero-Puetas, M.C.; del Rio, L.A. Cadmium-induced changes in the growth and oxidative metabolism of pea plants. J. Exp. Bot. 2001, 364, 2115–2126. [Google Scholar] [CrossRef] [PubMed]
- Kochetkov, V.V.; Anokhina, T.O.; Siunova, T.V.; Sizova, O.I.; Boronin, A.M. Formation and prospects of organic farming in Russia. 2018. Available online: http://potatoveg.ru/en/main-topic/formation-and-prospects-of-organic-farming-in-russia-review.html (accessed on 28 December 2022).
- Sharma, R.K.; Archana, G. Cadmium minimization in food crops by cadmium resistant plant growth promoting rhizobacteria. Appl. Soil Ecol. 2016, 107, 66–78. [Google Scholar] [CrossRef]
- Cardarelli, M.; Woo, S.L.; Rouphael, Y.; Colla, G. Seed treatments with microorganisms can have a siostimulant effect by Influencing germination and seedling growth of crops. Plants 2022, 11, 259. [Google Scholar] [CrossRef]
- Liu, A.; Wang, W.; Zheng, X.; Chen, X.; Fu, W.; Wang, G.; Ji, J.; Jin, C.; Guan, C. Improvement of the Cd and Zn phytoremediation efficiency of rice (Oryza sativa) through the inoculation of a metal-resistant PGPR strain. Chemosphere 2022, 302, 134900. [Google Scholar] [CrossRef]
- Gisbert, C.; Clemente, R.; Navarro-Aviñó, J.; Baixauli, C.; Ginér, A.; Serrano, R.; Walker, D.J.; Bernal, M.P. Tolerance and accumulation of heavy metals by Brassicaceae species grown in contaminated soils from mediterranean regions of Spain. Environ. Exp. Bot. 2006, 56, 19–27. [Google Scholar] [CrossRef]
- Vamerali, T.; Bandiera, M.; Mosca, G. Field crops for phytoremediation of metal-contaminated land. A review. Environ. Chem. Lett. 2009, 8, 1–17. [Google Scholar] [CrossRef]
- Dar, M.I.; Khan, F.A.; Rehman, F.; Masoodi, A.; Ansari, A.A.; Varshney, D.; Naushin, F.; Naikoo, M.I. Roles of brassicaceae in phytoremediation of metals and metalloids. In Phytoremediation; Ansari, A., Gill, S., Gill, R., Lanza, G., Newman, L., Eds.; Springer: Cham, Switzerland, 2014; pp. 201–215. [Google Scholar] [CrossRef]
- Inouhe, M.; Ninomiya, S.; Tohoyama, H.; Joho, M.; Murayama, T. Different characteristics of root in the cadmium-tolerance and cd-binding complex formation between mono- and dicotyledonous plants. J. Plant Res. 1994, 107, 201–207. [Google Scholar] [CrossRef]
- Mazen, A.M.A. Assessment of heavy metal accumulation and performance of some physiological parameters in Zea Mays L. and Vicia Faba L. grown on soil amended by sewage sludge resulting from sewage water treatment in the state of Qatar. Qatar Univ. Sci. J. 1995, 5, 353–359. [Google Scholar]
- Belimov, A.A.; Shaposhnikov, A.I.; Azarova, T.S.; Makarova, N.M.; Safronova, V.I.; Litvinskiy, V.A.; Nosikov, V.V.; Zavalin, A.A.; Tikhonovich, I.A. Microbial consortium of PGPR, rhizobia and arbuscular mycorrhizal fungus makes pea mutant SGECdt Comparable with Indian mustard in cadmium tolerance and accumulation. Plants 2020, 9, 975. [Google Scholar] [CrossRef]
- Artursson, V.; Finlay, R.D.; Jansson, J.K. Interactions between arbuscular mycorrhizal fungi and bacteria and their potential for stimulating plant growth. Environ. Microbiol. 2006, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- De-Bashan, L.E.; Hernandez, J.-P.; Bashan, Y. The potential contribution of plant growth-promoting bacteria to reduce environmental degradation—A comprehensive evaluation. Appl. Soil Ecol. 2012, 61, 171–189. [Google Scholar] [CrossRef]
- Gomez-Sagasti, M.T.; Marino, D. PGPRs and nitrogen-fixing legumes: A perfect team for ecient Cd phytoremediation? Front. Plant Sci. 2015, 6, 81. [Google Scholar] [CrossRef][Green Version]
- Ullah, A.; Heng, S.; Farooq, M.; Munis, H.; Fahad, S.; Yang, X. Phytoremediation of heavy metals assisted by plant growth promoting (PGP) bacteria: A review. Environ. Exp. Bot. 2015, 117, 28–40. [Google Scholar] [CrossRef]
- Khakimova, L.R.; Chubukova, O.V.; Muryasova, A.R.; Simoroz, E.V.; Chumakova, A.K.; Vershinina, Z.R. Impact of Pseudomonas spp. on Medicago sativa L. plants under the inhibitory effect of cadmium salts. Taurida Her. Agrar. Sci. 2022, 2, 155–163. Available online: https://elibrary.ru/item.asp?id=49231774 (accessed on 20 May 2022).
