1. Introduction
Gamebirds (order Galliformes) play an important role in maintaining the integrity of the forests in which they live, cultivating plant community structure through seed predation dynamics, and tilling the soil routinely [
1]. Moreover, they are often a vital protein source for people and predatory wildlife alike [
2]. However, owing to their desirability as a protein source, gamebirds are often characterized by unsustainable harvest, not only for protein, but also for the feather and live bird markets [
3]. These trends, along with the wholesale destruction of the habitats they thrive in, have led to many species of gamebirds being threatened with extinction (e.g., [
4,
5]).
The Birdlife International Redlist records 15 species of gamebirds as present in Lao People’s Democratic Republic (PDR hereafter), 3 of them stable and the other 12 decreasing [
6,
7]. A limited amount of work has been done on gamebirds in Lao PDR, primarily comprising wildlife bio-inventories, which include reports of several species of gamebirds. These include Xe Bang Nouan National Protected Area (NPA, south-central Lao PDR) [
8] and Nam Et—Phou Louey NPA (northern Lao PDR) [
9,
10]. Johnson et al. [
9] camera-trapped 35 species of mammals and birds, including five species of gamebirds, and concluded that the relative abundance of small prey animals (such as gamebirds) was significantly higher where human density was low. Rasphone et al. [
10] camera-trapped 43 species and estimated the probabilities of occupancy, persistence, colonization and detection, including three species of pheasants and Rufous-throated Partridge. Additionally, Brickle et al. [
4] reviewed the Indochinese range (Lao PDR, Cambodia and Vietnam), habitat and conservation status of 21 species of gamebirds to assess the overall conservation requirements, with non-functional protected areas as the primary threat. Previous surveys recorded the presence of six species of gamebirds in and around the Namha NPA: Mountain Bamboo Partridge
Bubusicola fytchii, Rufous-throated Partridge
Arborophila rufogularis, Bar-backed Partridge
A. brunneopectus, Red Junglefowl
Gallus gallus, Silver Pheasant
Lophura nycthemera and Gray Peacock Pheasant
Polyplectron bicalcaratum [
11,
12].
For many species of gamebirds, it is extremely challenging to study intimate aspects of life history and ecology due to their shy, retiring nature. A major advantage of camera-traps is they can be used successfully in remote regions to help fill in large information gaps for cryptic birds that are difficult to observe [
13]. The studies of Savini and Sukumal [
14] and Sailo [
15], while being notable contributions for Silver Pheasant
Lophura nycthemera and Gray Peacock Pheasant
Polyplectron bicalcaratum, respectively, did not utilize camera-trapping in their methodologies. We were fortunate to opportune datasets for both of these species, thanks to the utilization of camera trap images that were bycatch from general diversity surveys [
16].
Relatively few in-depth autoecological (species-level) studies have been undertaken for the species comprising the focus of our work. Our objectives herein are to add to knowledge of the general biology and ecology of several species of gamebirds (i.e., Arborophila partridges and pheasants) and assess species associations in Namha National Protected Area (NNPA hereafter) in Luang Namtha Province, Lao PDR. While NNPA contains valuable habitat for the gamebirds studied here and other species of wildlife, there are growing threats to the continued persistence of these species. Understanding their ecology, behavior and status could inform conservation management and improve their prospects for survival. We discuss our results by comparing and contrasting our findings with other studies, and consider implications for conservation in the region.
