
Bird Communities and the Rehabilitation of Al Karaana Lagoons in Qatar
 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
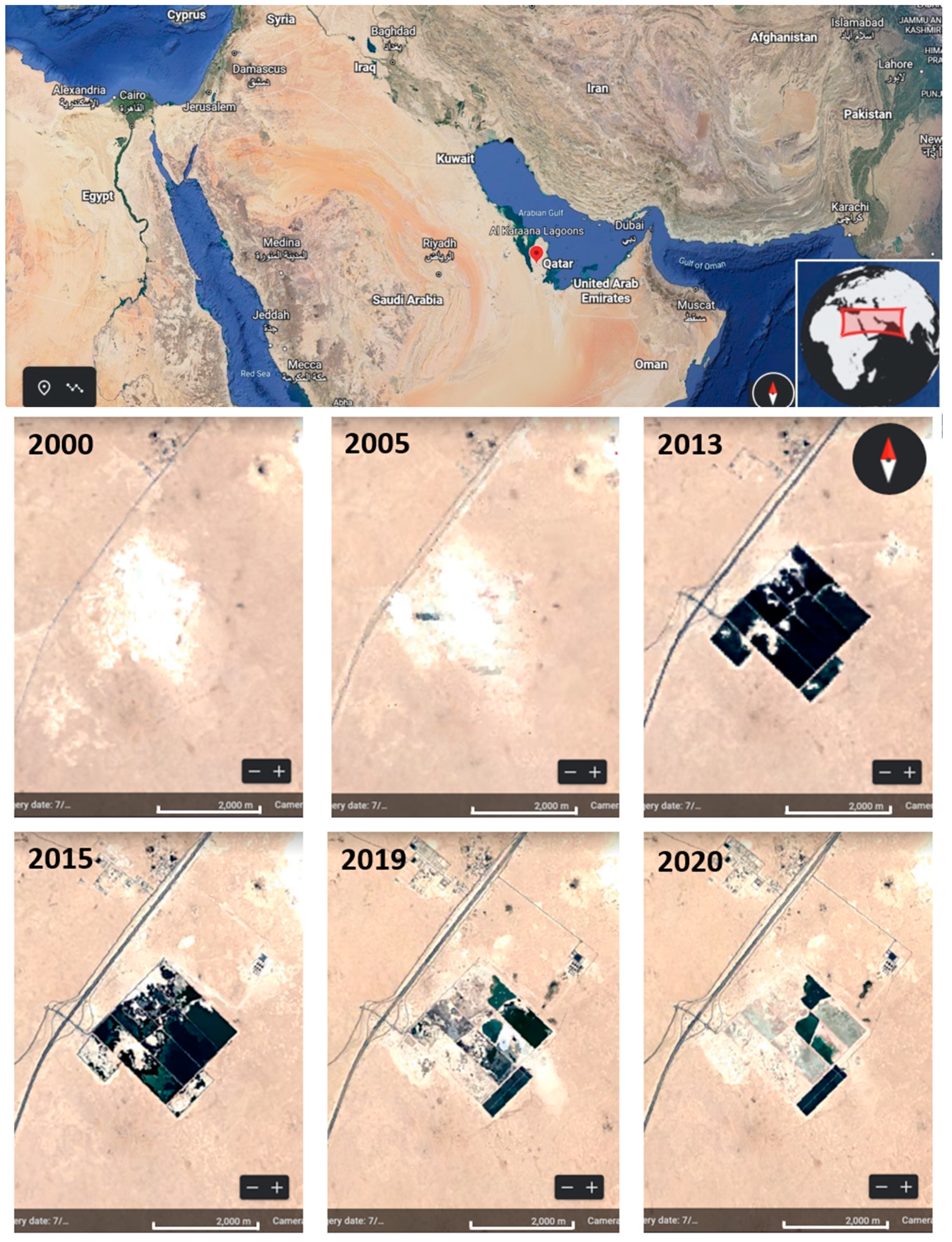
2.1. Study Site Description and History
2.2. Survey Protocols
2.3. Pre-Remediation Surveys
2.4. Statistical Analysis
2.4.1. Descriptive Statistics
2.4.2. Statistical Comparisons
3. Results
3.1. Comparison before and after Rehabilitation
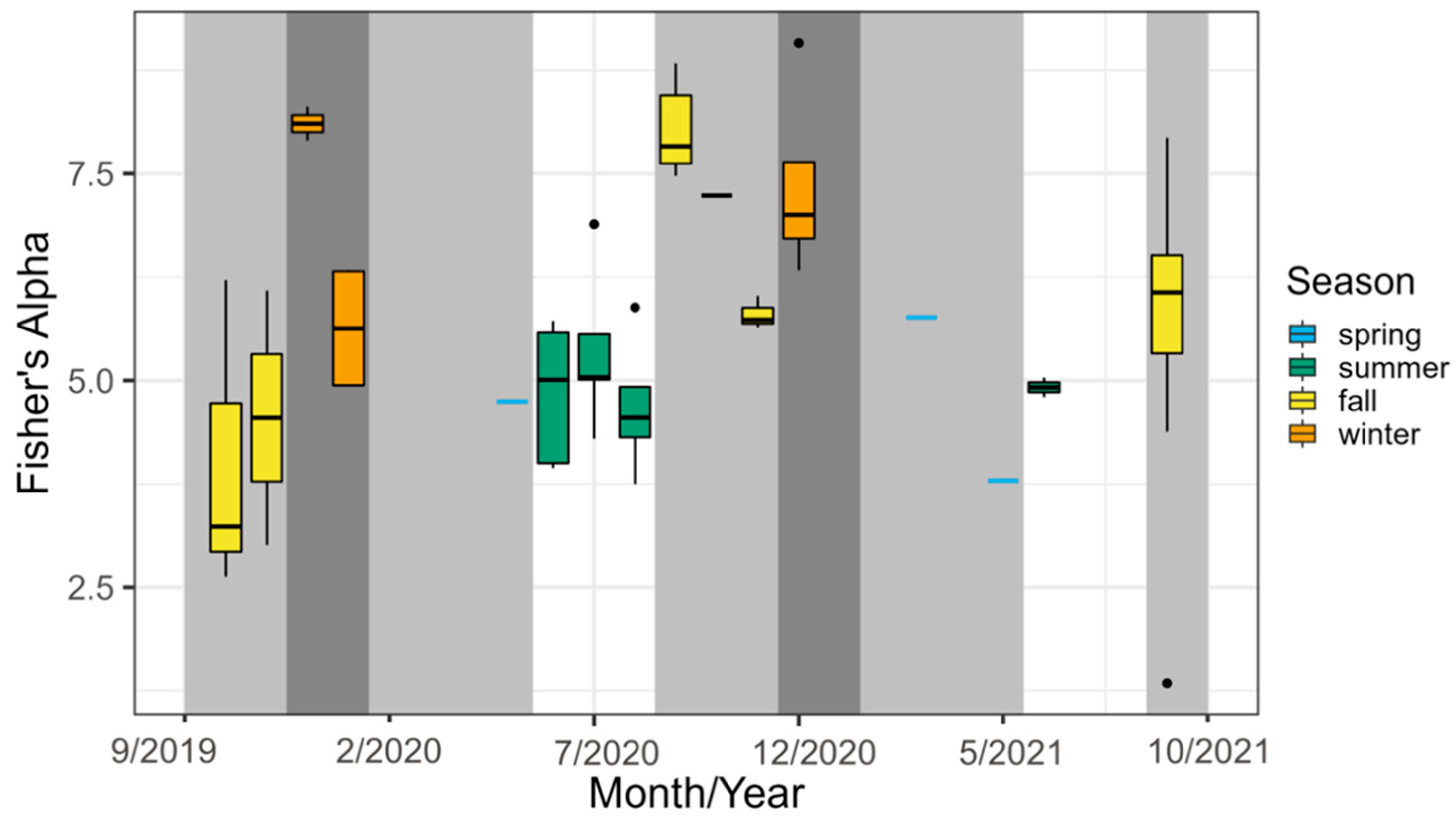
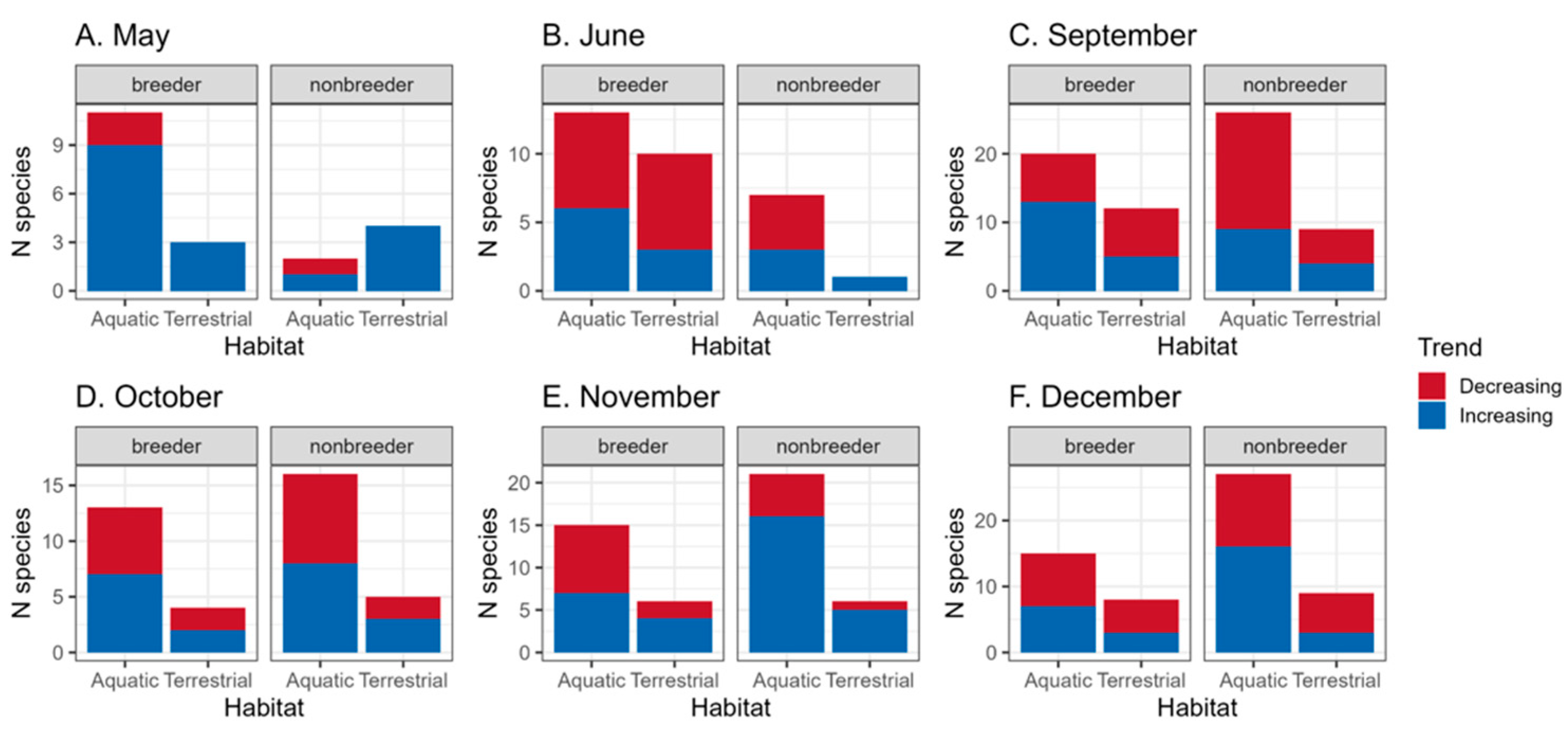
3.2. Impact of Migrants and Over-Wintering Birds on the Bird Community
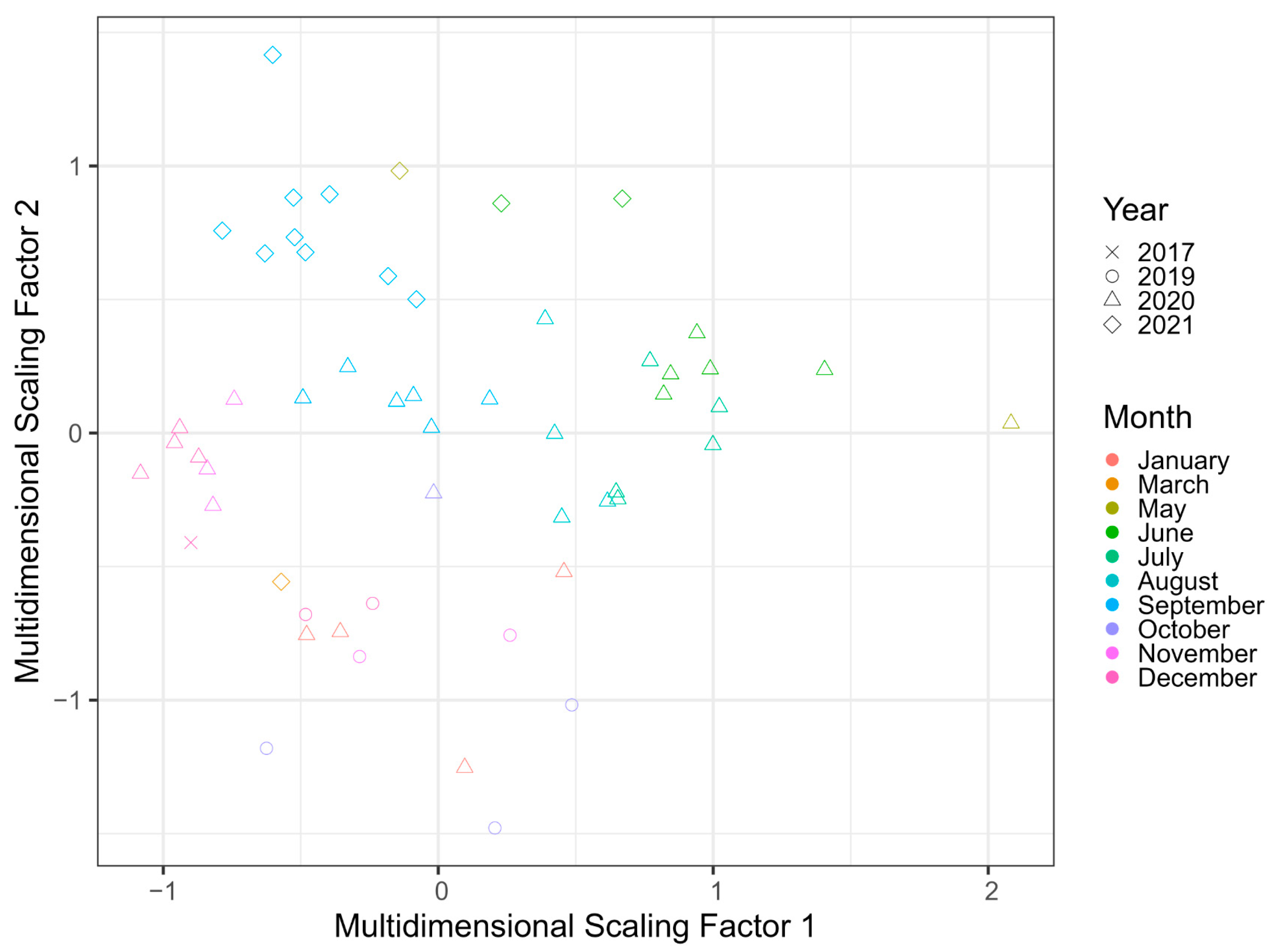
3.3. Change in the Site following Rehabilitation: Whole Site
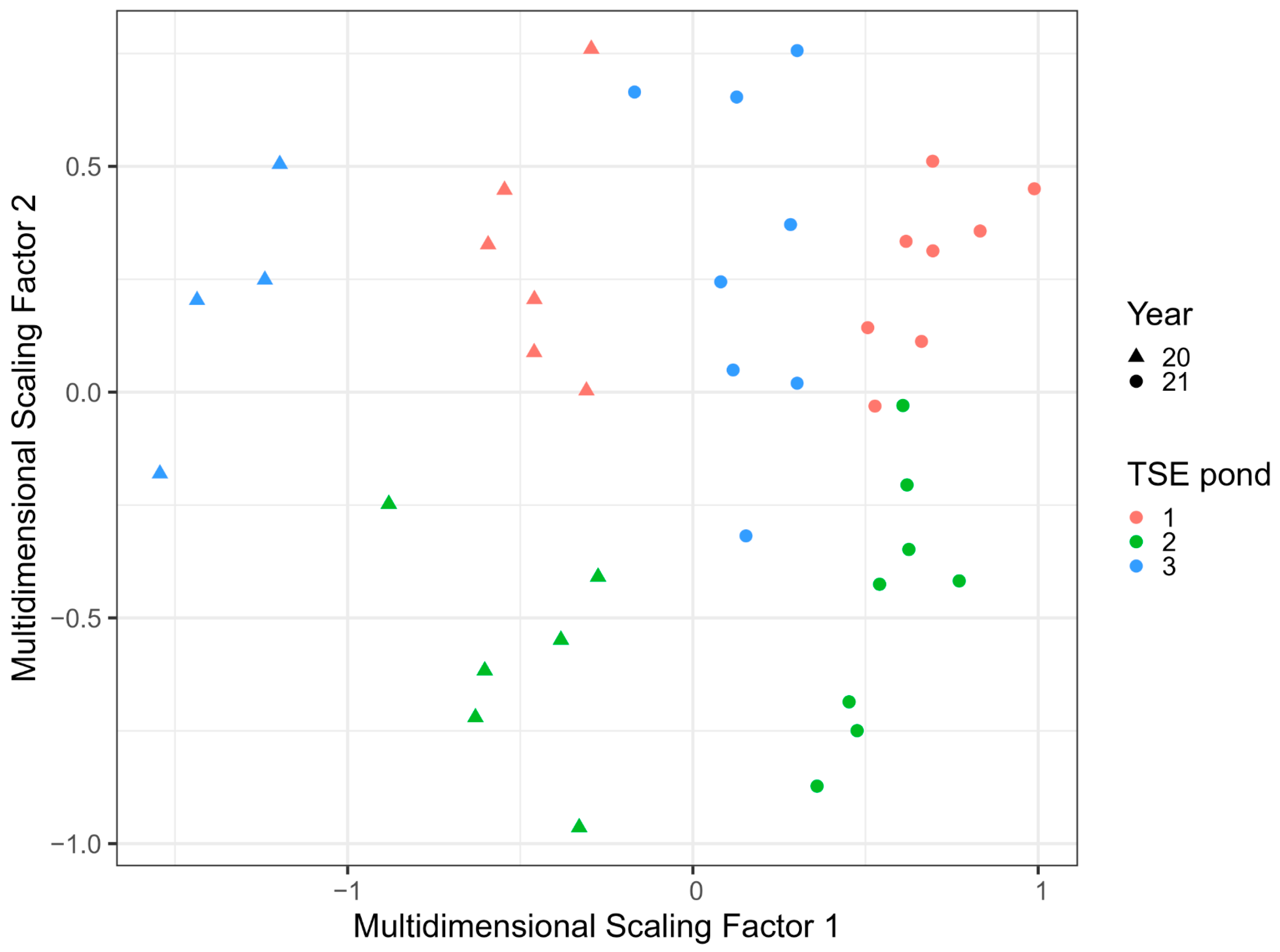
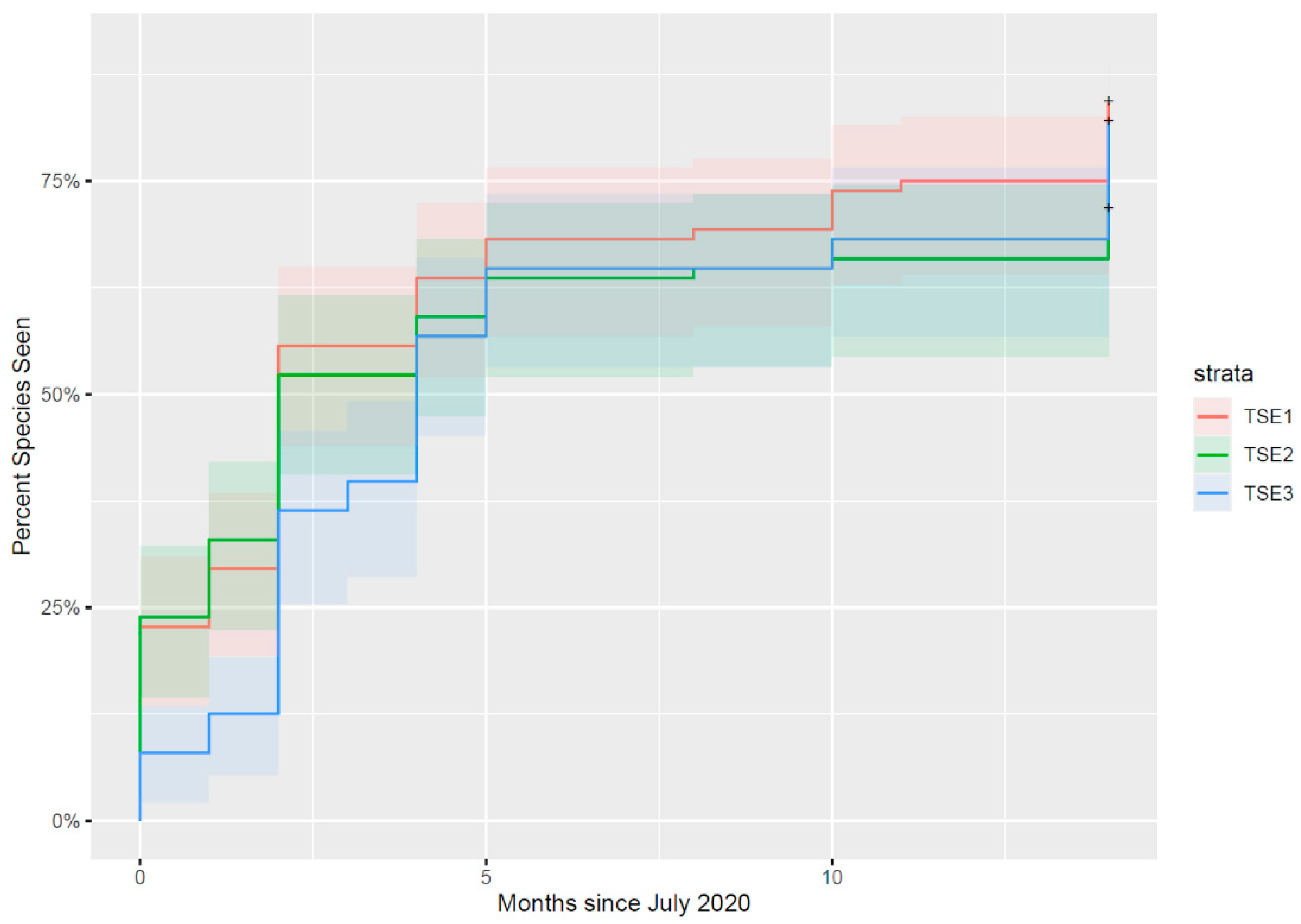
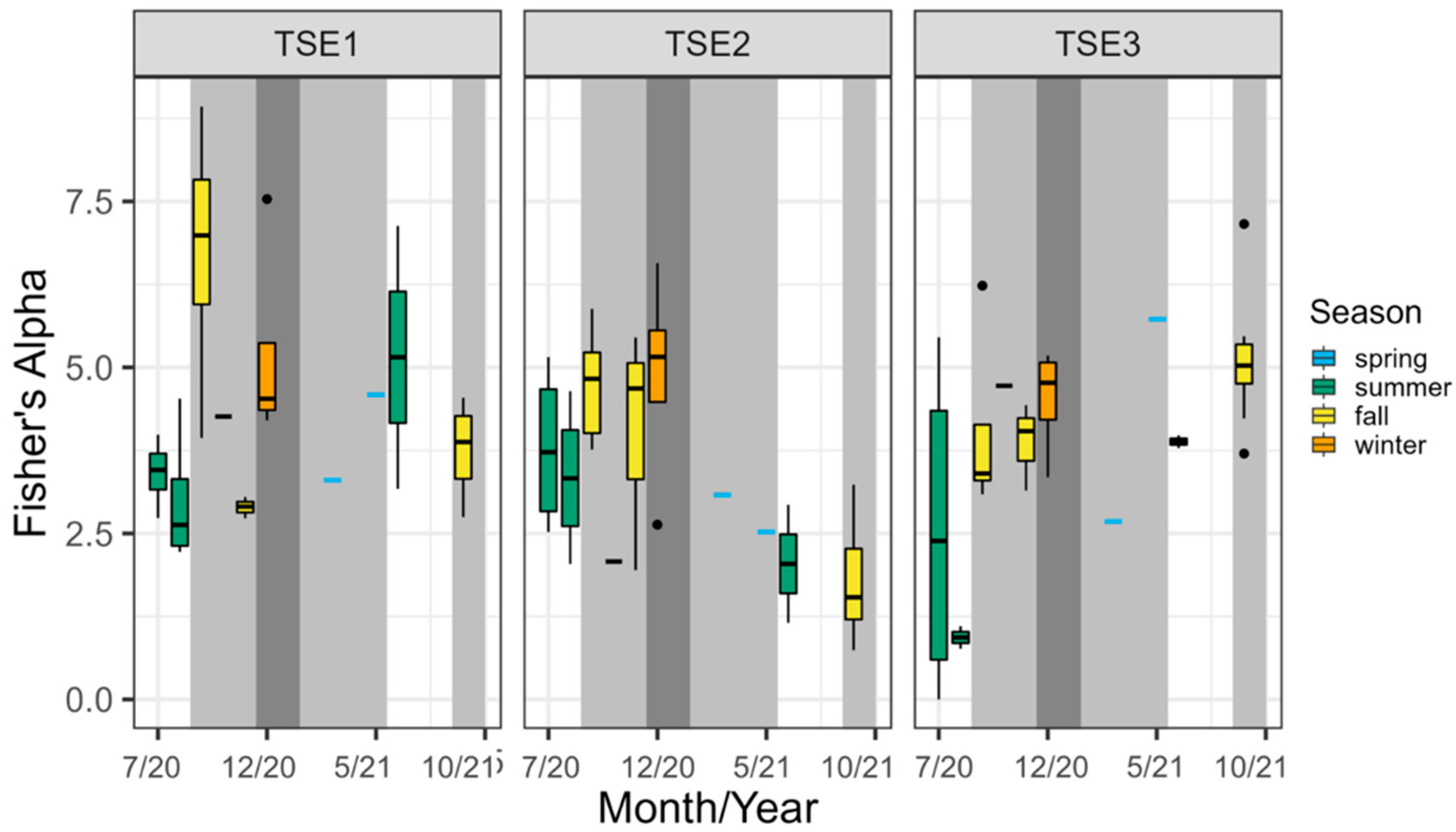
3.4. Change in the Site following Rehabilitation: Comparing TSE Ponds
4. Discussion
4.1. Changes in the Bird Community following Rehabilitation
4.2. Comparison of Different TSE Pond Designs
4.3. Importance for Migratory Species
4.4. Management and Future Prospects
