
Evaluation of Accuracy and Precision of the Sound-Recorder-Based Point-Counts Applied in Forests and Open Areas in Two Locations Situated in a Temperate and Tropical Regions




Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Soundscape Recording and Analysis
2.3. Statistical Analyses
3. Results
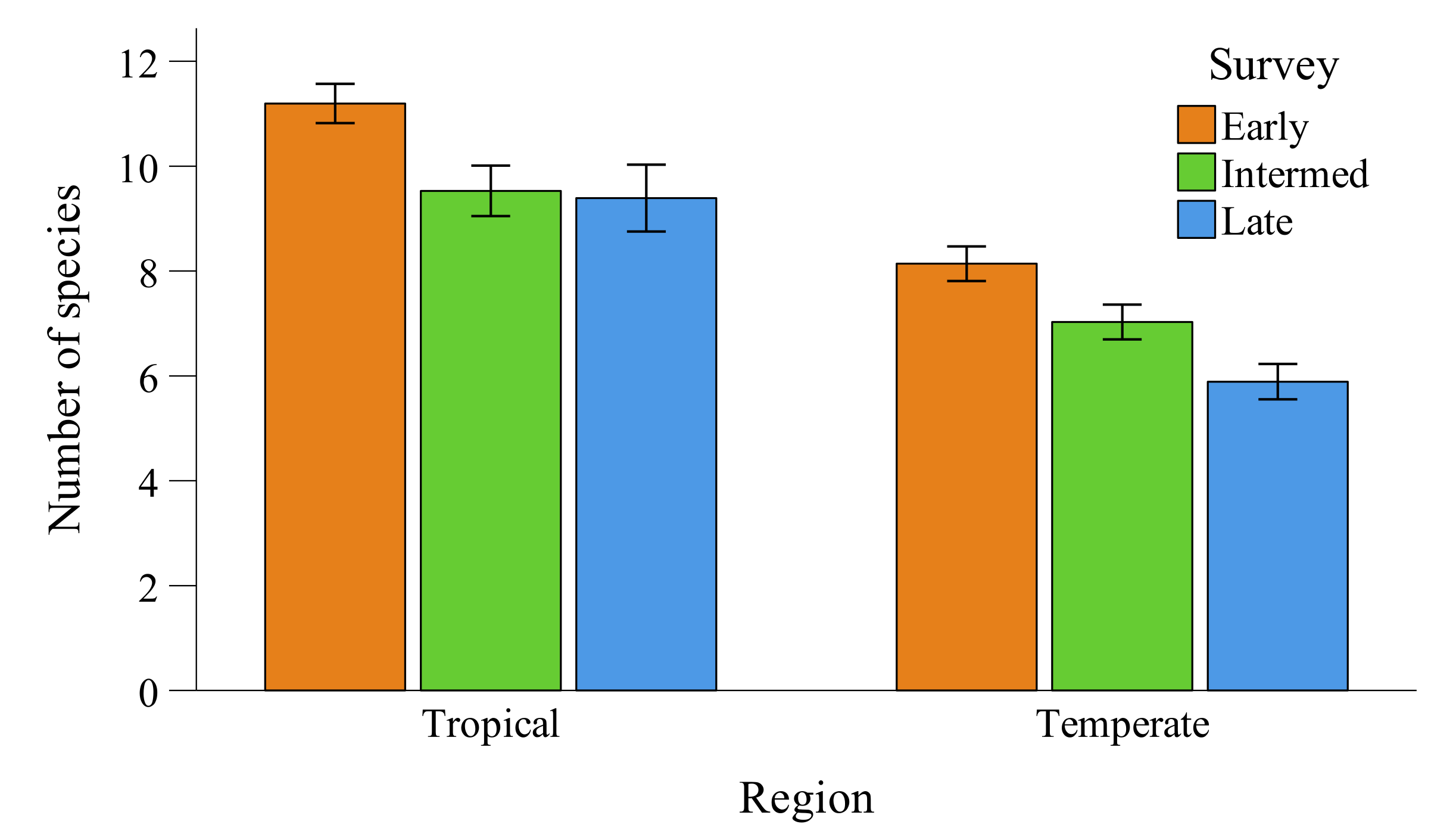
3.1. Species Richness
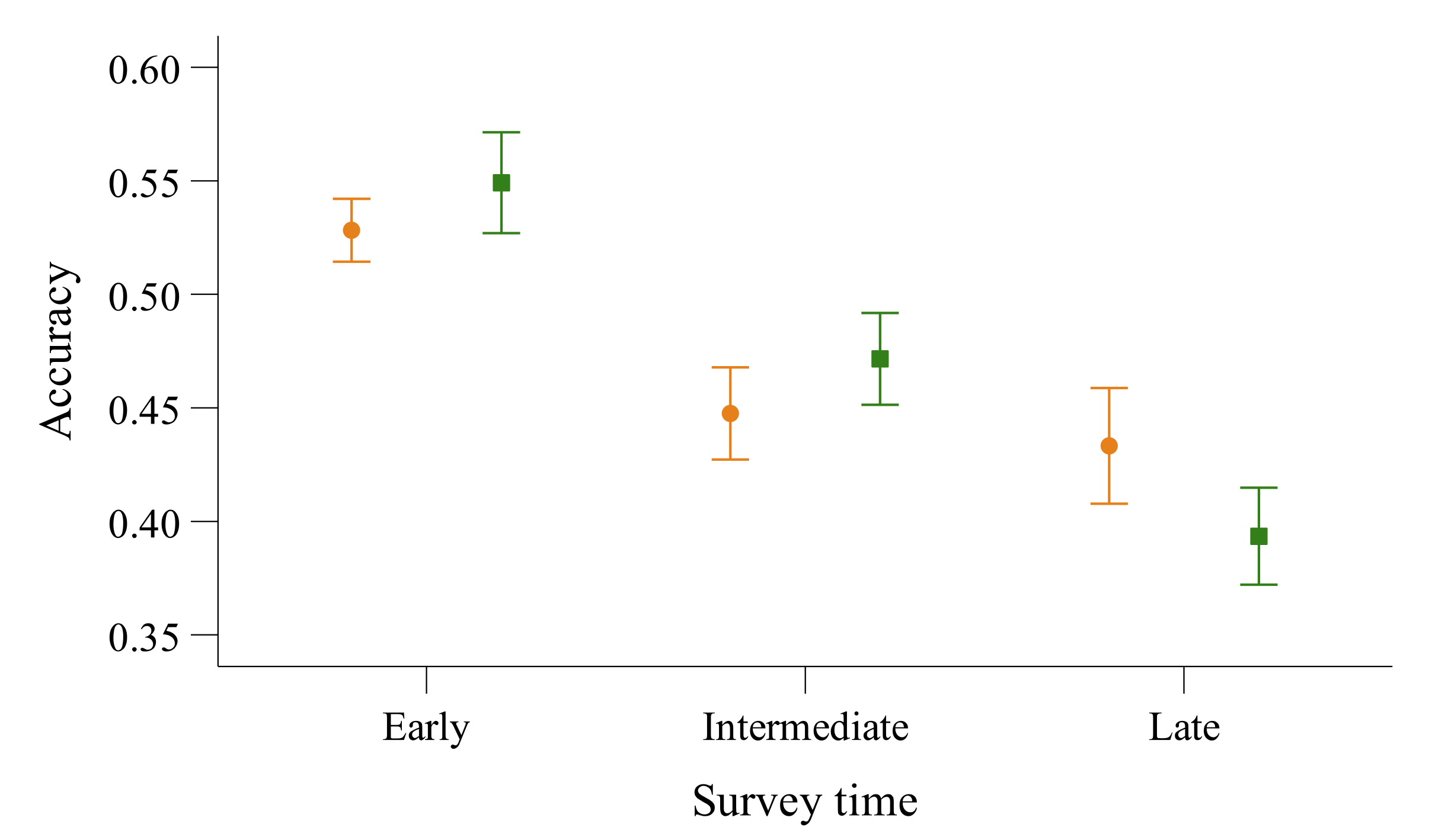
3.2. Accuracy and Precision of 5-min Surveys
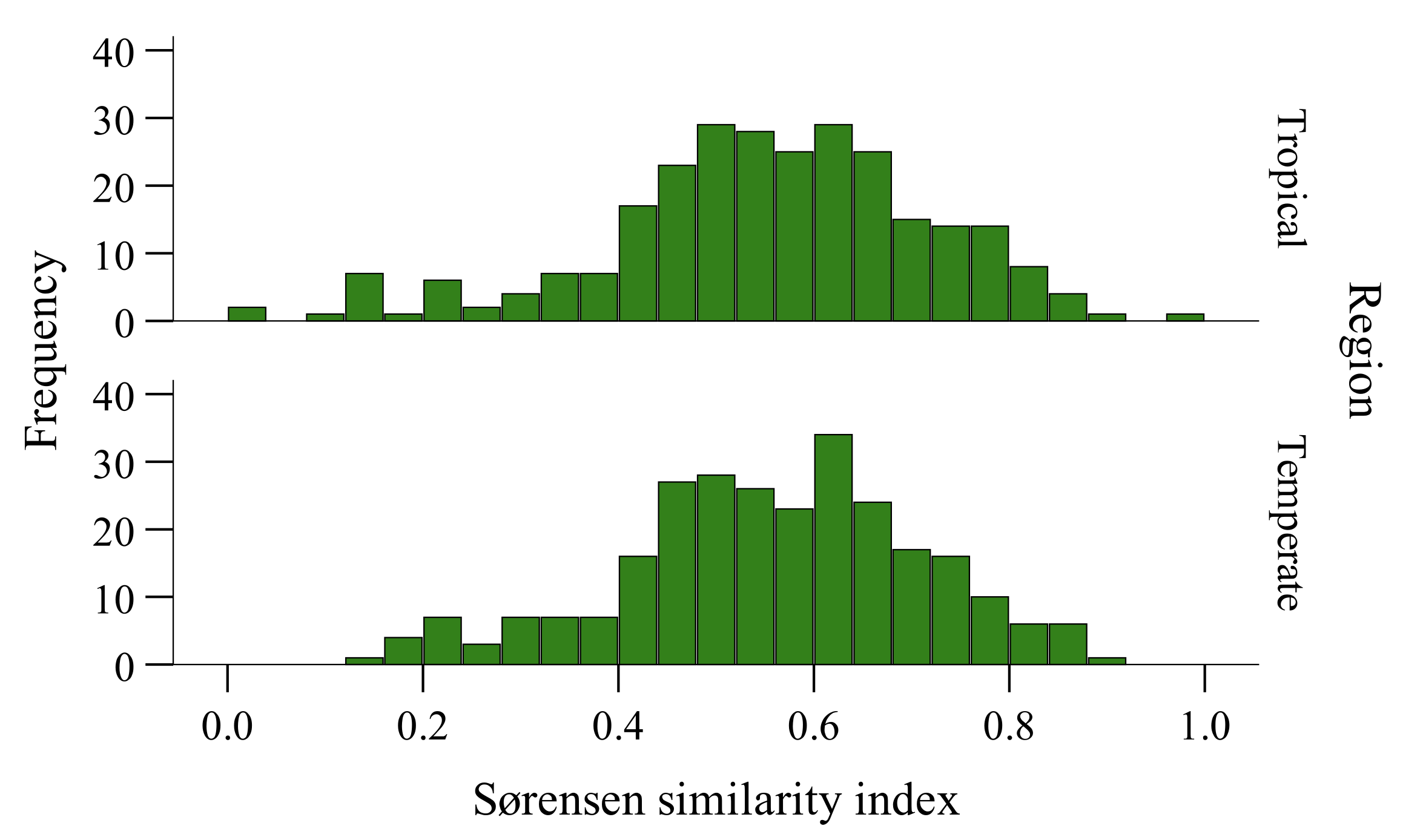
3.3. Species Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bibby, C.J.; Hill, D.A.; Burgess, N.D.; Mustoe, S. Bird Census Techniques, 2nd ed.; Academic Press: London, UK, 2000. [Google Scholar]
- Matsuoka, S.M.; Mahon, C.L.; Handel, C.M.; Sólymos, P.; Bayne, E.M.; Fontaine, P.C.; Ralph, C.J. Reviving common standards in point-count surveys for broad inference across studies. Condor 2014, 116, 599–608. [Google Scholar] [CrossRef] [Green Version]
- Crisol-Martínez, E.; Moreno-Moyano, L.T.; Horgan, F.G. Bioacoustics Reveal Species-Rich Avian Communities Exposed to Organophosphate Insecticides in Macadamia Orchards. Birds 2020, 1, 35–52. [Google Scholar] [CrossRef]
- Ralph, C.J.; Sauer, J.R.; Droege, S. Monitoring Bird Populations by Point Counts; General Technical Report PSW-GTR-149, Pacific Southwest Research Station; U.S. Department of Agriculture, Forest Service: Albany, CA, USA, 1997. [Google Scholar]
