
Effects of Lectins from Alpinia purpurata Inflorescence (ApuL) and Schinus terebinthifolia Leaf (SteLL) on Human Leukemic Cell Lines and Mesenchymal Stem Cells
 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation of Lectins
2.2. Determination of Hemagglutinating Activity (HA) and Protein Concentration
2.3. Leukemic Tumor Lines
2.4. Evaluation of Cytotoxicity to Leukemic Tumor Lines
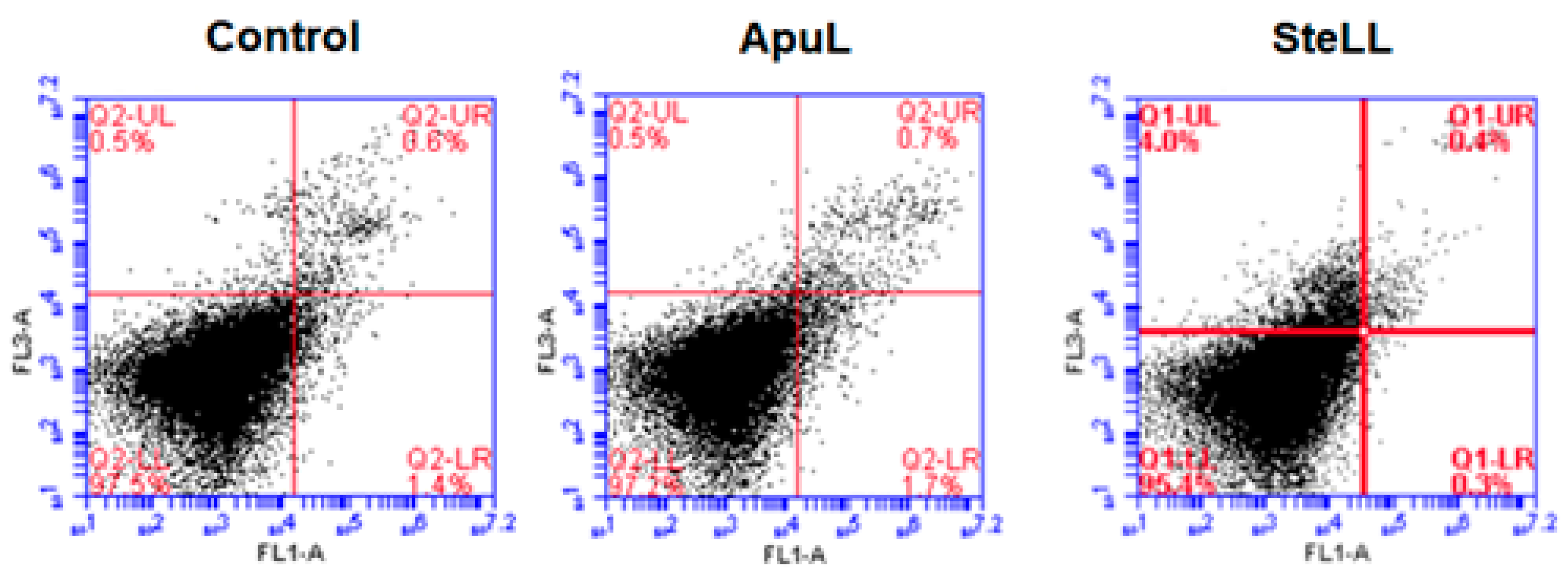
2.5. Evaluation of Cell Death
2.6. Human Umbilical Cords
2.7. Isolation and Cultivation of MSCs
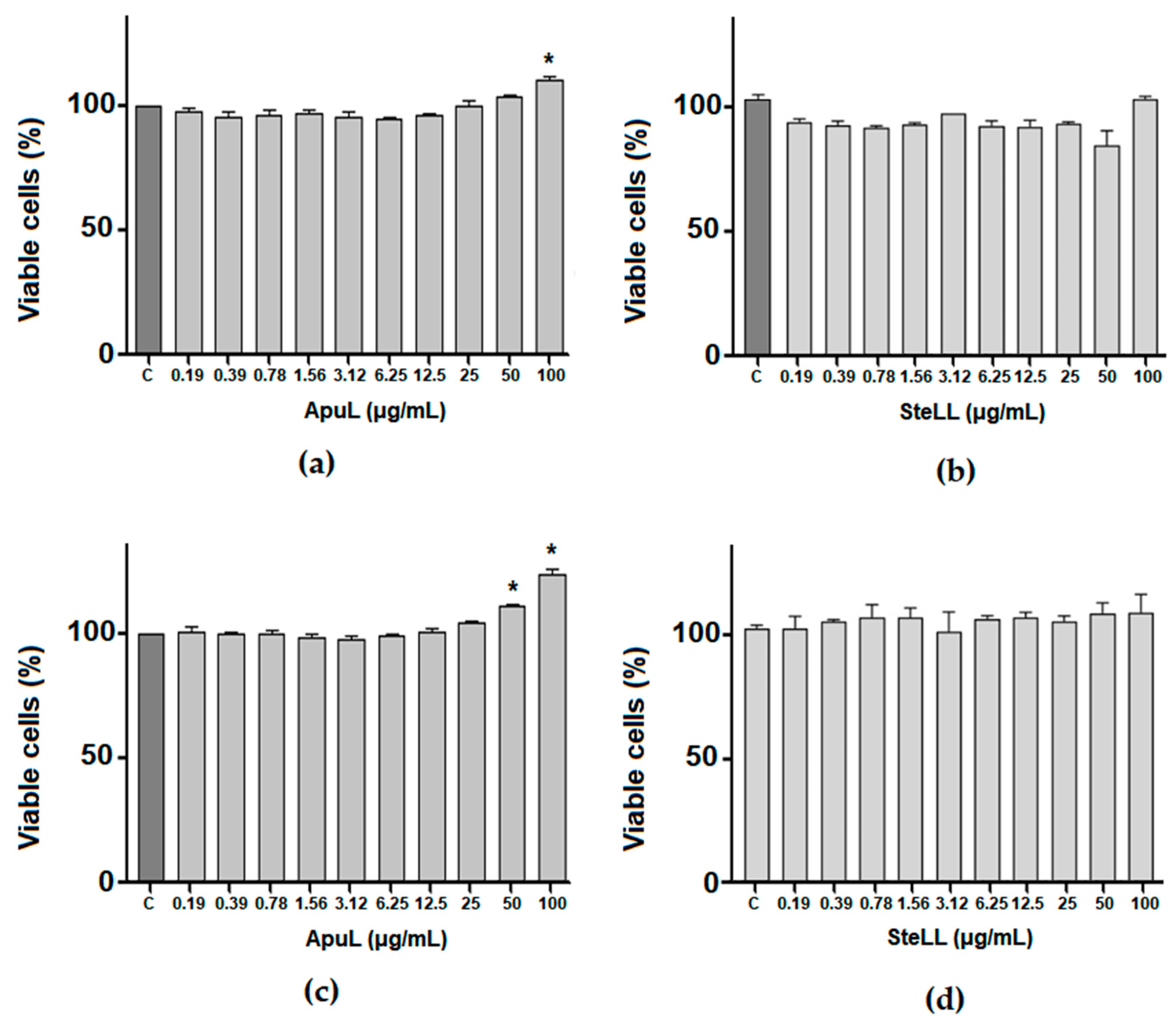
2.8. Evaluation of Cytotoxicity of ApuL and SteLL to MSCs
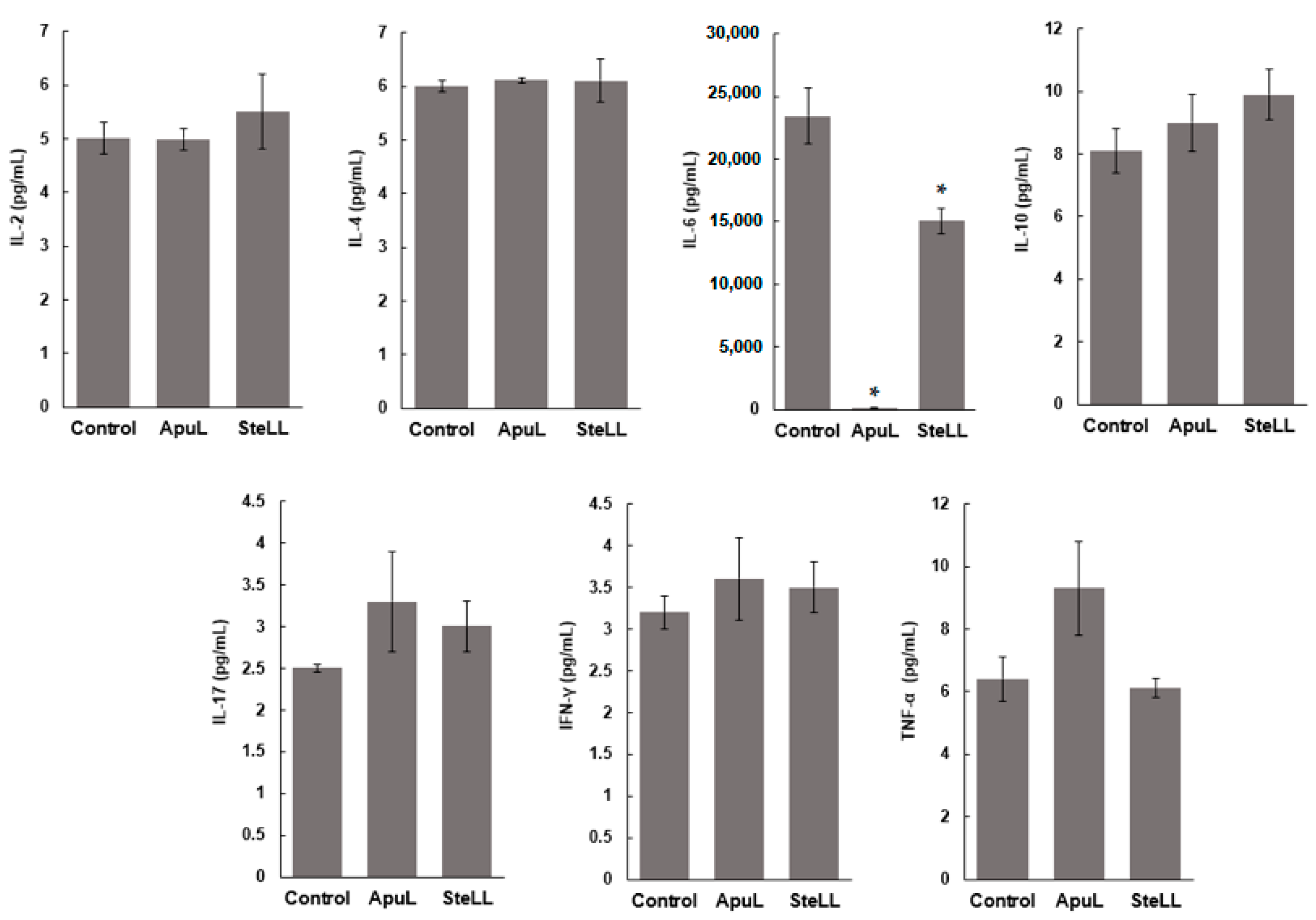
2.9. Cytokine Dosage in MSCs Culture Supernatants
2.10. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Coelho, L.C.B.B.; Silva, P.M.S.; Oliveira, W.F.; Moura, M.C.; Pontual, E.V.; Gomes, F.S.; Paiva, P.M.G.; Napoleão, T.H.; Correia, M.T.S. Lectins as antimicrobial agents. J. Appl. Microbiol. 2018, 125, 1238–1252. [Google Scholar] [CrossRef]
- Patriota, L.L.S.; Brito, J.S.; Ramos, D.B.M.; Procópio, T.F.; Paiva, P.M.G.; Pontual, E.V.; Melo, C.M.L.; Napoleão, T.H. Plant-derived lectins: A review of their status as alternatives to anticancer chemotherapy. In Horizons in Cancer Research; Watanabe, H.S., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2019; pp. 171–205. [Google Scholar]
- Patriota, L.L.S.; Brito, J.S.; Barboza, B.R.; Paiva, P.M.G.; Melo, C.M.L.; Napoleão, T.H. A review on the immunomodulatory effects of plant lectins. In Hemagglutinins: Structures, Functions and Mechanisms; Ng, T.B., Wong, J., Tse, R., Tse, T.F., Chan, H., Eds.; Nova Science Publishers, Inc.: New York, NY, USA, 2019; pp. 53–82. [Google Scholar]
- Brito, J.S.; Ferreira, G.R.S.; Klimczak, E.; Gryshuk, L.; Santos, N.D.L.; Patriota, L.L.S.; Moreira, L.R.; Sores, A.K.A.; Barboza, B.R.; Paiva, P.M.G.; et al. Lectin from inflorescences of ornamental crop Alpinia purpurata acts on immune cells to promote Th1 and Th17 responses, nitric oxide release, and lymphocyte activation. Biomed. Pharmacother. 2017, 94, 865–872. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, G.R.S.; Brito, J.S.; Procópio, T.F.; Santos, N.D.L.; Lima, B.J.R.C.; Coelho, L.C.B.B.; Navarro, D.M.A.F.; Paiva, P.M.G.; Soares, T.S.; Moura, M.C.; et al. Antimicrobial potential of Alpinia purpurata lectin (ApuL): Growth inhibitory action, synergistic effects in combination with antibiotics, and antibiofilm activity. Microb. Pathogen. 2018, 124, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Ramos, D.B.M.; Araújo, M.T.M.; Araújo, T.C.L.; Neto, O.G.S.; Silva, M.G.; Silva, Y.A.; Torres, D.J.L.; Patriota, L.L.S.; Melo, C.M.L.; Lorena, V.M.B.; et al. Evaluation of antitumor activity and toxicity of Schinus terebinthifolia leaf extract and lectin (SteLL) in sarcoma 180-bearing mice. J. Ethnopharmacol. 2019, 233, 148–157. [Google Scholar] [CrossRef]