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dar, S.A.; Bhat, S.U.; Rashid, I.; Dar, S.A. Current status of wetlands in Srinagar city: Threats, management strategies, and future perspectives. Front. Environ. Sci. 2020, 7, 199. [Google Scholar] [CrossRef]
- Greenway, M.; Simpson, J.S. Artificial wetlands for wastewater treatment, water reuse and wildlife in Queensland, Australia. Water Sci. Technol. 1996, 33, 221–229. [Google Scholar] [CrossRef]
- Hsu, C.; Hsieh, H.; Yang, L.; Wu, S.; Chang, J.; Hsiao, S.; Su, H.; Yeh, C.; Ho, Y.; Lin, H. Biodiversity of constructed wetlands for wastewater treatment. Ecol. Eng. 2011, 37, 1533–1545. [Google Scholar] [CrossRef]
- Moreno-Mateos, D.; Power, M.E.; Comin, F.A.; Yockten, R. Structural and functional loss in restored wetland ecosystems. PLoS Biol. 2012, 10, e1001247. [Google Scholar] [CrossRef]
- Moreno-Mateos, D.; Meli, P.; Vara-Rodriquez, M.I.; Aronson, J. Ecosystem response to interventions: Lessons from restored and created wetland ecosystems. J. Appl. Ecol. 2015, 52, 1528–1537. [Google Scholar] [CrossRef]