3. Results
3.1. Camera Trap Captures
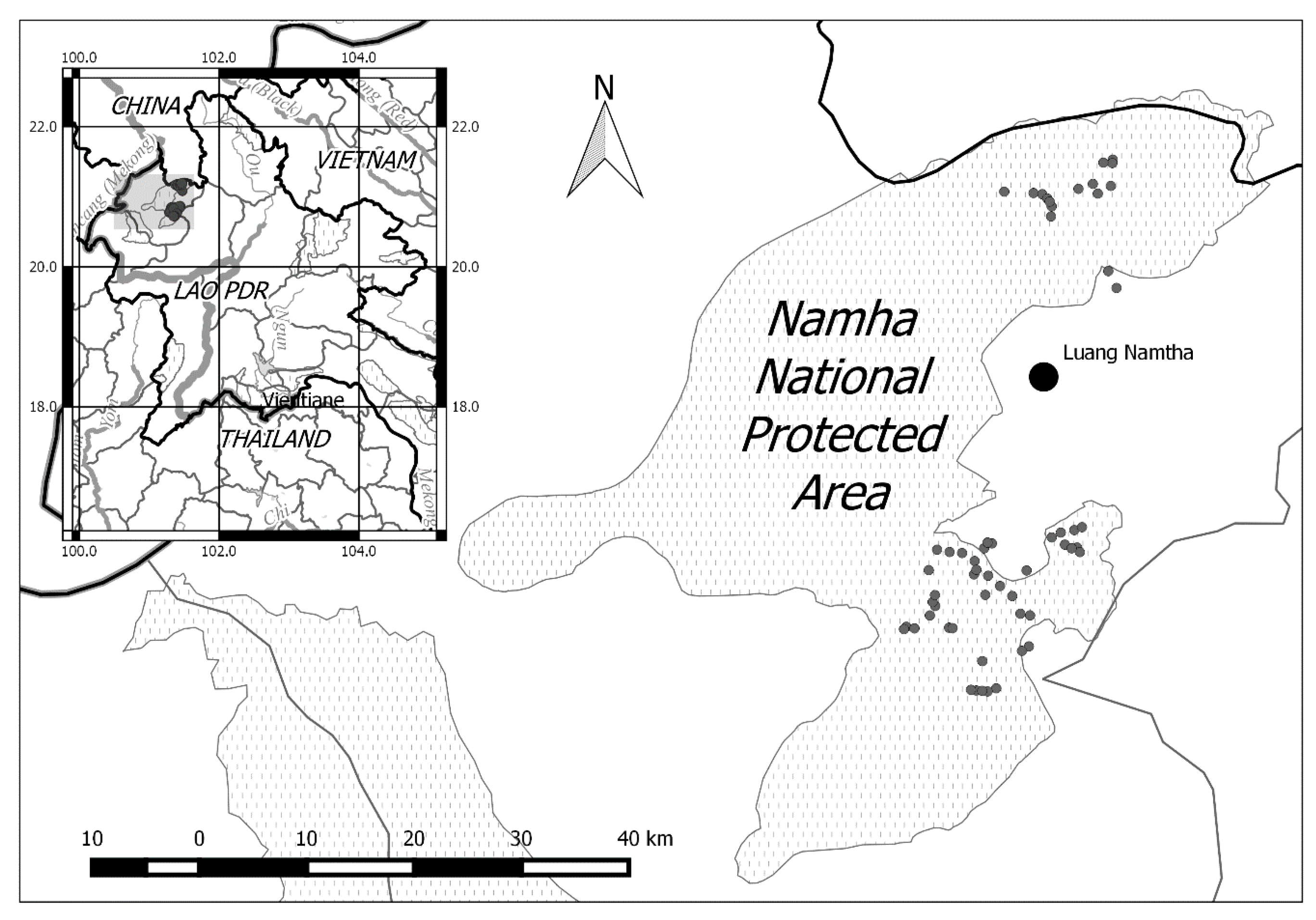
Between 6 April 2013–3 October 2015 (4003 trap days/nights), 5533 images representing 996 separate captures of wildlife were collected from 31 sampling stations in NNPA (first session). Following initial analyses, data from an additional set of 22 traps active between 7 May 2017–16 February 2019 (second session) were collated, adding an additional 4788 images from 488 captures (1947 trap days/nights). In sum, 10,321 images representing 1484 independent captures of wildlife were collected during 5950 trap days/nights of sampling.
A total of 31 species of birds were detected (
Table S1), including six species of gamebirds (
Table 1), which are highlighted in the species accounts below. A rarefaction curve [
33] (
Figure S1) shows an exponentially slowing growth in the accumulation of gamebird species over the 5950 days of sampling.