- Marinho, A.O.; Brito, J.S.; Costa, J.A.; Silva, A.B.; Silva, S.P.; Amorim, L.C.; Correia, M.T.S.; Paiva, P.M.G.; Oliveira, A.M.; Patriota, L.L.S.; et al. Schinus terebinthifolia leaf lectin has central and peripheral antinociceptive action mediated by its carbohydrate-recognition domain and delta-opioid receptors. J. Ethnopharmacol. 2023, 301, 115817. [Google Scholar] [CrossRef]
- Lima, B.R.F.; Patriota, L.L.S.; Marinho, A.O.; Costa, J.A.; Napoleão, T.H.; Rosa, M.M.; Paiva, P.M.G. The lectin from Schinus terebinthifolia leaf (SteLL) reduces immobility of mice on the tail suspension test dependent on the monoaminergic and nitric oxide signaling. Neurosci. Lett. 2023, 801, 137092. [Google Scholar] [CrossRef]
- Lima, B.R.F.; Patriota, L.L.S.; Marinho, A.O.; Costa, J.A.; Napoleão, T.H.; Rosa, M.M.; Paiva, P.M.G. The anxiolytic activity of Schinus terebinthifolia leaf lectin (SteLL) is dependent on monoaminergic signaling although independent of the carbohydrate-binding domain of the lectin. Pharmaceuticals 2022, 15, 136. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.J.C.A.; Araujo, V.F.S.; França, R.P.M.; Silva, E.L.A.; Almeida, W.A.; Procópio, T.F.; Paiva, P.M.G.; Napoleão, T.H.; Costa, E.V.L.; Nogueira, R.A.; et al. Schinus terebinthifolia Raddi leaf lectin is an antiangiogenic agent for Coturnix japonica embryos. An. Acad. Bras. Ciênc. 2022, 94, e20211297. [Google Scholar] [CrossRef]
- Santos, A.J.C.A.; Barros, B.R.S.; Aguiar, L.M.S.; Patriota, L.L.S.; Lima, T.A.; Zingali, R.B.; Paiva, P.M.G.; Napoleão, T.H.; Melo, C.M.L.; Pontual, E.V. Schinus terebinthifolia leaf lectin (SteLL) is an immunomodulatory agent by altering cytokine release by mice splenocytes. 3 Biotech 2020, 10, 144. [Google Scholar] [CrossRef]
- Silva, A.R.; Oliveira, W.F.; Silva, P.M.; Patriota, L.L.S.; Alves, R.R.V.; Oliveira, A.P.S.; Correia, M.T.S.; Paiva, P.M.G.; Vainstein, M.H.; Filho, P.E.C.; et al. Quantum dots conjugated to lectins from Schinus terebinthifolia leaves (SteLL) and Punica granatum sarcotesta (PgTeL) as potential fluorescent nanotools for investigating Cryptococcus neoformans. Int. J. Biol. Macromol. 2021, 192, 232–240. [Google Scholar] [CrossRef]
- World Health Organization. Cancer Fact Sheet; OMS: Genebra, Switzerland, 2022; Available online: https://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 15 March 2023).
- Tessmann, J.W.; Buss, J.; Begnini, K.R.; Berneira, L.M.; Paula, F.R.; Pereira, C.M.P.; Collares, T.; Seixas, F.K. Antitumor potential of 1-thiocarbamoyl-3,5-diaryl-4,5-dihydro-1H- pyrazoles in human bladder cancer cells. Biomed. Pharmacother. 2017, 94, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Shivamadhu, M.C.; Srinivas, B.K.; Jayarama, S.; Chandrashekaraiah, S.A. Anti-cancer and anti-angiogenic effects of partially purified lectin from Praecitrullus fistulosus fruit on in vitro and in vivo model. Biomed. Pharmacother. 2017, 96, 1299–1309. [Google Scholar] [CrossRef] [PubMed]