- Rajpar, M.; Zakaria, M. Assessing an artificial wetland in Putrajaya, Malaysia, as an alternate habitat for waterbirds. Waterbirds 2013, 36, 482–493. [Google Scholar] [CrossRef]
- Fazaa, N.; Dunn, J.; Whittingham, M. Distributions and community composition of birds in Iraq’s central marsh. Int. J. Biodivers. 2017, 2017, 4198690. [Google Scholar] [CrossRef]
- Giosa, E.; Mammides, C.; Zotos, S. The importance of artificial wetlands for birds: A case study from Cyprus. PLoS ONE 2018, 13, e0197286. [Google Scholar]
- Kačergytė, I.; Pärt, T.; Berg, Å.; Debora, A.; Michal, Ż.; Jonas, K. Quantifying effects of wetland restorations on bird communities in agricultural landscapes. Biol. Conserv. 2022, 273, 109676. [Google Scholar] [CrossRef]
- Davidson, N. How much wetland has the world lost? Long-term and recent trends in global wetland area. Mar. Freshw. Res. 2014, 65, 934–941. [Google Scholar] [CrossRef]
- Hu, S.; Niu, Z.; Chen, Y.; Li, L.; Zhang, H. Global wetlands: Potential distribution, wetland loss, and status. Sci. Total Environ. 2017, 586, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Dawoud, M.A.; Sallam, O.M.; Abdelfattah, M.A. Groundwater economics in arid regions: Abu Dhabi Emirate case study. Desalination Water Treat. 2020, 176, 84–93. [Google Scholar] [CrossRef]
- Galbraith, C.A.; Jones, T.; Kirby, J.; Mundkur, T. A Review of Migratory Bird Flyways and Priorities for Management; CMS Technical Series Publication No. 27; UNEP/CMS Secretariat: Bonn, Germany, 2014. [Google Scholar]
- Qatar Awards Contract for Lagoon Clean-Up. Construction Index. The Construction Index. 26 March 2015. Available online: https://www.theconstructionindex.co.uk/news/view/qatar-awards-contract-for-lagoon-clean-up (accessed on 18 July 2022).
- Yu, R.; Chen, G.Z.; Wong, Y.S.; Tam, N.F.Y.; Lan, C.Y. Benthic macrofauna of the mangrove swamp treated with municipal wastewater. Hydrobiologia 1997, 347, 127–137. [Google Scholar] [CrossRef]