- Desoky, E.S.M.; Merwad, A.R.M.; Semida, W.M.; Ibrahim, S.A.; El-Saadony, M.T.; Rady, M.M. Heavy metals-resistant bacteria (HM-RB): Potential bioremediators of heavy metalsstressed Spinacia oleracea plant. Ecotoxicol. Environ. Saf. 2020, 198, 110685. [Google Scholar] [CrossRef]
- Chubukova, O.V.; Vershinina, Z.R.; Matnyazov, R.T.; Baymiev, A.K.; Baymiev, A.K. Creation of an inducible vector system based on the rhizobia nodA gene promoter. Ekol. Genet. 2021, 19, 13–21. [Google Scholar] [CrossRef]
- Baymiev, A.K.; Akimova, E.S.; Gumenko, R.S.; Vladimirova, A.A.; Muldashev, A.A.; Chemeris, A.V.; Baymiev, A.K. Genetic diversity and phylogeny of root nodule bacteria isolated from nodules of plants of the Lupinaster genus inhabiting the southern Urals. Russ. J. Genet. 2019, 55, 45–51. [Google Scholar] [CrossRef]
- Mulet, M.; Bennasar, A.; Lalucat, J.; García-Valdés, E. An rpoD-based PCR procedure for the identification of Pseudomonas species and for their detection in environmental samples. Mol. Cell Probes. 2009, 23, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Chubukova, O.V.; Khakimova, L.R.; Akimova, E.S.; Vershinina, Z.R. Phylogeny and Properties of New Pseudomonas spp. from the rhizosphere of Southern Ural leguminous plants. Microbiology 2022, 91, 537–546. [Google Scholar] [CrossRef]
- Baymiev, A.K.; Yamidanov, R.S.; Matniyazov, R.T.; Blagova, D.K.; Chemeris, A.V. Preparation of fluorescent labeled nodule bacteria strains of wild legumes for their detection in vivo and in vitro. Mol. Biol. 2011, 45, 904–910. [Google Scholar] [CrossRef]
- Vershinina, Z.R.; Chubukova, O.V.; Nikonorov, Y.M.; Khakimova, L.R.; Lavina, A.M.; Karimova, L.R.; Baymiev, A.K. Effect of rosr gene overexpression on biofilm formation by Rhizobium leguminosarum. Microbiology 2021, 90, 198–209. [Google Scholar] [CrossRef]
- Yegorshina, A.A.; Khayrullin, R.M.; Lukyantsev, M.A.; Kuramshina, Z.M.; Smirnova, Y.V. Phosphate-mobilizing activity of the endophytic Bacillus subtilis strains and their effect on wheat roots micorrhization ratio. J. Sib. Fed. Univ. Biol. 2011, 4, 172–182. [Google Scholar]
- Schwynan, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Maslennikova, D.; Nasyrova, K.; Chubukova, O.; Akimova, E.; Baymiev, A.; Blagova, D.; Ibragimov, A.; Lastochkina, O. Effects of Rhizobium leguminosarum Thy2 on the growth and tolerance to cadmium stress of wheat plants. Life 2022, 12, 1675. [Google Scholar] [CrossRef] [PubMed]
- Bindschedler, L.V.; Minibaeva, F.; Gardner, S.L.; Gerrish, C.; Davies, D.R.; Bolwell, G.P. Early Signalling events in the apoplastic oxidative burst in suspension cultured french bean cells involve cAMP and Ca2+. New Phytol. 2001, 151, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Maslennikova, D.; Lastochkina, O. Contribution of ascorbate and glutathione in endobacteria bacillus subtilis- mediated drought tolerance in two Triticum aestivum L. genotypes contrasting in drought sensitivity. Plants 2021, 10, 2557. [Google Scholar] [CrossRef] [PubMed]