- Gregory, R.D.; Gibbson, D.W.; Donald, P. Bird census and survey techniques. In Bird Ecology and Conservation: A Handbook of Techniques; Sutherland, W.J., Newton, I., Green, R.E., Eds.; Oxford University Press: New York, NY, USA, 2004; pp. 17–56. [Google Scholar]
- Tozer, D.C.; Abraham, K.F.; Nol, E. Improving the accuracy of counts of wetland breeding birds at the point scale. Wetlands 2006, 26, 518–527. [Google Scholar] [CrossRef]
- Gale, G.; Round, P.D.; Pierce, A.J.; Nimnuan, S.; Pattanavibool, A.; Brockelman, W.Y. A field test of distance sampling methods for a tropical forest bird community. Auk 2009, 126, 439–448. [Google Scholar] [CrossRef]
- Fuller, R.J.; Langslow, D.R. Estimating numbers of birds by point counts: How long should counts last? Bird Study 1984, 31, 195–202. [Google Scholar] [CrossRef]
- Budka, M.; Jobda, M.; Szałański, P.; Piórkowski, H. Effect of agri-environment measure for the Aquatic Warbler on bird biodiversity in the extensively managed landscape of Biebrza Marshes (Poland). Biol. Conserv. 2019, 239, 108279. [Google Scholar] [CrossRef]
- Burner, R.C.; Styring, A.R.; Rahman, M.A.; Sheldon, F.H. Occupancy patterns and upper range limits of lowland Bornean birds along an elevational gradient. J. Biogeogr. 2019, 46, 2583–2597. [Google Scholar] [CrossRef] [Green Version]
