Simple Summary
Accounts of neurotoxic manifestations in patients exposed to gadobutrol have been anecdotally reported. Given the seriousness of the reported adverse effects, more research is needed to study the causes and the possible mechanisms underlying this toxicity. The aim of this study was to evaluate the safety of gadobutrol by assessing its effect on the nervous system under physiological and inflammatory conditions. Our results showed that, in normal male Sprague Dawley rats, repeated injections of gadobutrol over 20 days can cause gadolinium deposition in the brain, spinal cord, and peripheral nerves with no behavioral changes. On the other hand, rats with a lipopolysaccharide-induced inflammation present higher gadolinium deposition and heightened pain sensitivity. These findings can help guide the administration of gadobutrol in patients with inflammatory diseases or those requiring multiple MRI scans.
Abstract
This study aimed to evaluate the safety of gadobutrol, a gadolinium-based contrast agent used in medical imaging, by investigating its effect on the nervous system under physiological and inflammatory conditions. Male Sprague Dawley rats were divided randomly into four groups, including gadobutrol, saline, LPS + gadobutrol, and LPS + saline, and were given intraperitoneal injections of gadobutrol (2.5 mmol/kg) or saline for 20 days. Weekly sensorimotor and cognitive behavioral tests were performed over 4 weeks, and Gd concentration in nervous tissues was analyzed using inductively coupled plasma mass spectrometry (ICP-MS). Lactate dehydrogenase (LDH) activity was measured to evaluate cytotoxicity, and electromyography (EMG) recordings from the gastrocnemius muscle were also obtained to examine signal transmission in sciatic nerves. The results indicated that gadobutrol did not induce significant behavioral changes under normal conditions. However, when administered along with LPS, the combination led to behavioral dysfunction. ICP-MS analysis revealed a higher concentration of Gd in the cerebrum and spinal cord of gadobutrol + LPS-treated rats, while peripheral nerves showed lower concentrations. In addition, there was a significant increase in LDH activity in the hippocampus of the gadobutrol group. EMG responses to electrical stimulation of the sciatic nerve demonstrated a decreased threshold of nociceptive reflexes in the gadobutrol group. Overall, while gadobutrol may be considered safe under normal physiological conditions, the findings suggest that its safety may be compromised under inflammatory conditions.
Keywords:
gadolinium; gadobutrol; GBCA; brain; spinal cord; nerve; pain; ICP-MS; inflammation; lipopolysaccharide 1. Introduction
Gadolinium-based contrast agents (GBCAs) are widely used to enhance imaging in medical procedures such as MRI mainly due to their ability to decrease the longitudinal and transverse relaxation times [1]. While GBCAs are generally considered safe, there is growing concern that the deposition of gadolinium in various tissues of the body including the brain may lead to long-term neurological impairment. Different GBCAs have varying effects due to their distinct chemical structures and properties [2]. Some types of GBCAs may be more likely to deposit gadolinium in the central nervous system compared to others [2]. In fact, studies have shown that linear GBCAs such as gadodiamide and gadopentetate dimeglumine have a higher propensity to deposit gadolinium in the brain compared to macrocyclic GBCAs such as gadoteridol and gadobutrol [2]. This is believed to be due to the fact that linear GBCAs have a lower stability constant than macrocyclic GBCAs and are more likely to release free gadolinium ions that can deposit in neural tissues [2]. Preclinical studies have shown that repeated injections of GBCAs over 20 days increased Gd deposition not only in various areas of the brain, but also in the spinal cord and peripheral nerves [3,4]. The effect of Gd deposition on sensory functions was evident in clinical and preclinical studies that have investigated the long-term consequences of GBCA exposure. In one study, it was shown that Gd deposition following gadodiamide but not gadoterate meglumine administration in rats could induce pain hypersensitivity to mechanical or thermal stimuli [4]. Another study demonstrated that the perinatal administration of gadodiamide or gadoterate dimeglumine led to Gd deposition in various organs of mice mothers and pups [5]. Intravenous administration of gadodiamide or gadoterate meglumine (2 mmol/kg) in pregnant mice leads to behavioral dysfunction and possible developmental neurotoxicity in the pups [6]. The offspring displayed decreased nociceptive threshold, memory deficits, and impaired motor function, particularly following gadodiamide [6]. Clinically, symptoms such as burning, itching, numbness, “pins and needles”, and confusion, which were most likely attributed to the neurotoxic effects of GBCAs, have been reported by patients [7,8]. While these findings suggest that the deposition of Gd in the brain and other tissues may have adverse effects on neurological functions, a study has shown that rats subjected to repeated injections of GBCAs did not exhibit any behavioral deficits [9]. As the current literature presents conflicting results, further studies are needed to confirm whether repeated GBCA administration can impair neurological functions.