- Cannici, S.; Bartolini, F.; Dahdouh-Guebas, F.; Fratini, S.; Litulo, C.; Macia, A.; Mrabu, E.J.; Penha-Lopes, G.; Paula, J. Effects of urban wastewater on crab and mollusc assemblages in equatorial and subtropical mangroves of East Africa. Estuar. Coast. Shelf Sci. 2009, 84, 305–317. [Google Scholar] [CrossRef]
- Penha-Lopes, G.; Torres, P.; Narciso, L.; Cannicci, S.; Paula, J. Comparison of fecundity, embryo loss and fatty acid composition of mangrove crab species in sewage contaminated and pristine mangrove habitats in Mozambique. J. Exp. Mar. Biol. Ecol. 2009, 381, 25–32. [Google Scholar] [CrossRef]
- VanRees-Siewert, K.L.; Dinsmore, J.J. Influence of wetland age on bird use of restored wetlands in Iowa. Wetlands 1996, 16, 577–582. [Google Scholar] [CrossRef]
- Campbell, O.; Moran, N. Phenology of spring landbird migration through Abu Dhabi island, United Arab Emirates, 2007–2014. Sandgrouse 2016, 38, 38–70. [Google Scholar]
- Campbell, O.; Smiles, M. A phenological analysis of spring and autumn bird migration through the United Arab Emirates. Sandgrouse 2021, 43, 43–89. [Google Scholar]
- Qatar Meteorology Department (n.d.). QWeather Meteorology. Available online: https://qweather.gov.qa/CAA/Index.aspx (accessed on 20 September 2022).
- Egis International; (Doha, Qatar). Environmental Impact Assessment—Al Karaana Sewage Lagoons (Rev1). 2016; (Unpublished work). [Google Scholar]
- KEO Internation Consultants; (Doha, Qatar). Reclamation and Rehabilitation of the Al Karaana Lagoons. Ecological Study: Technical Report (Rev. 2). 2018; (Unpublished work). [Google Scholar]
- Porter, R.; Aspinall, S. Birds of the Middle East, 2nd ed.; Christopher Helm Publishers Ltd.: London, UK, 2010. [Google Scholar]
- Erikson, H.; Erikson, J.; Gillespie, F. Common Birds of Qatar; Frances Gillespie: Paris, France, 2011. [Google Scholar]
- Schulze, A.; Dingler, K. The Bird Songs of Europe, North Africa and the Middle East—[CD]; Ample, Inc.: San Francisco, CA, USA, 2007. [Google Scholar]
- Mikrasya. Counter (Version 2019.3) [Mobile App] App Store. 2012. Available online: https://apps.apple.com/app/id1144352844 (accessed on 1 February 2020).
