
Fecal Glucocorticoid Metabolite Concentration as a Tool for Assessing Impacts of Interventions in Humboldt Penguins (Spheniscus humboldti)



{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Analysis of Fecal Glucocorticoid Metabolites (ELISA)
2.3. Statistical Analysis
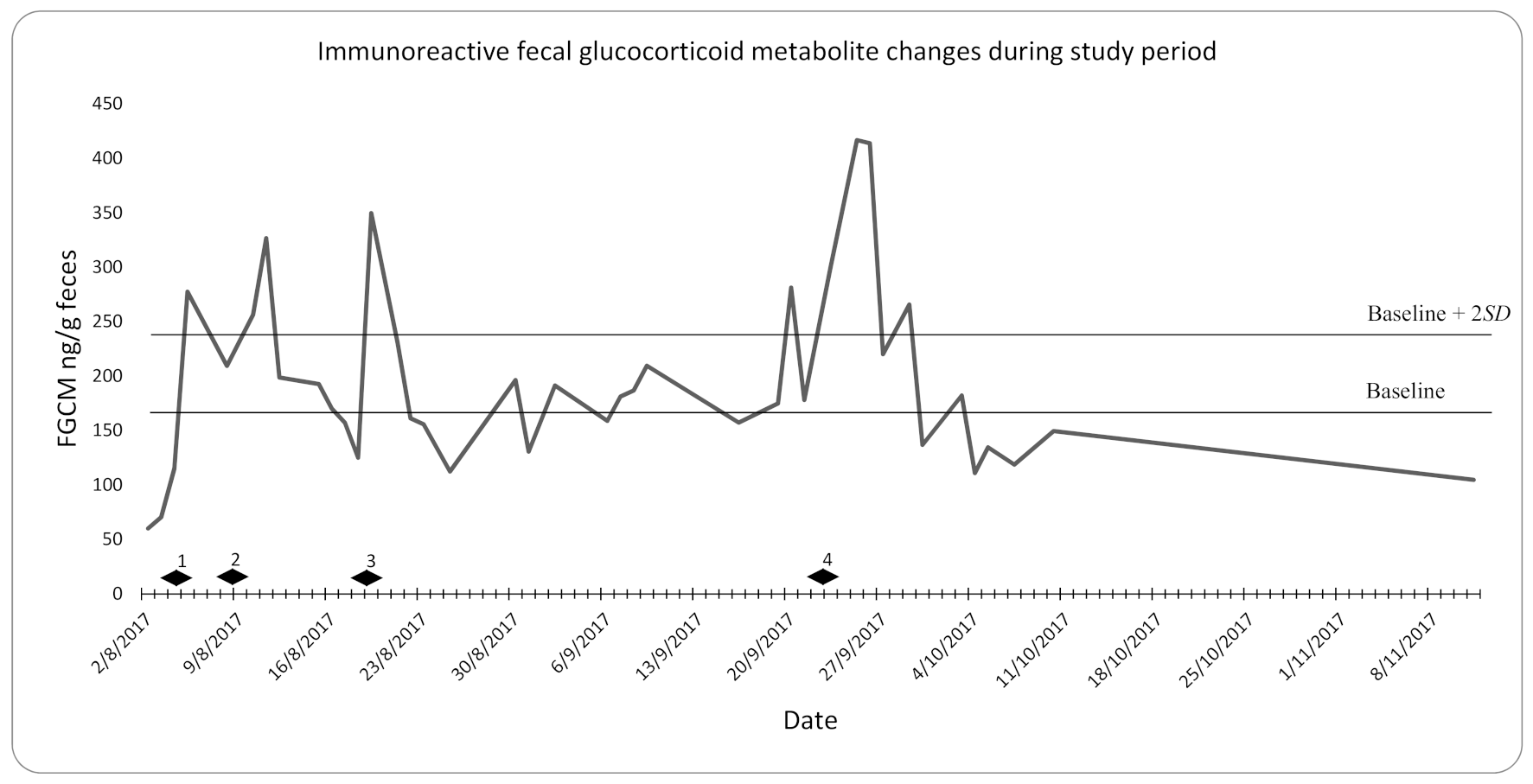
- Transport from holding containers to the exhibit (4 August 2017)
- Veterinary examination (8 August 2017)
- Transport from the exhibit to holding containers (18 August 2017)
- Transport from holding containers to the exhibit (22 September 2017).