Gadobutrol is frequently used in MRIs as a diagnostic tool for patients with multiple sclerosis, patients with brain tumors, or patients with non-tumorigenic lesions in the central nervous system such as white matter disease or a hemorrhage [10]. These patients have to undergo multiple MRIs following diagnosis to evaluate the effectiveness of treatment and monitor disease progression [11]. Given the frequent use of gadobutrol in such patients, it is imperative to thoroughly investigate its safety profile and provide a comprehensive evaluation of repeated gadobutrol injection on the nervous system. Due to gadobutrol’s high kinetic and thermodynamic stability, it is administered intravenously at double the concentration of other types of GBCAs with a dose of 0.1 mmol/kg [10,12]. It is a macrocyclic and non-ionic GBCA that is more commonly known as GadavistTM [13]. Moreover, it has a half-life of 1.33–2.13 h and is 90% eliminated through urine within 12 h after an intravenous injection [12]. Several clinical studies have shown that the rate of adverse reactions such as nausea, dizziness, and headaches following gadobutrol is low [14,15]. In clinical and pre-clinical studies that investigated the effect of serial injections of gadobutrol in the dentate nucleus or globus pallidus, no T1 signal changes were seen in the targeted areas [16]. However, it is undetermined whether other brain structures were examined in these studies.
Up until now, evidence suggests that possible Gd-induced neurotoxicity can be ascribed to different mechanisms acting either singly or in synergy. The first is that Gd can modulate Ca2+ flow across voltage or ligand-gated channels to affect neuronal transmission [17,18]. Gd can also induce apoptosis accompanied by an increased release of lactate dehydrogenase (LDH) and oxidative stress, leading to tissue microdamage in rat cortical neurons [19,20]. This is also supported by a study on neuroblastoma cells in which the addition of GBCAs was shown to reduce cell viability, increase LDH release, and promote an increase in the expression of apoptotic markers [21]. These studies highlight the possible neurotoxic complications associated with the repeated administration of GBCAs in specific patients. Other potential mechanisms by which Gd can exert its effect are through the release of inflammatory interleukins (IL) by monocytes and the induction of NLRP3 (NOD-, LRR-and pyrin domain-containing protein 3)-dependent IL-1β production in macrophages [22,23]. Interestingly, when the macrophages were treated with lipopolysaccharide (LPS), Gd ions were able to modulate the release of different pro- and anti-inflammatory ILs [24]. This interaction between Gd and inflammatory mediators raises questions regarding the toxic action of released Gd3+ on neurons in a setting of a pre-existing inflammatory state. Due to the scarcity of research in this area, it is yet to be determined whether administering gadobutrol to patients with underlying inflammatory conditions increases their risk of neurotoxicity. In this preliminary study, we sought to investigate and compare the impact of repeated gadobutrol administration on the peripheral and central nervous system under physiological and inflammatory conditions.
2. Materials and Methods
2.1. Animals
Twenty-four male Sprague Dawley rats weighing around 150–250 g were used in this study. The rats were housed in a room maintained at a constant temperature (20–22 °C) with a 12-h light/dark cycle. Standard chow and water were provided ad libitum. All experimental procedures were conducted with the approval of the Institutional Animal Care and Use Committee at the American University of Beirut (July 2020).