3.2. Partridges
For Rufous-throated Partridge (Arborophila rufogularis), only two birds, an adult with an approximately 75% grown subadult, were photo-trapped at 545 m asl with 68% forest cover, 2.49 km from the nearest village. This occurred at midday for 30 min on 25 May 2018.
Bar-backed Partridge (
Arborophila brunneopectus) was another rarely recorded species of partridge, recorded during five independent events (
Table 1) at the same site, 694 m asl with 80% forest cover, 3.2 km from the nearest village. Five solitary individuals (e.g.,
Figure 2A) were recorded at ca. 09:00 (
n = 2) and 11:00–15:35 (
n = 3), 8–13 March and 8 April 2014.
Scaly-breasted Partridge (
Arborophila chloropus), the third rarely recorded congener, had four independent events (
Table 1) at three different sites, averaging 612 m asl (range = 545–694) with 76% forest cover (range = 68–80), 3.38 km from the nearest village (range = 2.49–4.45). Four solitary individuals were recorded primarily during crepuscular periods (06:25 [
n = 2], 14:40–18:25 [
n = 2]) spanning 16 April–26 May.
3.3. Red Junglefowl (Gallus gallus)
This species was recorded in 21 independent events (
Table 1) at 10 different sites, averaging 777 m asl (range = 551–1011) with 75% forest cover (range = 65–80), 4.15 km from the nearest village (range = 1.34–7.28).
Twelve solitary individuals (6 males, 6 females) and a male–female pair (1:1 sex ratio) were present at camera-traps, primarily ≤ 1 min (
n = 17), also for 2, 5, 13 and 24 min (
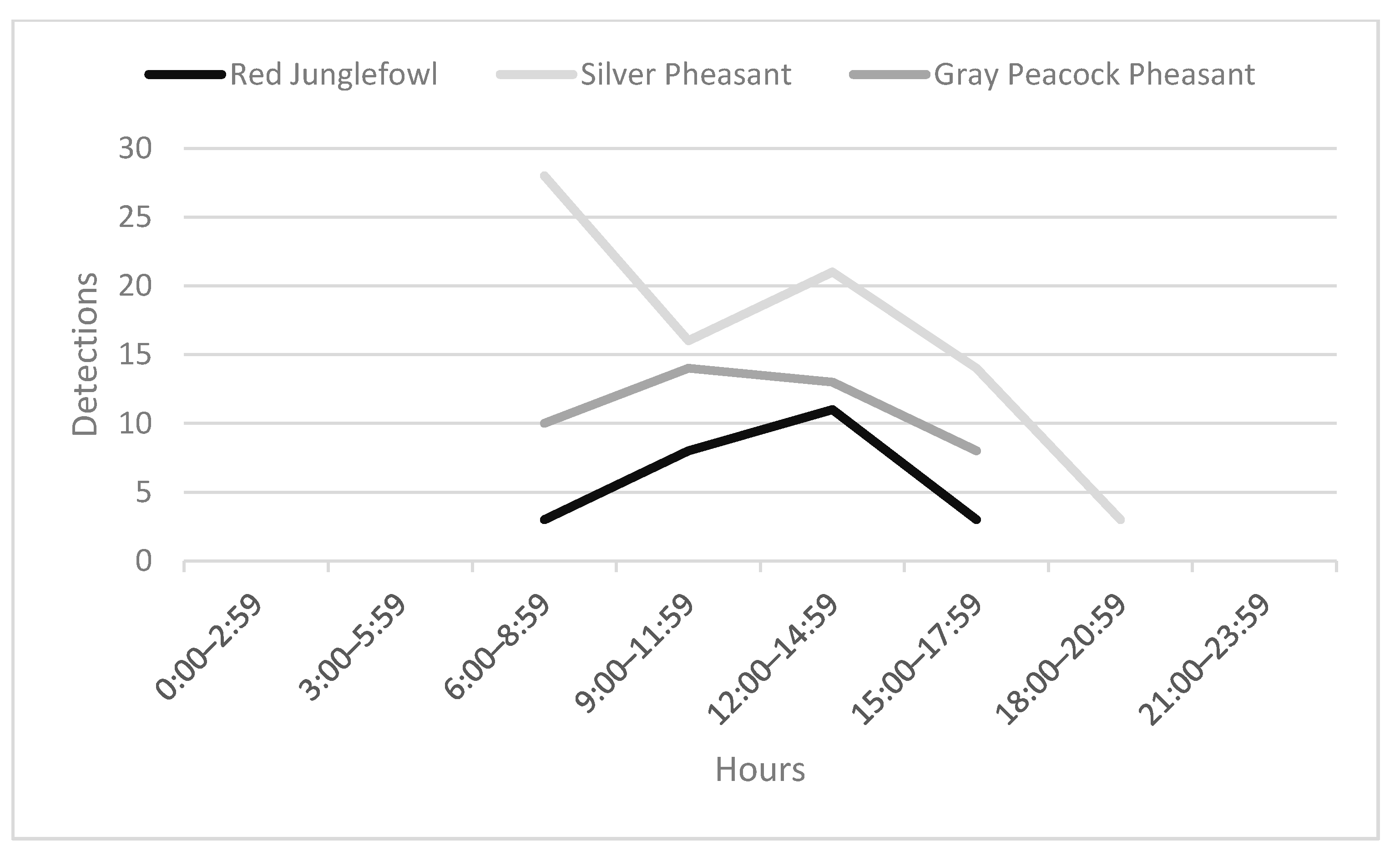
n = 1 each). There was an increase in detections towards late morning, rising to a peak just after midday (
Figure 3). Mean occupancy indicated that this species was probably available for capture at less than two-thirds of the camera-trap stations (mean = 41.9%, HPDI 14.6–68.6%;
Table S2). Mean predicted individual detectability (r, the probability of detecting an available bird within any given seven-day occasion) was 3.91% (HPDI 0.004–12.00%;
Table S2).
On 18 May 2014 at 15:02 a male–female pair of Red Junglefowl was observed associated with a boar