- Procópio, T.F.; Patriota, L.L.S.; Moura, M.C.; Silva, P.M.; Oliveira, A.P.S.; Carvalho, L.V.N.; Lima, T.A.; Soares, T.; Silva, T.D.; Coelho, L.C.B.B.; et al. CasuL: A new lectin isolated from Calliandra surinamensis leaf pinnulae with cytotoxicity to cancer cells, antimicrobial activity and antibiofilm effect. Int. J. Biol. Macromol. 2017, 98, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Poiroux, G.; Barre, A.; Van Damme, E.; Benoist, H.; Rougé, P. Plant lectins targeting O-glycans at the cell surface as tools for cancer diagnosis, prognosis and therapy. Int. J. Mol. Sci. 2017, 18, 1232. [Google Scholar] [CrossRef] [PubMed]
- Patriota, L.L.S.; Ramos, D.B.M.; Silva, Y.A.; Santos, A.C.L.A.; Araújo, M.T.M.F.; Brito, J.S.; Torres, D.J.L.; Oliveira, A.M.; Silva, D.C.N.; Lorena, V.M.B.; et al. Microgramma vacciniifolia frond lectin (MvFL) exhibits antitumor activity against sarcoma 180 in mice. Phytomed. Plus 2021, 1, 100013. [Google Scholar] [CrossRef]
- Wong, S.W.; Lenzini, S.; Shin, J.W. Perspective: Biophysical regulation of cancerous and normal blood cell lineages in hematopoietic malignancies. APL Bioeng. 2018, 2, 031802. [Google Scholar] [CrossRef]
- Ahmed, A.; Jani, C.; Bhatt, P.; Singh, H.; Jani, R.; Shalhoub, J.; Marshall, D.; Lam, P.; Salciccioli, J. EPR22-104: A comparison of the burden of leukemia amongst various regions of the world, 1990–2019. J. Natl. Compr. Cancer Netw. 2022, 20, EPR22–EPR104. [Google Scholar] [CrossRef]
- Faheina-Martins, G.V.; Silveira, A.L.; Cavalcanti, B.C.; Ramos, M.V.; Moraes, M.O.; Pessoa, C.; Araújo, D.A.M. Antiproliferative effects of lectins from Canavalia ensiformis and Canavalia brasiliensis in human leukemia cell lines. Toxicol. Vitr. 2012, 26, 1161–1169. [Google Scholar] [CrossRef]
- Yau, T.; Dan, X.; Ng, C.C.; Ng, T.B. Lectins with potential for anti-cancer therapy. Molecules 2015, 20, 3791–3810. [Google Scholar] [CrossRef]
- Catanzaro, E.; Calcabrini, C.; Bishayee, A.; Fimognari, C. Antitumor potential of marine and freshwater lectins. Mar. Drugs 2019, 18, 11. [Google Scholar] [CrossRef]
- Konozy, E.H.E.; Osman, M.E.M. Plant lectin: A promising future anti-tumor drug. Biochimie 2022, 202, 136–145. [Google Scholar] [CrossRef]
- Zwetsloot, P.P.; Végh, A.M.; Jansen, S.J.L.; Hout, G.P.V.; Currie, G.L.; Sena, E.S.; Gremmels, H.; Buikema, J.W.; Goumans, M.J.; Macleod, M.R.; et al. Cardiac stem cell treatment in myocardial infarction: A systematic review and meta-analysis of preclinical studies. Circ. Res. 2016, 118, 1223–1232. [Google Scholar] [CrossRef]