- Drapeau, P.; Leduc, A.; McNeil, R. Refining the use of point counts at the scale of individual points in studies of bird-habitat relationship. J. Avian Biol. 1999, 30, 367–382. [Google Scholar] [CrossRef]
- Forcey, G.M.; Anderson, J.T.; Ammer, F.K.; Whitmore, R.C. Comparison of two double-observer point-count approaches for estimating breeding bird abundance. J. Wildl. Manag. 2006, 70, 1674–1681. [Google Scholar] [CrossRef]
- Howell, C.A.; Porneluzi, P.A.; Clawson, R.L.; Faaborg, J. Breeding density affects point-count accuracy in Missouri forest birds. J. Field Ornithol. 2004, 75, 123–133. [Google Scholar] [CrossRef]
- Efford, M.G.; Dawson, D.K. Effect of distance-related heterogeneity on population size estimates from point counts. Auk 2009, 126, 100–111. [Google Scholar] [CrossRef]
- Rigby, E.A.; Johnson, D.H. Factors affecting detection probability, effective area surveyed, and species misidentification in grassland bird point counts. Condor 2019, 12, 1–10. [Google Scholar] [CrossRef]
- Celis-Murillo, A.; Deppe, J.L.; Allen, M.F. Using soundscape recordings to estimate bird species abundance, richness, and composition. J. Field Ornithol. 2009, 80, 64–78. [Google Scholar] [CrossRef]