2.2. Treatment Regimen
Rats were randomly divided into 4 groups: saline group, gadobutrol group, LPS + saline group, and LPS + gadobutrol group (Figure 1). Gadobutrol (2.5 mmol/kg) and saline were administered intraperitoneally and daily over 20 days. For the LPS groups, a single injection of LPS (5 mg/kg) was given intraperitoneally on day 1. One hour after this initial LPS injection, rats received either gadobutrol or saline injection daily for 20 days [25].
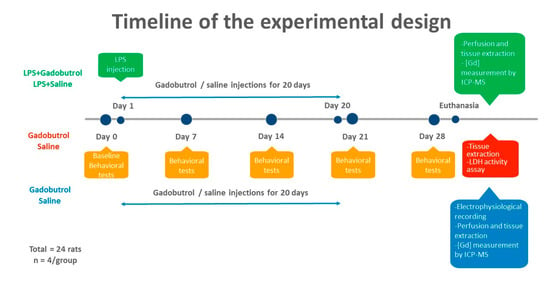
Figure 1.
Timeline of the experiment. Rats were randomly divided into 4 groups: saline group (n = 8), gadobutrol group (n = 8), LPS + saline group (n = 4), and LPS + gadobutrol group (n = 4). Gadobutrol (2.5 mmol/kg) and saline were administered intraperitoneally for 20 days. For the LPS groups, a single injection of LPS was given intraperitoneally at a dosage of 5 mg/kg followed one hour later by gadobutrol or saline injection for 20 days. Behavioral tests were conducted at the baseline and weekly over 4 weeks while electrophysiological assessment was conducted before perfusion. Following tissue collection, gadolinium concentration was measured by ICP-MS and cytotoxicity was evaluated using the lactate dehydrogenase assay.
2.3. Animal Perfusion
Prior to perfusion or surgical procedures, the rats were anesthetized using an intraperitoneal injection of ketamine (80 mg/kg, Ketalar; Panpharma, Luitré, France) and xylazine (10 mg/kg, Xyla; Interchemie, Harju County, Estonia). Following anesthesia, the rats were perfused through the left ventricle with a heparinized saline solution (0.9%) followed by 10% formalin for tissue fixation. The brain, spinal cord, and peripheral nerves were extracted and stored in 4% paraformaldehyde overnight, then in 30% sucrose solution for storage at 4 °C.
2.4. Quantification of Gadolinium Using ICP-MS
To measure the concentration of Gd in the central and peripheral nervous systems, the cerebrum, spinal cord, and peripheral nerves (trigeminal and sciatic nerves) were extracted following the last gadobutrol injection from all groups (n = 4). Then, these extracted tissues were digested at 180 °C for 30 min [26]. Along with the samples, a blank, spiked blank, certified referenced sample, and a matrix spike were also run. The concentration of Gd was measured by using ICP-MS (Agilent 7500ce; Agilent, Waldbronn, Germany).
2.5. LDH Assay
The activity of LDH was measured in the saline group (n = 4) and gadobutrol group (n = 4) one day after the last injection. The rats were anesthetized with isoflurane then decapitated to extract the sciatic nerve and hippocampus. Tissues and solutions were prepared according to the manufacturer’s instructions (QuantiChrom™ Lactate Dehydrogenase Kit, D2DH-100, Bioassay Systems, Hayward, CA, USA). The OD565 was measured using a Microplate Fluorometer/Luminometer Fluoroskan Ascent FL (Thermo Scientific, Waltham, MA, USA) at t = 0 min and at t = 25 min and the LDH activity was subsequently calculated.