Sus scrofa (
Figure 2B), probably commensally eating arthropod prey displaced by the foraging boar.
3.4. Silver Pheasant (Lophura nycthemera)
This was the most frequently recorded species in this study, with 84 independent events (
Table 1) at 25 different sites, averaging 837 m asl (range = 545–1277) with 76% forest cover (range = 65–81), 4.51 km from the nearest village (range = 1.10–11.32).
The sex ratio was 1:1.13 (53 males, 60 females;
X2 = 0.319,
p = 0.572), including 13 yearling males (mid-May–mid-November) that comprised 24.5% of the male population (adult vs. yearling males:
X2 = 12.76,
p = 0.0003). Age-sex groups varied extensively, with 75.6% solitary birds (solitary vs. groups;
X2 = 25.0,
p = 0.00001) making up the majority (25 males, 32 females, 5 yearling males). Other age-sex classes (e.g.,
Figure 2C) included primarily adult male–female pairs (
n = 5), with yearling males (
n = 2); single adult males with two (
n = 3) or three (
n = 3) adult females; two adult males with a single female (
n = 1) or three adult males with two females (
n = 1); two females (
n = 3) or a single female with two yearling males (
n = 2) and two yearling males (
n = 2). Two males were growing new tail rectrices post-molt in mid-August.
Durations at camera-traps were primarily ≤ 1 min (
n = 70), also 2–10 (
n = 6), 11–25 (
n = 8) and 41–120 (
n = 3) min. Activity was bimodal, exploding at 06:00, then gradually diminishing until a second peak just after midday, until ceasing at 19:00 (
Figure 3). Occupancy was estimated to be the highest of all six species of gamebirds, indicating that this species was present at over one-third of the trap stations (mean 60.8%, HPDI 40.8–85.7%,
Table S2). Mean predicted individual detectability (r) was 6.48% (HPDI 0.88–12.30%,
Table S2).
3.5. Gray Peacock Pheasant (Polyplectron bicalcaratum)
This second most commonly detected gamebird in the study had 44 independent events (
Table 1) at nine different sites, averaging 816 m asl (range = 545–1010) with 76% forest cover (range = 68–80), 4.43 km from the nearest village (range = 2.29–7.51).