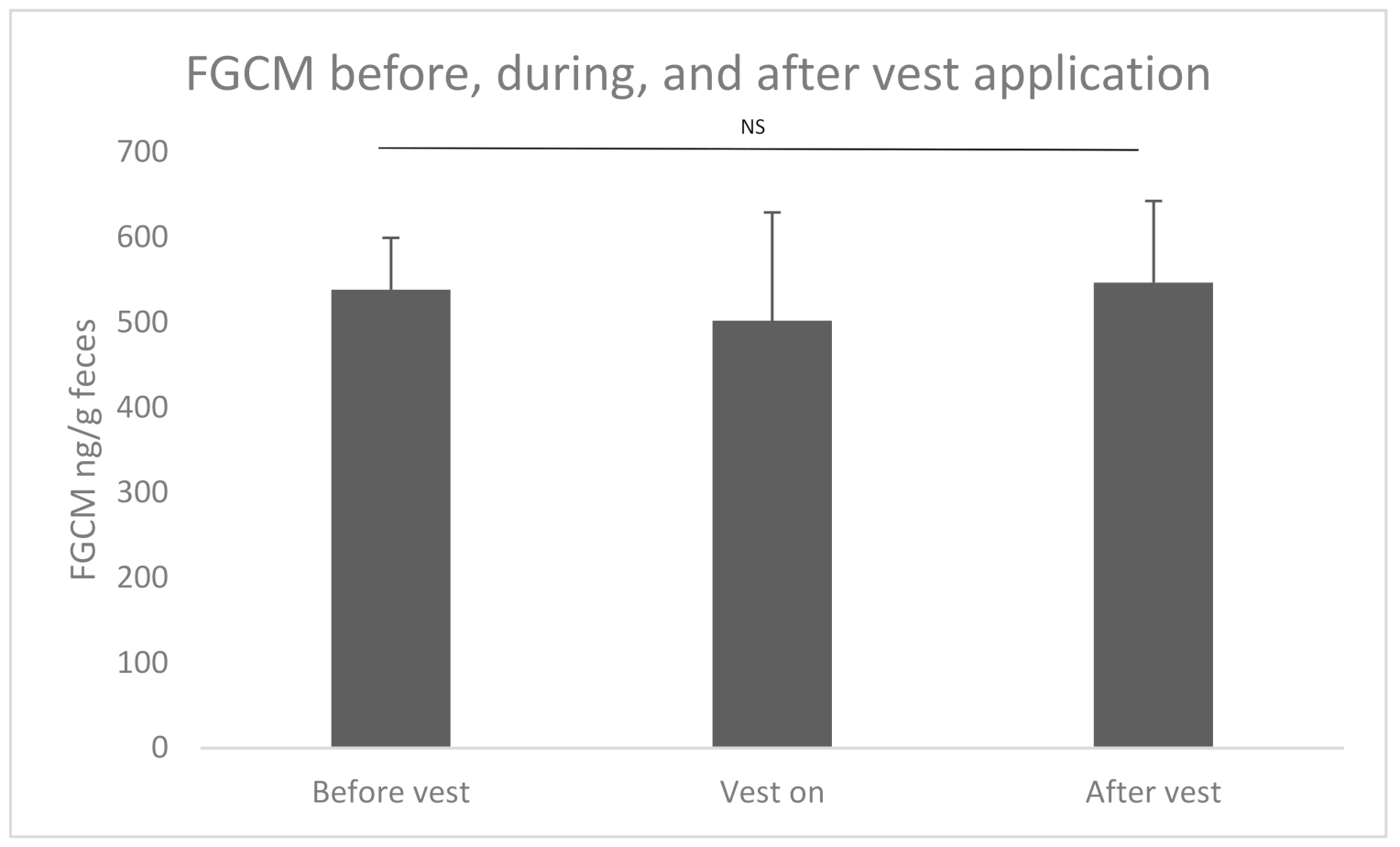
3. Results
3.1. Capture/Transport Stress
3.2. Accelerometer Vest Application
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ridgely, R.S.; Greenfield, P.J. The Birds of Ecuador: Status, Distribution, and Taxonomy; Cornell University Press: New York, NY, USA, 2001; Volume 1. [Google Scholar]
- Williams, T.D.; Busby, J. Bird Families of the World. 2. The Penguins: Spheniscidae; Oxford University Press: Oxford, UK, 1995. [Google Scholar]
- Hulsman, A.; Dalerum, F.; Ganswindt, A.; Muenscher, S.; Bertschinger, H.J.; Paris, M. Non-invasive monitoring of glucocorticoid metabolites in brown hyaena (Hyaena brunnea) feces. Zoo Biol. 2011, 30, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Anfossi, L.; Ozella, L.; Di Nardo, F.; Giovannoli, C.; Passini, C.; Favaro, L.; Pessani, D.; Mostl, E.; Baggiani, C. A broad-selective enzyme immunoassay for non-invasive stress assessment in African penguins (Spheniscus demersus) held in captivity. Anal. Methods 2014, 6, 8222–8231. [Google Scholar] [CrossRef]
- Walker, B.G.; Dee Boersma, P.; Wingfield, J.C. Habituation of adult Magellanic penguins to human visitation as expressed through behavior and corticosterone secretion. Conserv. Biol. 2006, 20, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Hood, L.C.; Boersma, P.D.; Wingfield, J.C. The adrenocortical response to stress in incubating Magellanic penguins (Spheniscus magellanicus). Auk 1998, 115, 76–84. [Google Scholar]
- Carroll, G.; Turner, E.; Dann, P.; Harcourt, R. Prior exposure to capture heightens the corticosterone and behavioural responses of little penguins (Eudyptula minor) to acute stress. Conserv. Physiol. 2016, 4. [Google Scholar] [CrossRef]
- Mostl, E.; Rettenbacher, S.; Palme, R. Measurement of corticosterone metabolites in birds’ droppings: An analytical approach. Ann. N. Y. Acad. Sci. 2005, 1046, 17–34. [Google Scholar] [CrossRef]
- Watson, R.; Munro, C.; Edwards, K.L.; Norton, V.; Brown, J.L.; Walker, S.L. Development of a versatile enzyme immunoassay for non-invasive assessment of glucocorticoid metabolites in a diversity of taxonomic species. Gen. Comp. Endocrinol. 2013, 186, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Touma, C.; Palme, R. Measuring fecal glucocorticoid metabolites in mammals and birds: The importance of validation. Ann. N. Y. Acad. Sci. 2005, 1046, 54–74. [Google Scholar] [CrossRef]
- Dehnhard, M.; Schreer, A.; Krone, O.; Jewgenow, K.; Krause, M.; Grossmann, R. Measurement of plasma corticosterone and fecal glucocorticoid metabolites in the chicken (Gallus domesticus), the great cormorant (Phalacrocorax carbo), and the goshawk (Accipiter gentilis). Gen. Comp. Endocrinol. 2003, 131, 345–352. [Google Scholar] [CrossRef]