2.6. Behavioral Tests
2.6.1. Heat Sensitivity Test
The heat sensitivity test was conducted to assess sensitivity to noxious thermal stimulus. All groups were tested once a week over 4 weeks. To carry out the test, the rats were allowed to habituate to the testing area for one hour. A heat stimulus, at an intensity of 35 infrared units, was then applied to the plantar surface of the hindpaws. Foot withdrawal latency was measured over 5 trials for both paws with a 3-min interval between each trial to avoid paw injury. A cut-off time of 20 s was set to avoid tissue damage.
2.6.2. Beam Walking Test
The beam walking test was performed to assess motor coordination and balance. One day prior to testing, the rats were trained to walk across the beam and familiarize with the apparatus and new height. In order to encourage the rats to cross the beam, they were placed at the same starting point and only moved back to their cage once the finish point was reached. Tests were conducted by recording the time required to cross the beam from one side to another during 3 trials for each rat at the set time points.
2.6.3. Spontaneous Alternation T-Maze Test
To assess spatial working memory, all rats underwent the spontaneous alternation T-maze test. This is a test driven by spontaneous alternation and was carried out according to a previously described protocol [27]. Briefly, the test was conducted by first placing the rat in a start arm and allowing it to choose either the left or right arm. Once a choice is made, the rat is trapped in its chosen arm for 30 s, then returned to the start arm to rechoose an arm. The percentage of successful alternation was calculated for 3 trials.
2.7. Electrophysiological Assessment
For electromyographic recordings, each rat in the gadobutrol (n = 4) and saline (n = 4) groups was placed under general anesthesia (ketamine, 100 mg/kg and xylazine, 10 mg/kg). Compound muscle action potentials (CMAPs) were evoked in the gastrocnemius muscle in response to electrical stimulation of the sciatic nerve at various intensities (1 V, 5 V, and 7 V). Electromyographic (EMG) responses were recorded using a microneedle electrode placed in the belly of the gastrocnemius muscle, while the reference electrode was placed in the Achilles tendon. The signal from the recording electrode was fed into a differential amplifier, filtered, and monitored on an oscilloscope (Tektronix Instruments, Tektronix, Beaverton, OR, USA). The analog signal was sampled in a 1401 data interface (CED 1401, Cambridge, UK) and analyzed using Spike 2 software. The amplitude of the evoked MAPs from peak to peak was calculated and analyzed.
2.8. Statistical Analysis
All results were statistically analyzed using GraphPad Prism 8 (GraphPad Software, Inc., Boston, MA, USA). The data were represented as the average ± standard error of the mean (SEM) and normality was assumed throughout all tests. For the ICP-MS results, the multiple t-tests, corrected using the Holm–Sidak method, were used to analyze the statistical significance between the experimental groups. For the behavioral tests, two-way ANOVA with repeated measures followed by Tukey’s multiple comparison test were used. Multiple t-tests within each group were also carried out to compare the results at different time points with the baseline as a reference point. An unpaired t-test and multiple t-tests corrected using the Holm–Sidak method were used to assess significance in the LDH activity assay and EMG results, respectively. Statistical significance was indicated by a p value less than 0.05.
3. Results
3.1. Quantification of Gadolinium Using ICP-MS
Comparisons of the amount of Gd detected in the central and peripheral neural tissues by ICP-MS are shown in Figure 2. In the cerebrum, the concentration of Gd was significantly higher in the LPS + gadobutrol group (2.315 ± 0.4671) compared to the gadobutrol (0.4465 ± 0.0394 [p = 0.007]) and the saline groups (0.0598 ± 0.0011 [p = 0.03]) (Figure 2A). In the spinal cord, the LPS + gadobutrol group (0.7995 ± 0.1023) showed a much higher Gd concentration compared to the saline group (0.0918 ± 0.0015 [p < 0.001]) and the gadobutrol group (0.3683 ± 0.0265 [p = 0.006]) (Figure 2B). Similarly, in the peripheral nerves, the concentration of Gd was significantly higher in the gadobutrol group (9.253 ± 1.069 [p < 0.001]) and the LPS + gadobutrol group (5.345 ± 1.434 [p = 0.01]) compared to the saline group (0.4084 ± 0.0401) (Figure 2C). However, the difference in Gd concentration between the gadobutrol (9.253 ± 1.069) and LPS + gadobutrol groups (5.345 ± 1.434 [p = 0.07]) was not significant.