- Mochida, T.; Ueno, H.; Yamazoe, N.T.; Hiyoshi, H.; Ito, R.; Matsumoto, H.; Toyoda, T. Insulin-deficient diabetic condition upregulates the insulin-secreting capacity of human induced pluripotent stem cell–derived pancreatic endocrine progenitor cells after implantation in mice. Diabetes 2020, 69, 634–646. [Google Scholar] [CrossRef]
- Xifro, W.A.; Vicino, U.; Martin, M.I.M.; Bortolozzi, A.; Bové, J.; Vila, M.; Cosma, M.P. Functional rescue of dopaminergic neuron loss in Parkinson’s disease mice after transplantation of hematopoietic stem and progenitor cells. EBioMedicine 2016, 8, 83–95. [Google Scholar] [CrossRef]
- Pittenger, M.F.; Martin, B.J. Mesenchymal stem cells and their potential as cardiac therapeutics. Circulat. Res. 2004, 95, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Chao, K.C.; Yang, H.T.; Chen, M.W. Human umbilical cord mesenchymal stem cells suppress breast cancer tumourigenesis through direct cell-cell contact and internalization. J. Cell. Mol. Med. 2012, 16, 1803–1815. [Google Scholar] [CrossRef] [PubMed]
- Ding, D.; Chang, Y.; Shyu, W.; Lin, S. Human umbilical cord mesenchymal stem cells: A new era for stem cell therapy. Cell Transpl. 2015, 24, 339–347. [Google Scholar] [CrossRef]
- Chen, X.; Wang, S.; Cao, W. Mesenchymal stem cell-mediated immunomodulation in cell therapy of neurodegenerative diseases. Cell. Immunol. 2018, 326, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Huang, L.; Li, Y.; Fang, B.; Li, G.; Chen, L.; Xu, L. Mesenchymal stem cells and cancer: Clinical challenges and opportunities. BioMed Res. Int. 2019, 2019, 2820853. [Google Scholar] [CrossRef]
- English, K. Mechanisms of mesenchymal stromal cell immunomodulation. Immunol. Cell Biol. 2013, 91, 19–26. [Google Scholar] [CrossRef]
- Dabrowska, S.; Andrzejewska, A.; Janowski, M.; Lukomska, B. Immunomodulatory and regenerative effects of mesenchymal stem cells and extracellular vesicles: Therapeutic outlook for inflammatory and degenerative diseases. Front. Immunol. 2021, 11, 591065. [Google Scholar] [CrossRef] [PubMed]
- Mikkola, M.; Toivonen, S.; Tamminen, K.; Alfthan, K.; Tuuri, T.; Satomaa, T.; Natunen, J.; Saarinen, J.; Tiittanen, M.; Lampinen, M.; et al. Lectin from Erythrina cristagalli supports undifferentiated growth and differentiation of human pluripotent stem cells. Stem Cells Dev. 2012, 22, 707–712. [Google Scholar] [CrossRef] [PubMed]
- Tateno, H.; Toyota, M.; Saito, S.; Onuma, Y.; Ito, Y.; Hiemori, K.; Asashima, M. Glycome diagnosis of human induced pluripotent stem cells using lectin microarray. J. Biol. Chem. 2011, 286, 20345–20353. [Google Scholar] [CrossRef] [PubMed]