- Darras, K.; Batáry, P.; Furnas, B.J.; Grass, I.; Mulyani, Y.A.; Tscharntke, T. Autonomous sound recording outperforms human observation for sampling birds: A systematic map and user guide. Ecol. Appl. 2019, 29, e01954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutto, R.L.; Stutzman, R.J. Humans versus autonomous recording units: A comparison of point-count results. J. Field Ornithol. 2009, 80, 387–398. [Google Scholar] [CrossRef]
- Wheeldon, A.; Mossman, H.L.; Sullivan, M.J.P.; Mathenge, J.; de Kort, S.R. Comparison of acoustic and traditional point count methods to assess bird diversity and composition in the Aberdare National Park, Kenya. African J. Ecol. 2019, 57, 168–176. [Google Scholar] [CrossRef]
- Sedláček, O.; Vokurkova, J.; Ferenc, M.; Djomo, E.N.; Albrecht, T.; Horak, D. A comparison of point counts with a new acoustic sampling method: A case of a bird community from the montane forests of Mount Cameroon. Ostrich 2015, 86, 1–8. [Google Scholar] [CrossRef]
- Darras, K.; Batáry, P.; Furnas, B.; Celis-Murillo, A.; van Wilgenburg, S.L.; Mulyani, Y.A.; Tscharntkeet, T. Comparing the sampling performance of sound recorders versus point counts in bird surveys: A meta-analysis. J. Appl. Ecol. 2018, 55, 2575–2586. [Google Scholar] [CrossRef]
- Brewster, J.P.; Simons, T.R. Testing the importance of auditory detections in avian point counts. J. Field Ornithol. 2008, 80, 178–182. [Google Scholar] [CrossRef]