- Teitzel, G.M.; Parsek, M.R. Heavy metal resistance of biofilm and planktonic Pseudomonas aeruginosa. Appl. Environ. Microbiol. 2003, 69, 2313–2320. [Google Scholar] [CrossRef]
- Pishchik, V.N.; Vorob'ev, N.I.; Provorov, N.A.; Khomyakov, Y.V. Mechanisms of plant and microbial adaptation to heavy metals in plant–microbial systems. Microbiology 2016, 85, 257–271. [Google Scholar] [CrossRef]
- Khan, N.F.; Rasool, A.; Mansoor, S.; Saleem, S.; Baba, T.R.; Haq, S.M.; Rehman, S.A.; Adetunji, C.O.; Popesc, S.M. Potential applications of Rhizobacteria as eco-friendly biological control, plant growth promotion and soil metal bioremediation. In Sustainable Crop Production Recent Advances; Meena, V., Choudhary, M., Meena, S.K., Yadav, R.P., Eds.; IntechOpen Limited: London, UK, 2022; pp. 104–170. [Google Scholar] [CrossRef]
- Meena, M.; Swapnil, P.; Divyanshu, K.; Kumar, S.; Tripathi, Y.N.; Zehra, A.; Marwal, A.; Upadhyay, R.S. PGPR-mediated induction of systemic resistance and physiochemical alterations in plants against the pathogens: Current perspective. J. Basic Microbiol. 2020, 60, 828–861. [Google Scholar] [CrossRef]
- Zulfiqar, U.; Ayub, A.; Hussain, S.; Waraich, E.A.; El-Esawi, M.A.; Ishfaq, M.; Ahmad, M.; Ali, N.; Maqsood, M.F. Cadmium toxicity in plants: Recent progress on morpho-physiological effects and remediation strategies. J. Soil Sci. Plant Nutr. 2022, 22, 212–269. [Google Scholar] [CrossRef]
- Jalil, A.; Selles, F.; Clarke, J.M. Effects of cadmium on growth and the uptake of cadmium and other elements by durum wheat. Plant Nutr. 1994, 17, 1839–1895. [Google Scholar] [CrossRef]
- Yang, J.F.; Bu, Y.S.; Guo, X.Y. Research on effects of soil exogenous cadmium and lead pollution on rape growth. Shanxi Agric. Sci. 2005, 3, 26–28. [Google Scholar]
- Liu, J.X. Effects of cadmium and zinc interaction on corn seedling physiological and biochemical characteristics. Yi Chun Coll. J. 2004, 26, 55–57. [Google Scholar]
- Da-Lin, L.; Kai-Qi, H.; Jing-Jing, M.; Wei-Wei, Q.; Xiu-Ping, W.; Shu-Pan, Z. Effects of cadmium on the growth and physiological characteristics of sorghum plants. Afr. J. Biotechnol. 2011, 10, 15770–15776. [Google Scholar] [CrossRef]
- Raklami, A.; Oufdou, K.; Tahiri, A.I.; Mateos-Naranjo, E.; Navarro-Torre, S.; Rodríguez-Llorente, I.D.; Meddich, A.; Redondo-Gómez, S.; Pajuelo, E. Safe cultivation of Medicago sativa in metal-polluted soils from semi-arid regions assisted by heat- and metallo-resistant PGPR. Microorganisms 2019, 7, 212. [Google Scholar] [CrossRef]
- Konkolewska, A.; Piechalak, A.; Ciszewska, L.; Antos-Krzemińska, N.; Skrzypczak, T.; Hanć, A.; Sitko, K.; Małkowski, E.; Barałkiewicz, D.; Małecka, A. Combined use of companion planting and PGPR for the assisted phytoextraction of trace metals (Zn, Pb, Cd). Environ. Sci. Pollut. Res. Int. 2020, 27, 13809–13825. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R. Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol. Res. 2014, 169, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Goswami, D.; Thakker, J.N.; Dhandhukia, P.C. Portraying mechanics of plant growth promoting rhizobacteria (PGPR): A review. Cogent Food Agric. 2016, 2, 1127500. [Google Scholar] [CrossRef]