- Hasehira, K.; Hirabayashi, J.; Tateno, H. Structural and quantitative evidence of α2–6-sialylated N-glycans as markers of the differentiation potential of human mesenchymal stem cells. Glycoconj. J. 2017, 34, 797–806. [Google Scholar] [CrossRef]
- Green, A.A.; Hughes, L. Protein fractionation on the basis of solubility in aqueous solution of salts and organic solvents. In Methods in Enzymology; Colowick, S., Kaplan, N., Eds.; Academic Press: New York, NY, USA, 1955; pp. 67–90. [Google Scholar]
- Gomes, F.S.; Procópio, T.F.; Napoleão, T.H.; Coelho, L.C.B.B.; Paiva, P.M.G. Antimicrobial lectin from Schinus terebinthifolius leaf. J. Appl. Microbiol. 2013, 114, 672–679. [Google Scholar] [CrossRef]
- Bing, D.H.; Weyand, J.G.; Stavinsky, A.B. Hemagglutination with aldehyde-fixed erythrocytes for assay of antigens and antibodies. Proc. Soc. Exp. Biol. Med. 1967, 124, 1166–1170. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Präbst, K.; Engelhardt, H.; Ringgeler, S.; Hübner, H. Basic Colorimetric Proliferation Assays: MTT, WST, and Resazurin. In Cell Viability Assays; Springer: New York, NY, USA, 2017; Volume 1601, pp. 1–17. [Google Scholar]
- Miranda, J.M.; Arruda, J.A.A.; Moreno, L.M.M.; Gaião, W.D.C.; Nascimento, S.V.B.; Silva, E.V.S.; Silva, M.B.; Rodrigues, C.G.; Albuquerque, D.S.; Braz, R.; et al. Photobiomodulation therapy in the proliferation and differentiation of human umbilical cord mesenchymal stem cells: An in vitro study. J. Laser Med. Sci. 2020, 11, 469–474. [Google Scholar] [CrossRef]
- Tai, T.; Yamashita, K.; Ito, S.; Kobata, A. Structures of the carbohydrate moiety of ovalbumin glycopeptide III and the difference in specificity of endo-beta-N-acetylglucosaminidases CII and H. J. Biol. Chem. 1977, 252, 6687–6694. [Google Scholar] [CrossRef]
- Singh, T.; Wu, J.H.; Peumans, W.J.; Rougé, P.; Van Damme, E.J.M.; Wu, A.M. Recognition profile of Morus nigra agglutinin (Morniga G) expressed by monomeric ligands, simple clusters and mammalian polyvalent glycotopes. Mol. Immunol. 2007, 44, 451–462. [Google Scholar] [CrossRef]
- Weis, W.I.; Drickamer, K. Structural basis of lectin-carbohydrate recognition. Ann. Rev. Biochem. 1996, 65, 441–473. [Google Scholar] [CrossRef]
- Cao, J.-T.; Chen, Z.-X.; Hao, X.-Y.; Zhang, P.-H.; Zhu, J.-J. Quantum dots-based immunofluorescent microfluidic chip for the analysis of glycan expression at single-cells. Anal. Chem. 2012, 84, 10097–10104. [Google Scholar] [CrossRef]
- Nayak, S.; Zhao, Y.; Mao, Y.; Li, N. System-wide quantitative N-glycoproteomic analysis from K562 cells and mouse liver tissues. J. Proteome Res. 2021, 20, 5196–5202. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhao, Y.; Jiang, L.; Miao, X.; Zhou, H.; Ji, L. Glycomic alterations are associated with multidrug resistance in human leukemia. Int. J. Biochem. Cell Biol. 2012, 44, 1244–1253. [Google Scholar] [CrossRef] [PubMed]