- Narayan, E.J.; Webster, K.; Nicolson, V.; Mucci, A.; Hero, J.-M. Non-invasive evaluation of physiological stress in an iconic Australian marsupial: The Koala (Phascolarctos cinereus). Gen. Comp. Endocrinol. 2013, 187, 39–47. [Google Scholar] [CrossRef]
- Palme, R. Measuring fecal steroids: guidelines for practical application. Ann. N. Y. Acad. Sci. 2005, 1046, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Ozella, L.; Anfossi, L.; Di Nardo, F.; Pessani, D. Non-invasive monitoring of adrenocortical activity in captive African Penguin (Spheniscus demersus) by measuring faecal glucocorticoid metabolites. Gen. Comp. Endocrinol. 2015, 224, 104–112. [Google Scholar] [CrossRef]
- Nakagawa, S.; Möstl, E.; Waas, J.R. Validation of an enzyme immunoassay to measure faecal glucocorticoid metabolites from Adelie penguins (Pygoscelis adeliae): A non-invasive tool for estimating stress? Polar Biol. 2003, 26, 491–493. [Google Scholar] [CrossRef]
- Ponganis, P.J. Bio-logging of physiological parameters in higher marine vertebrates. Deep Sea Res. Part II Top. Stud. Oceanogr. 2007, 54, 183–192. [Google Scholar] [CrossRef]
- Kooyman, G.; Cherel, Y.; Maho, Y.L.; Croxall, J.; Thorson, P.; Ridoux, V.; Kooyman, C. Diving behavior and energetics during foraging cycles in king penguins. Ecol. Monogr. 1992, 62, 143–163. [Google Scholar] [CrossRef]
- Ropert-Coudert, Y.; Beaulieu, M.; Hanuise, N.; Kato, A. Diving into the world of biologging. Endanger. Species Res. 2009, 10, 21–27. [Google Scholar] [CrossRef]
- Yoda, K.; Naito, Y.; Sato, K.; Takahashi, A.; Nishikawa, J.; Ropert-Coudert, Y.; Kurita, M.; Le Maho, Y. A new technique for monitoring the behaviour of free-ranging Adelie penguins. J. Exp. Biol. 2001, 204, 685–690. [Google Scholar] [PubMed]
- Yoda, K.; Sato, K.; Niizuma, Y.; Kurita, M.; Bost, C.; Le Maho, Y.; Naito, Y. Precise monitoring of porpoising behaviour of Adélie penguins determined using acceleration data loggers. J. Exp. Biol. 1999, 202, 3121–3126. [Google Scholar]
- Wilson, R.P.; Grant, W.S.; Duffy, D.C. Recording devices on free-ranging marine animals: Does measurement affect foraging performance? Ecology 1986, 67, 1091–1093. [Google Scholar] [CrossRef]
- Kooyman, G.L. Diverse Divers: Physiology and Behavior; Springer: Berlin/Heidelberg, Germany, 2012; Volume 23. [Google Scholar]
- Ropert-Coudert, Y.; Knott, N.; Chiaradia, A.; Kato, A. How do different data logger sizes and attachment positions affect the diving behaviour of little penguins? Deep Sea Res. Part II Top. Stud. Oceanogr. 2007, 54, 415–423. [Google Scholar] [CrossRef]