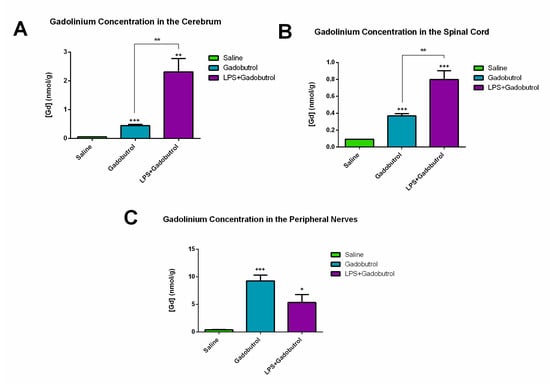
Figure 2.
Increased gadolinium concentration in the central and peripheral nervous systems. (A) Concentration of gadolinium in the cerebrum (area between the optic chiasm and the midbrain) of the saline group, gadobutrol group, and LPS + gadobutrol group. (B) Concentration of gadolinium in the spinal cord of the saline group, gadobutrol group, and LPS + gadobutrol group. (C) Concentration of gadolinium in the peripheral nerves (sciatic and trigeminal nerves) of the saline group, gadobutrol group, and LPS + gadobutrol group. Data are represented as the mean ± SEM. Multiple t-tests corrected using the Holm–Sidak method were used to determine statistical significance. Statistical significance compared to the saline group is indicated by +: p < 0.05; ++: p < 0.01; +++: p < 0.001 while significance compared to the gadobutrol group is indicated by **: p < 0.01.
3.2. LDH Assay
Cytotoxic changes in the hippocampus and sciatic nerves were assessed by measuring the activity of LDH, a biomarker used to determine cellular membrane integrity and permeability. LDH released in the hippocampus of the gadobutrol group (1661 ± 75.46) was found to be higher compared to the saline group (1260 ± 59.90 [p = 0.006]) (Figure 3A), an indicator of the loss of plasma membrane integrity. However, the LDH activity in the sciatic nerves did not show evidence of a difference between the two groups (gadobutrol: 209.7 ± 21.50; saline: 234.8 ± 42.26) (Figure 3B).
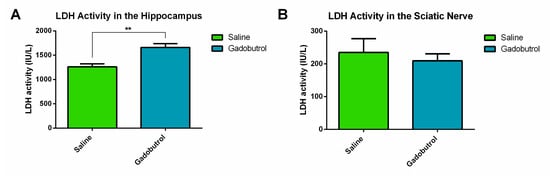
Figure 3.
Cytotoxic effects of gadobutrol. (A) Lactate dehydrogenase activity in the hippocampus of the saline group and the gadobutrol group. (B) Lactate dehydrogenase activity in the sciatic nerve of the saline group and the gadobutrol group. Data are represented as the mean ± SEM. For statistical analysis, an unpaired t-test was carried out (**: p < 0.01).
3.3. Heat Sensitivity Test
Heat sensitivity tests were conducted on all rats once a week over a period of 4 weeks to assess their response to a noxious heat stimulus (Figure 4A,B). A reduction in paw withdrawal latency indicates an increased sensitivity to noxious thermal stimulation. For the right paw, the LPS + gadobutrol group exhibited a significantly lower withdrawal latency than the gadobutrol group at week 4 (LPS + gadobutrol 4.1 ± 0.4975 [p = 0.001]; gadobutrol 6.61 ± 0.8103). Compared to the baseline, the withdrawal latency of the LPS + saline and LPS + gadobutrol groups declined at week 2 (LPS + saline 5.025 ± 0.4698 [p = 0.004]; LPS + gadobutrol 5.135 ± 0.11 [p = 0.004]). On the other hand, the withdrawal latency of only the LPS + gadobutrol group decreased compared to the baseline at week 3 (LPS + gadobutrol 5.18 ± 0.4976 [p = 0.006]) and week 4 (LPS + gadobutrol 4.1 ± 0.4975 [p = 0.002]).