- Navarro-Torre, S.; Barcia-Piedras, J.M.; Caviedes, M.A.; Pajuelo, E.; Redondo-Gómez, S.; Rodríguez-Llorente, I.D.; Mateos-Naranjo, E. Bioaugmentation with bacteria selected from the microbiome enhances Arthrocnemum macrostachyum metal accumulation and tolerance. Mar. Pollut. Bull. 2017, 117, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Paredes-Páliz, K.; Rodríguez-Vázquez, R.; Duarte, B.; Caviedes, M.A.; Mateos-Naranjo, E.; Redondo-Gómez, S.; Caçador, M.I.; Rodríguez-Llorente, I.D.; Pajuelo, E. Investigating themechanisms underlying phytoprotection by plant growth-promoting rhizobacteria in Spartina densifloraunder metal stress. Plant Biol. 2018, 20, 497–506. [Google Scholar] [CrossRef]
- Mesnoua, M.; Mateos-Naranjo, E.; Pérez-Romero, J.A.; Barcia-Piedras, J.M.; Lotmani, B.; Redondo-Gómez, S. Combined effect of Cr-toxicity and temperature rise on physiological and biochemical responses of Atriplex halimus L. Plant Physiol. Biochem. 2018, 132, 675–682. [Google Scholar] [CrossRef]
- Odoh, C.K.; Eze, C.N.; Apki, U.K.; Unah, V.U. Plant growth promoting rhizobacteria (PGPR): A novel agent for sustainable food production. Am. J. Agric. Biol. Sci. 2019, 14, 35–54. [Google Scholar] [CrossRef]
- Shabayev, V.P.; Ostroumov, V.E.; Bocharnikova, E.A. Plant and soil mechanisms of plant resistance to cd toxicity under application of plant growth promoting rhizobacterium in soil contaminated with heavy metal. Biol. Bull. 2019, 46, 381–386. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentrations of Cd, mM | Without inoculation, mm | Inoculation with Pseudomonas sp. OBA 2.4.1, mm |
|---|---|---|
| 0 (control) | 11.8 ± 0.35 | 30.9 ± 0.93 |
| 0.1 | 14.5 ± 0.43 | 23.5 ± 0.70 |
| 0.2 | 18.6 ± 0.55 | 22.4 ± 0.67 |
| 0.3 | 15.6 ± 0.46 | 21.3 ± 0.63 |
| 0.4 | 12.7 ± 0.38 | 19.2 ± 0.57 |
| 0.5 | 8.6 ± 0.25 | 18.8 ± 0.56 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khakimova, L.; Chubukova, O.; Vershinina, Z.; Maslennikova, D. Effects of Pseudomonas sp. OBA 2.4.1 on Growth and Tolerance to Cadmium Stress in Pisum sativum L. BioTech 2023, 12, 5. https://doi.org/10.3390/biotech12010005
Khakimova L, Chubukova O, Vershinina Z, Maslennikova D. Effects of Pseudomonas sp. OBA 2.4.1 on Growth and Tolerance to Cadmium Stress in Pisum sativum L. BioTech. 2023; 12(1):5. https://doi.org/10.3390/biotech12010005
Chicago/Turabian StyleKhakimova, Liliya, Olga Chubukova, Zilya Vershinina, and Dilara Maslennikova. 2023. "Effects of Pseudomonas sp. OBA 2.4.1 on Growth and Tolerance to Cadmium Stress in Pisum sativum L." BioTech 12, no. 1: 5. https://doi.org/10.3390/biotech12010005
APA StyleKhakimova, L., Chubukova, O., Vershinina, Z., & Maslennikova, D. (2023). Effects of Pseudomonas sp. OBA 2.4.1 on Growth and Tolerance to Cadmium Stress in Pisum sativum L. BioTech, 12(1), 5. https://doi.org/10.3390/biotech12010005