The sex ratio was 1:1.21 (19 males, 23 females;
X2 = 0.381,
p = 0.537, NS), including three yearling males (mid-March–late April; adult vs. yearling males:
X2 = 7.6,
p = 0.005) that comprised 15.7% of the male population. All individuals were solitary with the exception of three males actively displaying (e.g.,
Figure 2D) to females (late February–mid-June; solitary vs. pairs;
X2 = 26.3,
p = 0.00001).
Durations at camera-traps were primarily ≤ 1 min (
n = 45), also 3–4 (
n = 26) and 18–20 (
n = 2) min. Activity generally began around 07:00, and was somewhat bimodal, peaking mid-morning and again slightly less in mid-afternoon before terminating prior to 18:00 (
Figure 3). Occupancy was relatively low compared to the other two species of pheasants, indicating that this species was probably present at less than one-half of the trap stations (mean 28.7%, HPDI 12.2–55.1%,
Table S2). Mean predicted individual detectability (r) was 7.68% (HPDI 0.05–19.22%,
Table S2).
3.6. Species Associations
In addition to the aforementioned pair of Red Junglefowl commensally following a boar, several temporal species associations were anecdotally noted between gamebird species. Of the six species in this study, all but Scaly-breasted Partridge were occasionally photographed by the same camera within 1 h of each other (mean = 23 min, range = 3–60 min,
n = 8;
Table 2).
The primary species within these temporally proximal associations include Silver Pheasant (present in 75% of records, n = 6) and Gray Peacock Pheasant (63%, n = 5). In the case of Red Junglefowl both associations were males, whereas 60% (n = 3) of the associations were males for Gray Peacock Pheasant, and 33% contained males for Silver Pheasant (n = 2).
All cases involved solitary adults, except for a parent-offspring group of Rufous-throated Partridge associated with an adult pair of Silver Pheasants. The majority (75%,
n = 6) of these associations took place during breeding season (March–April), and many (63%,
n = 5) of the records were from a single camera trap (NH22) in March–April 2014 (
Table 2).
3.7. Environmental Associations with Occupancy
In preliminary models with 12–14 covariates, we found that the covariate factors camera model and wide-angle setting and the continuous covariates aspect and vegetation (ndvi) showed no significant association with individual detectability (
Table S3).
We found no significant differences in estimated availability (
λ) from the first and second trapping sessions and no significant effect of season or percentage of forest cover on occupancy of any species in this sample of camera stations (
Table S3).
Examining a model with only the six most influential covariates (distance to nearest village, distance to nearest tourism trail, altitude as a quadratic, terrain roughness, ndvi and aspect as a complex variable), both Silver Pheasant (SP) and Gray Peacock Pheasant (GPP) showed significantly higher estimated lambda at camera stations in rougher terrain. (
Table 3,
Tables S4 and S5; for SP, median estimated coefficient of association
= 0.538, mean 0.536 [95% HPDI 0.089–0.975], Bppp [Bayesian posterior predicted
p of absolute value > 0] = 0.010; for GPP, median 1.576, mean 1.691 [0.139–3.535], Bppp = 0.004, respectively). We found no significant effect of elevation on any species, although Gray Peacock Pheasant was significantly less associated with mid-elevations (
Table 3,
Tables S4 and S5, median −3.505, mean −3.980 [−9.183–0.184], Bppp = 0.008). Silver Pheasant availability tended to increase with increasing distance to the nearest village (median 0.473, mean 0.474 [−0.016–0.958] Bppp = 0.029). Red Junglefowl was significantly associated with sites far from trails used for nature tourism (median 1.726, mean 2.263 [−0.077 to 6.457] Bppp = 0.004).
4. Discussion
4.1. Species Detection and Occupancy
Several extremely shy species (
Table 1) that might have otherwise gone undetected were documented with camera-traps (cf. [
34]). Gamebirds, especially pheasants, are often shy due to hunting, and therefore, they are under-accounted for in avifaunal inventories [
5]. The camera-traps were set to detect species using lower strata of the forest, with eight primarily terrestrial species of birds recorded (the six gamebirds of this study as well as Rusty-naped Pitta (
Hydrornis oatesi) and White’s Thrush (
Zootheria aurea);
Table S1).