- Yip, D.; Leston, L.; Bayne, E.; Sólymos, P.; Grover, A. Experimentally derived detection distances from audio recordings and human observers enable integrated analysis of point count data. Avian Conserv. Ecol. 2017, 12, 11. [Google Scholar] [CrossRef] [Green Version]
- Sauer, J.R.; Peterjohn, B.C.; Link, W.A. Observer differences in the North American Breeding Bird Survey. Auk 1994, 111, 50–62. [Google Scholar] [CrossRef]
- Farmer, R.G.; Leonard, M.L.; Flemming, J.E.M.; Anderson, S.C. Observer aging and long-term avian survey data quality. Ecol. Evol. 2014, 4, 2563–2576. [Google Scholar] [CrossRef] [PubMed]
- IMGW. Climate of Poland; Institute of Meteorology and Water Management National Research Institute: Warsaw, Poland, 2020. [Google Scholar]
- Budka, M. Upper Nurzec River Valley. In Important Bird Areas of International Importance in Poland; Wilk, T., Jujka, M., Krogulec, J., Chylarecki, P., Eds.; Polish Society for the Protection of Birds: Marki, Poland, 2010. [Google Scholar]
- Wesołowski, T.; Czeszczewik, D.; Hebda, G.; Maziarz, M.; Mitrus, C.; Rowiński, P. 40 years of breeding bird community dynamics in a primeval temperate forest (Białowieża National Park, Poland). Acta Ornithol. 2015, 50, 95–120. [Google Scholar] [CrossRef]
- BirdLife International. Endemic Bird Areas factsheet: Cameroon mountains. Available online: http://www.birdlife.org (accessed on 24 October 2020).