- Saraux, C.; Le Bohec, C.; Durant, J.M.; Viblanc, V.A.; Gauthier-Clerc, M.; Beaune, D.; Park, Y.-H.; Yoccoz, N.G.; Stenseth, N.C.; Le Maho, Y. Reliability of flipper-banded penguins as indicators of climate change. Nature 2011, 469, 203–206. [Google Scholar] [CrossRef] [PubMed]
- White, C.R.; Cassey, P.; Schimpf, N.G.; Halsey, L.G.; Green, J.A.; Portugal, S.J. Implantation reduces the negative effects of bio-logging devices on birds. J. Exp. Biol. 2013, 216, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.E.; Converse, S.J.; Chandler, J.N.; Shafer, C.; Brown, J.L.; Keefer, C.L.; Songsasen, N. Female gonadal hormones and reproductive behaviors as key determinants of successful reproductive output of breeding whooping cranes (Grus americana). Gen. Comp. Endocrinol. 2016, 230, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.L.; Wasser, S.K.; Wildt, D.E.; Graham, L.H. Comparative aspects of steroid hormone metabolism and ovarian activity in felids, measured noninvasively in feces. Biol. Reprod. 1994, 51, 776–786. [Google Scholar] [CrossRef] [PubMed]
- Romero, L.M.; Reed, J.M. Repeatability of baseline corticosterone concentrations. Gen. Comp. Endocrinol. 2008, 156, 27–33. [Google Scholar] [CrossRef]
- Angelier, F.; Wingfield, J.C.; Weimerskirch, H.; Chastel, O. Hormonal correlates of individual quality in a long-lived bird: A test of the ‘corticosterone–fitness hypothesis’. Biol. Lett. 2010, 6, 846–849. [Google Scholar] [CrossRef] [PubMed]
- Ozella, L.; Anfossi, L.; Di Nardo, F.; Pessani, D. Effect of weather conditions and presence of visitors on adrenocortical activity in captive African penguins (Spheniscus demersus). Gen. Comp. Endocrinol. 2017, 242, 49–58. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, S.; McWhorter, T.J. Fecal Glucocorticoid Metabolite Concentration as a Tool for Assessing Impacts of Interventions in Humboldt Penguins (Spheniscus humboldti). Birds 2021, 2, 106-113. https://doi.org/10.3390/birds2010007
Xie S, McWhorter TJ. Fecal Glucocorticoid Metabolite Concentration as a Tool for Assessing Impacts of Interventions in Humboldt Penguins (Spheniscus humboldti). Birds. 2021; 2(1):106-113. https://doi.org/10.3390/birds2010007
Chicago/Turabian StyleXie, Shangzhe, and Todd J. McWhorter. 2021. "Fecal Glucocorticoid Metabolite Concentration as a Tool for Assessing Impacts of Interventions in Humboldt Penguins (Spheniscus humboldti)" Birds 2, no. 1: 106-113. https://doi.org/10.3390/birds2010007
APA StyleXie, S., & McWhorter, T. J. (2021). Fecal Glucocorticoid Metabolite Concentration as a Tool for Assessing Impacts of Interventions in Humboldt Penguins (Spheniscus humboldti). Birds, 2(1), 106-113. https://doi.org/10.3390/birds2010007