Of the six species of gamebirds detected, all were assessed by IUCN as Least Concern with Decreasing Populations [
7], but none are endemic and all have a broad geographic distribution [
35]. Silver Pheasant and Gray Peacock-Pheasant are on the list of Category 1 Protected Species in Lao PDR. Despite their reclusive nature, we are fairly confident that we detected all species of extant pheasants in the region, as they accounted for 53% (
n = 159) of the 297 total avian detections made (
Table S1). Moreover, these three species of pheasants were the only three accounted for by Johnson et al. [
9] and Rasphone et al. [
10] in northern Lao PDR, as well as in extensive market surveys [
16], appearing in the same order of relative detection as in our camera trapping study.
The smaller partridges were more rarely detected; whether this is a consequence of their smaller size not triggering the camera-traps or their relative rarity is uncertain. Of the 31 detected species of birds represented, the majority were only detected on one or two occasions (
Table S1). In light of the few partridge detections (
n = 10 events total, range = 1–5 events/sp.), it is likely that they were either too small to trigger the camera traps, or rare in the region relative to pheasants. That virtually no partridge were found during eight years of market surveys [
16], despite other species of small birds being common, supports the idea that partridge are indeed rare and seldom encountered.
We did not obtain any records of Japanese Quail (
Coturnix japonica), Blue-breasted Quail (
Synoicus chinensis), Chinese Francolin (
Francolinus pintadeanus), Mountain Bamboo Partridge or Green Peafowl (
Pavo muticus), all of which geographically overlapped the region of our study site on range maps (e.g., [
7]). These five species were not expected, as they all occupy more open habitats, including grassland, scrub, swamp and open woodland, and in some cases at higher altitudes [
35] than our camera traps were located.
4.2. Occupancy Estimates
Occupancy estimates for the three species of pheasants are available for another large tropical forest site in northern Lao PDR, Nam Et Phou Louey NPA [
10]. While methods were not identical, comparison showing occupancy 2.3 (Silver Pheasant) to 4.1 (Red Junglefowl) times higher at our site suggests that gamebirds may be more common at NNPA. Future work could address possible reasons for this, including differences in hunting intensity, in habitat or in abundance of predators.
4.3. Arborophila Partridges
Owing to the aforementioned rarity of this guild, little information was gleaned about their biology. All were found in forest with cover ranging 68–80%, and ≥2.5 km from the nearest village. This suggests that they may not tolerate human disturbance (cf. [
35]).
Judging by the age (ca. 6 weeks) of the Rufous-throated Partridge chick, it can be assumed that it hatched in mid-April. This is within the typical range of breeding dates (February–July) provided by Madge et al. [
35].
The Bar-backed Partridge was observed most often from morning through midday, whereas the Scaly-breasted Partridge was observed most often during crepuscular periods. While this information is apparently novel, numbers of observations were too limited to draw firm conclusions. While other studies have examined niche partitioning in
Arborophila partridges (e.g., [
36,
37]), the focus was on habitat separation rather than temporal asynchrony of circadian activity patterns (e.g., [
38]). Whether there could be a form of temporal niche allocation is unknown, but it certainly warrants further study.
4.4. Red Junglefowl (Gallus gallus)
We found junglefowl begin their activity post-dawn, then were moderately active until a sharp peak post-midday, followed by lower activity levels, until detections fell to zero by 18:00 (
Figure 3). This is similar to what was reported by Johnsgard [
1], where the species is active from dawn until around 09:00, then resting midday until becoming active again from mid-afternoon until dark.
The behavior of Red Junglefowl following large ungulates to consume displaced arthropods has been observed in association with several species in Thailand, including gaur
Bos gaurus [
39], sambar
Rusa unicolor (Kelvin Lim unpubl. data) and boar (Marcus Chua unpubl. data). Our preliminary observation of the boar association reported here is apparently the first published record involving this commensalism outside of Thailand.