- Lizzi, A.R.; D’alessandro, A.M.; Bozzi, A.; Cinque, B.; Oratore, A.; D’andrea, G. Pattern expression of glycan residues in AZT-treated K562 cells analyzed by lectin cytochemistry. Mol. Cell. Biochem. 2007, 300, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Piller, V.; Piller, F.; Klier, G.; Fukuda, M. O-Glycosylation of leukosialin in K562 cells. Eur. J. Biochem. 1989, 183, 123–135. [Google Scholar] [CrossRef]
- Piller, V.; Piller, F.; Fukuda, M. Biosynthesis of truncated O-glycans in the T cell line Jurkat. Localization of O-glycan initiation. J. Biol. Chem. 1990, 265, 9264–9271. [Google Scholar] [CrossRef] [PubMed]
- Mazalovska, M.; Kouokam, J.C. Transiently expressed mistletoe lectin II in Nicotiana benthamiana demonstrates anticancer activity in vitro. Molecules 2020, 25, 2562. [Google Scholar] [CrossRef]
- Queiroz, A.F.S.; Silva, R.A.; Moura, R.M.; Dreyfuss, J.L.; Gamero, E.J.P.; Souza, A.C.S.; Tersariol, I.L.S.; Santos, E.A.; Nader, H.B.; Justo, G.Z.; et al. Growth inhibitory activity of a novel lectin from Cliona varians against K562 human erythroleukemia cells. Cancer Chemother. Pharmacol. 2009, 63, 1023–1033. [Google Scholar] [CrossRef]
- Huang, L.H.; Yan, Q.J.; Kopparapu, N.K.; Jiang, Z.Q.; Sun, Y. Astragalus membranaceus lectin (AML) induces caspase-dependent apoptosis in human leucemia cells. Cell. Prolif. 2012, 45, 15–21. [Google Scholar] [CrossRef]
- Gao, F.; Chiu, S.; Motan, D.; Zhang, Z.; Chen, L.; Ji, H.-L.; Tse, H.-F.; Fu, Q.-L.; Lian, Q. Mesenchymal stem cells and immunomodulation: Current status and future prospects. Cell Death Dis. 2016, 7, e2062. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Sun, Y.; Shi, J.; Li, C.-F.; Fang, S.-B.; Wang, D.; Deng, X.-Q.; Wen, W.; Fu, Q.-L. Effects of mesenchymal stem cells from human induced pluripotent stem cells on differentiation, maturation, and function of dendritic cells. Stem Cell Res. Ther. 2017, 8, 48. [Google Scholar] [CrossRef] [PubMed]
- Nand, A.; Singh, V.; Wang, P.; Na, J.; Zhu, J. Glycoprotein profiling of stem cells using lectin microarray based on surface plasmon resonance imaging. Anal. Biochem. 2014, 465C, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Dang, K.; Zhang, W.; Jiang, S.; Lin, X.; Qian, A. Application of lectin microarrays for biomarker discovery. Chem. Open 2020, 9, 285–300. [Google Scholar] [CrossRef]
- Lima, I.M.S.F.; Zagmignan, A.; Santos, D.M.; Maia, H.S.; Silva, L.S.; Cutrim, B.S.; Vieira, S.L.; Filho, C.M.B.; Sousa, E.M.; Napoleão, T.H.; et al. Schinus terebinthifolia leaf lectin (SteLL) has anti-infective action and modulates the response of Staphylococcus aureus-infected macrophages. Sci. Rep. 2019, 9, 18159. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.R.; Patriota, L.L.S.; Brito, J.S.; Gaião, W.D.C.; Torres, D.J.L.; Lorena, V.M.B.; Silva, M.B.; Napoleão, T.H. Evaluation of the cytotoxicity of lectin from Microgramma vacciniifolia fronds (MvFL) to human mesenchymal stem cells and leukemia cell lines (K562 and JUKART). Biofarm 2020, 16, 44. [Google Scholar]