- WKC Group; (Doha, Qatar). Al Karaana WWTP Ecology Baseline Survey. 2015; (Unpublished work). [Google Scholar]
- Whittaker, R.H. Evolution and measurement of species diversity. Taxon 1972, 21, 213–251. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Oksanen, J.F.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, M.P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-7. 2020. Available online: https://CRAN.R-project.org/package=vegan (accessed on 13 March 2022).
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009; Volume 574. [Google Scholar]
- Hadfield, J.D. MCMC methods for multi-response generalized linear mixed models: The MCMCglmm R package. J. Stat. Softw. 2010, 33, 1–22. [Google Scholar] [CrossRef]
- De Villemereuil, P.; Nakagawa, S. General quantitative genetic methods for comparative biology. In Practical Materials for the Book of Modern Phylogenetic Comparative Methods; Garamszegi, L.Z., Ed.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 287–303. [Google Scholar]
- Therneau, T.M.; Grambsch, P.M. Modelling Survival Data: Extending the Cox Model; Springer: Berlin/Heidelberg, Germany, 2000. [Google Scholar]
- Therneau, T.M. Survival Analysis [R Package Survival Version 3.3-1]. The Comprehensive R Archive Network. 3 March 2022. Available online: https://cran.r-project.org/web/packages/survival/index.html (accessed on 13 March 2022).
- Brehm, G.; Süssenbach, D.; Fiedler, K. Unique elevational diversity patterns of geometrid moths in an Andean montane rainforest. Ecography 2003, 26, 456–466. [Google Scholar] [CrossRef]
- Eeva, T.; Belskii, E.; Gilyazov, A.S.; Kozlov, M.V. Pollution impacts on bird population density and species diversity at four non-ferrous smelter sites. Biol. Conserv. 2012, 150, 33–41. [Google Scholar] [CrossRef]
- Zhang, W.; Saint, C.; Weinstein, P.; Slaney, D. Reclaimed water systems: Biodiversity friend or foe? In Water Challenges and Solutions on a Global Scale; American Chemical Society: Washington, DC, USA, 2015; pp. 355–374. [Google Scholar]
- Stanton, R.; Morrissey, C.; Clark, R. Analysis of trends and agricultural drivers of farmland bird declines in North America: A review. Agric. Ecosyst. Environ. 2018, 254, 244–254. [Google Scholar] [CrossRef]
- Ibarra-Meza, I.; Barrera-Huertas, H.A.; López-López, E. Atmospheric pollution assessment using polar graphs and its relationship with the bird community from the Sierra de Guadalupe, Mexico (initial data). Environ. Sci. Pollut. Res. 2021, 29, 61711–61728. [Google Scholar] [CrossRef]
- Coveney, M.F.; Stites, D.L.; Lowe, E.F.; Battoe, L.E.; Conrow, R. Nutrient removal from eutrophic lake water by wetland filtration. Ecol. Eng. 2002, 19, 141–159. [Google Scholar] [CrossRef]
- Avornyo, S.Y. Water Quality Assessment of Some Coastal Lagoons in Ghana; Lambert Academic Publishing: Sunnyvale, CA, USA, 2017. [Google Scholar]
- Bilal, S.; Rais, M.; Anwar, M.; Hussain, I.; Sharif, M.; Kabeer, B. Habitat association of Little Grebe (Tachybaptus ruficollis) at Kallar Kahar Lake, Pakistan. J. King Saud Univ.-Sci. 2013, 25, 267–270. [Google Scholar] [CrossRef]
- Furness, R.; Greenwood, J. Birds as Monitors of Environmental Change; Chapman & Hall: London, UK, 1993. [Google Scholar]
- Pretty, J.N.; Mason, C.F.; Nedwell, D.B.; Hine, R.E.; Leaf, S.; Dils, R. Environmental costs of freshwater eutrophication in England and Wales. Environ. Sci. Technol. 2002, 37, 201–208. [Google Scholar] [CrossRef]
- Hoyer, M.V.; Canfield, D.E. Bird abundance and species richness on Florida lakes: Influence of trophic status, lake morphology, and aquatic macrophytes. In Aquatic Birds in the Trophic Web of Lakes; Springer: Dordrecht, The Netherlands, 1994; pp. 107–119. [Google Scholar]
- Vohla, C.; Kõiv, M.; Bavor, H.J.; Chazarenc, F.; Mander, Ü. Filter materials for phosphorus removal from wastewater in treatment wetlands—A review. Ecol. Eng. 2011, 37, 70–89. [Google Scholar] [CrossRef]
- Flynn, K.J.; Clark, D.R.; Mitra, A.; Fabian, H.; Hansen, P.J.; Glibert, P.M.; Wheeler, G.L.; Stoecker, D.K.; Blackford, J.C.; Brownlee, C. Ocean acidification with (de)eutrophication will alter future phytoplankton growth and succession. Proc. R. Soc. B Biol. Sci. 2015, 282, 20142604. [Google Scholar] [CrossRef]
- Zhang, H.; Tian, Y.; Cui, S.; Zhang, L.; Zhong, X.; Xiong, Y. Influence of macrophytes on phosphorus fractionation in surface sediments in a constructed wetland: Insight from sediment compositions. Ecol. Eng. 2016, 97, 400–409. [Google Scholar] [CrossRef]
- Ma, Z.; Cai, Y.; Li, B.; Chen, J. Managing wetland habitats for waterbirds: An international perspective. Wetlands 2010, 30, 15–27. [Google Scholar] [CrossRef]