- Tye, H. The climate of the highlands of western Cameroon. In Conservation of Cameroon Montane Forests; Stuart, S.N., Ed.; International Council for Bird Preservation: Cambridge, UK, 1986; pp. 18–19. [Google Scholar]
- Innocent, N.M.; Bitondo, D.; Azibo, B.R. Climate variability and change in the Bamenda highlands of north western Cameroon: Perceptions, impacts and coping mechanisms. Br. J. Appl. Sci. Technol. 2016, 12, 1–18. [Google Scholar] [CrossRef]
- Tye, H. Reversal of breeding season by lowland birds at higher altitudes in western Cameroon. Ibis 1992, 134, 154–163. [Google Scholar] [CrossRef]
- Sedláček, O.; Reif, J.; Hořák, D.; Riegert, J.; Pešata, M.; Klvana, P. The birds of a montane forest mosaic in Big Babanki area, Bamenda Highlands, Cameroon. Malimbus 2007, 29, 89–100. [Google Scholar]
- Bonthoux, S.; Balent, G. Point count duration: Five minutes are usually sufficient to model the distribution of bird species and to study the structure of communities for a French landscape. J. Ornithol. 2012, 153, 491–504. [Google Scholar] [CrossRef]
- Hardin, J.W.; Hilbe, J.M. Generalized Estimating Equations, 2nd ed.; Chapman and Hall/CRC: Boca Raton, FL, USA, 2013. [Google Scholar]
- Sørensen, T. A method of establishing groups of equal amplitude in plant sociology based on similarity of species and its application to analyses of the vegetation on Danish commons. K. Dan. Vidensk. Selsk. 1948, 5, 1–34. [Google Scholar]
- Kułaga, K.; Budka, M. Bird species detection by an observer and an autonomous sound recorder in two different environments: Forest and farmland. PLoS ONE 2019, 14, e0211970. [Google Scholar] [CrossRef]
- Harms, T.M.; Dinsmore, S.J. Optimizing surveys for marsh songbirds: Does timing matter? J. Field Ornithol. 2018, 89, 276–286. [Google Scholar] [CrossRef] [Green Version]
- Kéry, M.; Schmidt, B.R. Imperfect detection and its consequences for monitoring for conservation. Comm. Ecol. 2008, 9, 207–216. [Google Scholar] [CrossRef] [Green Version]
- Sliwinski, M.; Powell, L.; Koper, N.; Giovanni, M.; Schacht, W. Research design considerations to ensure detection of all species in an avian community. Methods Ecol. Evol. 2016, 7, 456–462. [Google Scholar] [CrossRef]
- MacKenzie, D.; Nichols, J.; Lachman, G.; Droege, G.S.; Royle, J.; Langtimm, C. Estimating site occupancy rates when detection probabilities are less than one. Ecology 2002, 83, 2248–2255. [Google Scholar] [CrossRef]
- Gu, W.; Swihart, R.K. Absent or undetected? Effects of non-detection of species occurrence on wildlife–habitat models. Biol. Conserv. 2003, 116, 195–203. [Google Scholar] [CrossRef]
- Dettmers, R.; Buehler, D.; Bartlett, J.; Klaus, N. Influence of point count length and repeated visits on habitat model performance. J Wild Manag. 1999, 63, 815–823. [Google Scholar] [CrossRef]
- Budka, M.; Czyż, M.; Skierczyńska, A.; Skierczyński, M.; Osiejuk, T.S. Duration of survey changes interpretation of habitat preferences: An example of an endemic tropical songbird, the Bangwa Forest Warbler. Ostrich 2020, 91, 195–203. [Google Scholar] [CrossRef]