- Bonturi, C.R.; Silva, M.C.C.; Motaln, H.; Salu, B.R.; Ferreira, R.S.; Batista, F.P.; Correia, M.T.S.; Paiva, P.M.G.; Turnšek, T.L.; Oliva, M.L.V. A bifunctional molecule with lectin and protease inhibitor activities isolated from Crataeva tapia bark significantly affects cocultures of mesenchymal stem cells and glioblastoma cells. Molecules 2019, 24, 2109. [Google Scholar] [CrossRef]
- Choi, J.H.; Lyu, S.Y.; Lee, H.J.; Jung, J.; Park, W.B.; Kim, G.J. Korean mistletoe lectin regulates self-renewal of placenta-derived mesenchymal stem cells via autophagic mechanisms. Cell Prolif. 2012, 45, 420–429. [Google Scholar] [CrossRef]
- Kim, G.D.; Choi, J.H.; Lim, S.M.; Jun, J.H.; Moon, J.W.; Kim, G.J. Alterations in IL-6/STAT3 signaling by korean mistletoe lectin regulate the self-renewal activity of placenta-derived mesenchymal stem cells. Nutrients 2019, 11, 2604. [Google Scholar] [CrossRef]
- Bouffi, C.; Bony, C.; Courties, G.; Jorgensen, C.; Noël, D. IL-6-Dependent PGE2 secretion by mesenchymal stem cells inhibits local inflammation in experimental arthritis. PLoS ONE 2010, 5, e14247. [Google Scholar] [CrossRef]
- Liu, C.; Hwang, S. Cytokine interactions in mesenchymal stem cells from cord blood. Cytokine 2005, 32, 270–279. [Google Scholar] [CrossRef] [PubMed]
- Klassen, H.J.; Imfeld, K.L.; Kirov, I.I.; Tai, L.; Gage, F.H.; Young, M.J.; Berman, M.A. Expression of cytokines by multipotent neural progenitor cells. Cytokine 2003, 22, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Pricola, K.L.; Kuhn, N.Z.; Haleem-Smith, H.; Song, Y.; Tuan, R.S. Interleukin-6 maintains bone marrow-derived mesenchymal stem cell stemness by an ERK1/2-dependent mechanism. J. Cell. Biochem. 2009, 108, 577–588. [Google Scholar] [CrossRef] [PubMed]
- Kondo, M.; Yamaoka, K.; Sakata, K.; Sonomoto, K.; Lin, L.; Nakano, K.; Tanaka, Y. Contribution of the interleukin-6/STAT-3 signaling pathway to chondrogenic differentiation of human mesenchymal stem cells. Arthritis Rheumatol. 2015, 67, 1250–1260. [Google Scholar] [CrossRef]
- Xie, Z.; Tang, S.; Ye, G.; Wang, P.; Li, J.; Liu, W.; Li, M.; Wang, S.; Wu, X.; Cen, S.; et al. Interleukin-6/interleukin-6 receptor complex promotes osteogenic differentiation of bone marrow-derived mesenchymal stem cells. Stem Cell Res. Ther. 2018, 9, 13. [Google Scholar] [CrossRef]
- He, M.; Shi, X.; Yang, M.; Yang, T.; Li, T.; Chen, J. Mesenchymal stem cells-derived IL-6 activates AMPK/mTOR signaling to inhibit the proliferation of reactive astrocytes induced by hypoxic-ischemic brain damage. Exp. Neurol. 2018, 311, 15–32. [Google Scholar] [CrossRef] [PubMed]