4.5. Silver Pheasant (Lophura nycthemera)
The sex ratio found in our study in Lao PDR was 1:1.13, which is mostly similar to sex ratios found in other studies. In central Thailand, the sex ratio averaged 1:1.6 over 5 months; however, the ratio during a given month changed between January (1:1.3) when there were more females, to May (1:0.7), when there were slightly more males [
14]). In Guangdong Province, China, the sex ratio varied depending upon the habitat substrate, that is 1:1 in dry soil and 1:3 in low grassland [
40].
The observation that yearling males (recorded mid-May–mid-November) comprised one-fourth of the male population is apparently a novel record of age-class distribution for this species, as are records of males growing new tail rectrices in mid-August. The description of age-class ratios are important data to help build life history tables to estimate population growth. Molt is relevant species biology information for sound management decisions (i.e., when individuals are most vulnerable). More detailed study is warranted on age structure and molt in Silver Pheasant populations.
Our findings of 75% of all records as solitary birds was not shared by others. Johnsgard [
1] considered the species to travel mostly in groups. For example, Savini and Sukumal [
14] found mean group size to range from 3.0–4.1 between January and May, but no results were reported outside of these months. Perhaps these differences can be attributed to these authors focusing strictly on social groups rather than incorporating solitary individuals into their datasets.
We recorded constant diurnal activity with bimodal peaks, albeit diminishing until complete termination by 19:00. Similarly, Beebe [
41] observed a general bimodal trend, active early morning and late afternoon, while completely ceasing activity during midday. Differing from both of these studies, in Guangdong Province, China, activity was fairly constant between 07:00–18:00, although it slightly diminished during mid-morning [
42]. While the starting and ending times were similar, the pattern in China was unimodal (more constant) rather than bimodal, as seen previously [
41] and here.
4.6. Gray Peacock Pheasant (Polyplectron bicalcaratum)
The sex ratio of peacock pheasants in our study was 1:1.21. In contrast, males were more abundant at two wildlife sanctuaries in Mizoram state, India: more than two-fold at Lengteng (2.47:1), and less dramatically at Tawi (1.29:1). These differences can perhaps be explained by bias, with auditory counts (calls performed only by males) being incorporated into the data collection methods by Sailo [
15].
The three yearling males recorded (mid-March–late April) comprising 15.7% of the male population is apparently a new estimate of age-class for this species. Nonetheless, more detailed work on age structure is needed within Gray Peacock Pheasant populations.
We recorded three male peacock pheasants actively displaying between late February and mid-June. This is only slightly earlier than the typical estimate of the breeding season beginning in March (through June) for this species (cf. [
35]).
Activity at our site generally begins around 07:00, and is somewhat bimodal, peaking mid-morning and again slightly less in mid-afternoon before terminating before 18:00 (
Figure 3). In Arunachal Pradesh, India, this species was active much earlier (03:00–11:00), peaking from 06:00–08:00 [
43]. While the overall times, as well as durations in Lao PDR (11 h) versus India (8 h) were different, activity still occurred around dawn in both studies.
4.7. Species Associations
With the exception of Scaly-breasted Partridge, the remaining five species in this study were occasionally photographed by the same camera ≤ 1 h of each other, with the primary species being Silver Pheasant and Gray Peacock Pheasant (
Table 2). In order to be considered a temporal association, the two species should be associated non-randomly not just in space but in time. For example, considering only events at camera trap station NR30, the probability that Red Junglefowl was randomly captured within 1 h before or after the three events of Silver Pheasant capture can be estimated as
p = 0.004. The probability that Rufous-throated Partridge was randomly captured within 1 h of one of the five events of other gamebird capture at camera trap station NR12 was
p = 0.008. For Bar-backed Partridge at NH22, where it was captured five times, the probability was
p = 0.19.