- Newton, S.F.; Newton, A.V. The effect of rainfall and habitat on abundance and diversity of birds in a fenced protected area in the central Saudi Arabian desert. J. Arid Environ. 1997, 35, 715–735. [Google Scholar] [CrossRef]
- Zuberogoitia, I.; Martínez, J.E.; González-Oreja, J.A.; de Buitrago, C.G.; Belamendia, G.; Zabala, J.; Laso, M.; Pagaldai, N.; Jiménez-Franco, M.V. Maximizing detection probability for effective large-scale nocturnal bird monitoring. Divers. Distrib. 2020, 26, 1034–1050. [Google Scholar] [CrossRef]
- Campbell, O.; Smiles, M. Notable breeding records from a recently established anthropogenic, agricultural, site in the United Arab Emirates. Sandgrouse 2019, 41, 18–31. [Google Scholar]
- Elhassan, E.; Sládeček, M.; Badaam, S.; Brynychová, K.; Chajma, P.; Firlová, V.; Janatová, V.; Kubelka, L.; Pešková, E.; Vozabulová, A.; et al. An artificial lakes system intended for human recreation supports a vital breeding population of Red-wattled Lapwing in the Arabian Desert. Avian Conserv. Ecol. 2021, 16, 20. [Google Scholar] [CrossRef]
- Nyman, J.A.; Chabreck, R.H. Fire in coastal marshes: History and recent concerns. In Fire in Wetlands: A Management Perspective. Proceedings of the Tall Timbers Fire Ecology Conference; Tall Timbers Research Station: Tallahassee, FL, USA, 1995; Volume 19, pp. 134–141. [Google Scholar]
- Eid, E.M.; Shaltout, K.H.; Al-Sodany, Y.M.; Haroun, S.A.; Galal, T.M.; Ayed, H.; Khedher, K.M.; Jensen, K. Seasonal potential of Phragmites australis in nutrient removal to eliminate the eutrophication in Lake Burullus, Egypt. J. Freshw. Ecol. 2020, 35, 135–155. [Google Scholar] [CrossRef]
- Zwarts, L.; Felemban, H.; Price, A.R. Wader counts along the Saudi Arabian coast suggests the Gulf harbours millions of waders. Wader Study Group Bull. 1991, 63, 25–32. [Google Scholar]
- Zogaris, S.; Kallimanis, A. Coastal zone habitat-use by birds in Qatar: Insights from a rapid assessment method during spring migration. Trop. Conserv. Sci. 2016, 9, 658–676. [Google Scholar] [CrossRef]
- Hoffman, T.; Carra, L.G.; Öhagen, P.; Fransson, T.; Barboutis, C.; Piacentini, D.; Figuerola, J.; Kiat, Y.; Onrubia, A.; Jaenson, T.G.; et al. Association between guilds of birds in the African-Western Palaearctic region and the tick species Hyalomma rufipes, one of the main vectors of Crimean-Congo hemorrhagic fever virus. One Health 2021, 13, 100349. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time Period | Whittaker’s Alpha Diversity | |
|---|---|---|
| Whittaker’s Alpha Diversity | September (Year) | December (Year) |
| Before remediation | 31 (2015) | 18 (2017) |
| After remediation | 53 (2020), 50 (2021) | 32 (2019), 31 (2020) |
| Months Before and After Remediation, Respectively | Beta Diversity | Gamma Diversity |
|---|---|---|
| September 2015 vs. September 2020 | 34 | 69 |
| September 2015 vs. September 2021 | 39 | 60 |
| September 2020 vs. September 2021 | 39 | 71 |
| December 2017 vs. December 2019 | 24 | 37 |
| December 2017 vs. December 2020 | 25 | 37 |
| December 2019 vs. December 2020 | 28 | 64 |
| Total No. of Avian Individuals | Most Abundant Species | Number of Individuals from the Most Abundant Species | |
|---|---|---|---|
| Before remediation (3 December 2017) | 863 | Little Stint (Calidris minuta) | 300 |
| After remediation (5 December 2020) | 1011 | Little Stint (Calidris minuta) | 116 |
| Month | Years Compared | N species Increasing | N species Decreasing |
|---|---|---|---|
| May | 2020–2021 | 17 | 3 |
| June | 2020–2021 | 13 | 18 |
| September | 2020–2021 | 31 | 36 |
| October | 2019–2020 | 20 | 18 |
| November | 2019–2020 | 32 | 16 |
| December | 2019–2020 | 29 | 30 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Draidia, A.; Tareen, M.; Bayraktar, N.; Cramer, E.R.A.; Chen, K.-C. Bird Communities and the Rehabilitation of Al Karaana Lagoons in Qatar. Birds 2022, 3, 320-340. https://doi.org/10.3390/birds3040022
Draidia A, Tareen M, Bayraktar N, Cramer ERA, Chen K-C. Bird Communities and the Rehabilitation of Al Karaana Lagoons in Qatar. Birds. 2022; 3(4):320-340. https://doi.org/10.3390/birds3040022
Chicago/Turabian StyleDraidia, Ayaterahman, Momina Tareen, Nuran Bayraktar, Emily R. A. Cramer, and Kuei-Chiu Chen. 2022. "Bird Communities and the Rehabilitation of Al Karaana Lagoons in Qatar" Birds 3, no. 4: 320-340. https://doi.org/10.3390/birds3040022
APA StyleDraidia, A., Tareen, M., Bayraktar, N., Cramer, E. R. A., & Chen, K.-C. (2022). Bird Communities and the Rehabilitation of Al Karaana Lagoons in Qatar. Birds, 3(4), 320-340. https://doi.org/10.3390/birds3040022