- Rasmusson, I. Immune modulation by mesenchymal stem cells. Exp. Cell Res. 2006, 312, 2169–2179. [Google Scholar] [CrossRef]
- Engela, A.U.; Baan, C.C.; Dor, F.J.M.F.; Weimar, W.; Hoogduijn, M.J. On the interactions between mesenchymal stem cells and regulatory T cells for immunomodulation in transplantation. Front. Immunol. 2012, 3, 126. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, X.; Cao, W.; Shi, Y. Plasticity of mesenchymal stem cells in immunomodulation: Pathological and therapeutic implications. Nat. Immunol. 2014, 15, 1009–1016. [Google Scholar] [CrossRef]
- Raffaghello, L.; Bianchi, G.; Bartolotto, M.; Montecucco, F.; Busca, A.; Dallegri, F.; Ottonello, L.; Pistoia, V. Human mesenchymal stem cells inhibit neutrophil apoptosis: A model for neutrophil preservation in the bone marrow niche. Stem Cells 2008, 26, 151–162. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Santana Brito, J.; de Oliveira Marinho, A.; de Siqueira Patriota, L.L.; Gaião, W.D.C.; Torres, D.J.L.; Paiva, P.M.G.; de Lorena, V.M.B.; Rodrigues, C.G.; da Silva, M.B.; Napoleão, T.H. Effects of Lectins from Alpinia purpurata Inflorescence (ApuL) and Schinus terebinthifolia Leaf (SteLL) on Human Leukemic Cell Lines and Mesenchymal Stem Cells. Macromol 2023, 3, 290-302. https://doi.org/10.3390/macromol3020018
de Santana Brito J, de Oliveira Marinho A, de Siqueira Patriota LL, Gaião WDC, Torres DJL, Paiva PMG, de Lorena VMB, Rodrigues CG, da Silva MB, Napoleão TH. Effects of Lectins from Alpinia purpurata Inflorescence (ApuL) and Schinus terebinthifolia Leaf (SteLL) on Human Leukemic Cell Lines and Mesenchymal Stem Cells. Macromol. 2023; 3(2):290-302. https://doi.org/10.3390/macromol3020018
Chicago/Turabian Stylede Santana Brito, Jéssica, Amanda de Oliveira Marinho, Leydianne Leite de Siqueira Patriota, Wyndly Daniel Cardoso Gaião, Diego José Lira Torres, Patrícia Maria Guedes Paiva, Virgínia Maria Barros de Lorena, Cláudio Gabriel Rodrigues, Márcia Bezerra da Silva, and Thiago Henrique Napoleão. 2023. "Effects of Lectins from Alpinia purpurata Inflorescence (ApuL) and Schinus terebinthifolia Leaf (SteLL) on Human Leukemic Cell Lines and Mesenchymal Stem Cells" Macromol 3, no. 2: 290-302. https://doi.org/10.3390/macromol3020018
APA Stylede Santana Brito, J., de Oliveira Marinho, A., de Siqueira Patriota, L. L., Gaião, W. D. C., Torres, D. J. L., Paiva, P. M. G., de Lorena, V. M. B., Rodrigues, C. G., da Silva, M. B., & Napoleão, T. H. (2023). Effects of Lectins from Alpinia purpurata Inflorescence (ApuL) and Schinus terebinthifolia Leaf (SteLL) on Human Leukemic Cell Lines and Mesenchymal Stem Cells. Macromol, 3(2), 290-302. https://doi.org/10.3390/macromol3020018