In addition to multiple associations noted by Beebe [
41], there have been other noted species associations, including
Arborophila Partridges in Cambodia [
37],
Arborophila and
Tropicoperdix Partridges in Viet Nam [
44], and Siamese Fireback
L. diardi and Silver Pheasant in Thailand [
14]. Temporal species associations provide a metric of resource allocation, and ultimately insight of competition or coexistence within a given community. However, these studies are traditionally employed for mammal communities, especially carnivores (e.g., [
45,
46]), rather than gamebirds (e.g., [
38]), especially gamebird communities in the Eastern Hemisphere. Nonetheless, such studies could prove fruitful if a fertile study site for such dynamics can be located. For example, nearly two-thirds of the records for the associations in
Table 2 came from a single camera-trap (NH22), primarily during the breeding season (March–April); thus, there was a narrow window of opportunity. Incidentally, NH22 was a special site, representing lowland forest proximal to a stream with large trees, rich understory and probably deep soil.
4.8. Environmental Associations
Both Gray Peacock Pheasant and Silver Pheasant occupancy was significantly associated with rougher terrain, while Red Junglefowl showed a slight tendency to be associated with lower elevations in this sample (
Table 3). Terrain roughness is measured as the largest difference in elevation between the target grid cell and adjacent grid cells, roughly isomorphic with slope [
22]. The results suggest that, in NNPA, Silver Pheasant and Gray Peacock Pheasant may favor steep slopes, while Red Junglefowl is less fastidious.
We found that Red Junglefowl showed significantly less occupancy near well-groomed trails used for trekking tourism. Silver Pheasant and Gray Peacock Pheasant showed a slight tendency for this as well (
Table 3). Silver Pheasant also showed a tendency for lower occupancy near villages. Red Junglefowl may be particularly easy to hunt because of its obvious call, which can be heard from a long distance, and hence, it may be especially vulnerable near well-traveled routes. Alternatively, this species may simply be more wary than others and avoid areas with regular human presence.
4.9. Conservation
While the presence of three sympatric species of partridges inside NNPA is encouraging for the future of these species, the low number of detections as well as the relatively low occupancy of all gamebirds is worrying and probably reflects intensive hunting, especially near accessible trails and villages. Partridges in particular are poorly studied in the region. While assumed to be of Least Concern, their low rates of detection found here suggest that they may be rarer than often assumed. Yu et al. [
36] suggested that the Bar-backed Partridge, with its restricted distribution, small population and low ecological adaptability, might be the most vulnerable gamebird species in the karst forests of Guangxi.
Arborophila Partridges might, thus, be particularly vulnerable to hunting or habitat degradation. Patrolling, enforcement and behavioral change campaigns are needed to ensure that hunting is within sustainable limits, and is excluded from the Totally Protected Zone of NNPA. Over-hunting of vulnerable species can be further reduced by strictly controlling illegal sales of wildlife, and by campaigns to change attitudes and behaviors of sellers and consumers.
Other threats also need to be addressed to better protect the habitat of these species. Rubber plantations support few birds, and while swidden landscapes may be suitable for some avian species, they are not for those that depend on undisturbed old-growth forests. As climate change continues and accelerates, catastrophic crown fires in the dry season present an increasing threat to habitat, and this should be addressed with fire prevention and suppression, and reduction in greenhouse gas emissions. Conservation management efforts should be supported with uninterrupted funding and political backing to ensure the protection and restoration of natural habitat, continuous monitoring, thorough removal of snares and successful prosecution of repeat offenders apprehended by the newly formed patrol teams.
5. Conclusions
Camera-trapping records from a general survey provided useful data for study of six species of gamebirds, including three species of partridges that are rarely observed and have been under-studied in the region. While the dataset was small for partridges, in part because cameras were not placed to optimize capture of these species, the data were sufficient to suggest some specific conclusions about their ecology, occurrence and potential threats to their survival. Future studies might be able to avoid problems of small numbers of detection by setting more cameras closer to the ground (e.g., 35 cm) and farther apart in space (700 m or more between cameras). Simulations might then be able to address questions of temporal associations, and avoidance, between species. This could be particularly valuable for partridges, which are little-studied and perhaps rarer than assumed. Researchers should consider the value of “data-mining” other general camera trap surveys even if not optimized for the study of gamebirds. The results of such efforts will add to the body of information about these and other little-studied terrestrial birds, increasing understanding of their ecology and supporting efforts for their conservation.








{kind=link}
{kind=link}
{kind=link}