- Yip, D.; Bayne, E.; Sólymos, P.; Campbell, J.; Proppe, D. Sound attenuation in forest and roadside environments: Implications for avian point-count surveys. Condor 2017, 119, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Catchpole, C.K. The functions of advertising song in the Sedge Warbler (Acrocephalus schoenobaenus) and the Reed Warbler (A. scirpaceus). Behaviour 1973, 46, 300–319. [Google Scholar] [CrossRef]
- Gil, D.; Llusia, D. The bird dawn chorus revisited. In Coding Strategies in Vertebrate Acoustic Communication; Aubin, T., Mathevon, N., Eds.; Springer Nature: London, UK, 2020; pp. 45–92. [Google Scholar]
- Bateman, H.L.; Riddle, S.B.; Cubley, E.S. Using Bioacoustics to Examine Vocal Phenology of Neotropical Migratory Birds on a Wild and Scenic River in Arizona. Birds 2021, 2, 261–274. [Google Scholar] [CrossRef]
- Haselmayer, J.; Quinn, J.S. A comparison of point counts and sound recording as bird survey methods in Amazonian Southeast Peru. Condor 2000, 102, 887–893. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Predictor | B | SE | Wald χ2 | df | p-Value |
|---|---|---|---|---|---|
| Intercept | 2.442 | 0.0635 | 1480.179 | 1 | <0.001 |
| Region (temperate) | −0.362 | 0.0703 | 26.536 | 1 | <0.001 |
| Habitat (open habitats) | −0.012 | 0.0705 | 0.028 | 1 | 0.867 |
| Day (second) | 0.002 | 0.0323 | 0.002 | 1 | 0.961 |
| Survey (late) | −0.247 | 0.0528 | 21.890 | 1 | <0.001 |
| Survey (intermediate) | −0.154 | 0.0335 | 21.050 | 1 | <0.001 |
| Predictor | B | SE | Wald χ2 | df | p-Value |
|---|---|---|---|---|---|
| Intercept | −0.628 | 0.0439 | 205.055 | 1 | <0.001 |
| Region (temperate) | −0.003 | 0.0501 | 0.003 | 1 | 0.955 |
| Habitat (open habitats) | 0.032 | 0.0505 | 0.396 | 1 | 0.529 |
| Day (second) | −0.008 | 0.0355 | 0.045 | 1 | 0.832 |
| Survey (late) | −0.264 | 0.552 | 22.933 | 1 | <0.001 |
| Survey (intermediate) | −0.160 | 0.0346 | 21.253 | 1 | <0.001 |
| Predictor | B | SE | Wald χ2 | df | p-Value |
|---|---|---|---|---|---|
| Intercept | 0.546 | 0.0266 | 420.792 | 1 | <0.001 |
| Region (temperate) | 0.004 | 0.0294 | 0.020 | 1 | 0.888 |
| Habitat (open habitats) | 0.016 | 0.0294 | 0.292 | 1 | 0.589 |
| Day (different) | −0.027 | 0.0106 | 6.361 | 1 | 0.012 |
| Time of survey (the same) | 0.069 | 0.0144 | 22.953 | 1 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Budka, M.; Kułaga, K.; Osiejuk, T.S. Evaluation of Accuracy and Precision of the Sound-Recorder-Based Point-Counts Applied in Forests and Open Areas in Two Locations Situated in a Temperate and Tropical Regions. Birds 2021, 2, 351-361. https://doi.org/10.3390/birds2040026
Budka M, Kułaga K, Osiejuk TS. Evaluation of Accuracy and Precision of the Sound-Recorder-Based Point-Counts Applied in Forests and Open Areas in Two Locations Situated in a Temperate and Tropical Regions. Birds. 2021; 2(4):351-361. https://doi.org/10.3390/birds2040026
Chicago/Turabian StyleBudka, Michał, Kinga Kułaga, and Tomasz Stanislaw Osiejuk. 2021. "Evaluation of Accuracy and Precision of the Sound-Recorder-Based Point-Counts Applied in Forests and Open Areas in Two Locations Situated in a Temperate and Tropical Regions" Birds 2, no. 4: 351-361. https://doi.org/10.3390/birds2040026
APA StyleBudka, M., Kułaga, K., & Osiejuk, T. S. (2021). Evaluation of Accuracy and Precision of the Sound-Recorder-Based Point-Counts Applied in Forests and Open Areas in Two Locations Situated in a Temperate and Tropical Regions. Birds, 2(4), 351-361. https://doi.org/10.3390/birds2040026