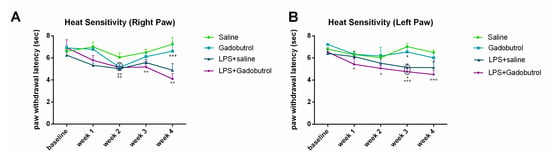
Figure 4.
Effect of gadobutrol administration on heat sensitivity. (A) Effect of gadobutrol administration on heat sensitivity in the right hindpaw. Multiple t-tests corrected using the Holm–Sidak method were used to compare the mean at each time point with the baseline (++: p < 0.01). Two-way ANOVA test followed by Tukey’s test were used to compare the means at each time point between groups. At week 4, statistical significance was present between the LPS + gadobutrol and gadobutrol groups (***: p < 0.001). (B) Effect of gadobutrol administration on heat sensitivity in the left hindpaw. Data are represented as mean ± SEM. Multiple t-tests corrected using the Holm–Sidak method were used to compare the mean at each time point with the baseline (+: p < 0.05; +++: p < 0.001). Two-way ANOVA test followed by Tukey’s test were used to compare the means at each time point between groups. At weeks 3 and 4, statistical significance was obtained between the LPS + gadobutrol and gadobutrol groups (*: p < 0.05).
For the left paw, the LPS + gadobutrol group exhibited lower withdrawal latency than the gadobutrol group at week 3 (LPS + gadobutrol 4.755 ± 0.3163 [p = 0.02]; gadobutrol 6.545 ± 0.538) and week 4 (LPS + gadobutrol 4.505 ± 0.3815 [p = 0.049], gadobutrol 6.01 ± 0.5204). Compared to the baseline, the LPS + gadobutrol group exhibited a significant decline in withdrawal latency for the duration of the experiment (week 1: LPS + gadobutrol 5.425 ± 0.5438 [p = 0.03]; week 2: LPS + gadobutrol 5.055 ± 0.2766 [p = 0.02]; week 3: LPS + gadobutrol 4.755 ± 0.3163 [p < 0.001]; week 4: LPS + gadobutrol 4.505 ± 0.3815 [p < 0.001]). The LPS + saline showed a drop in withdrawal latency compared to the baseline at week 3 (LPS + saline 5.14 ± 0.3275 [p = 0.014]) only.
3.4. Beam Walking Test
Beam walking test was conducted to assess balance and coordination (Figure 5A). Statistical analysis between groups showed that only the LPS + gadobutrol group (2.697 ± 0.3511) had a longer beam crossing duration than the gadobutrol group (1.823 ± 0.0974 [p = 0.049]) during week 3. On the other hand, compared to the baseline, the LPS and LPS + gadobutrol groups required a longer time to cross the beam when tested at week 3 (LPS + saline 2.393 ± 0.1894 [p = 0.008], LPS + gadobutrol 2.697 ± 0.3511, p = 0.001), and week 4 (LPS + saline 2.578 ± 0.2296 [p = 0.004], LPS + gadobutrol 2.795 ± 0.2475, [p = 0.001]).
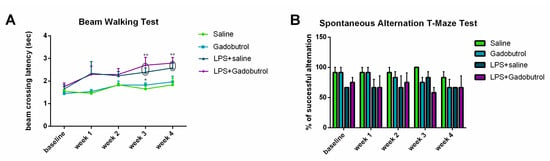
Figure 5.
Effect of gadobutrol administration on motor and cognitive abilities. (A) Effect of gadobutrol administration on motor performance in the beam walking test. (B) Effect of gadobutrol administration on spatial working memory in the T-maze test. Data are represented as the mean ± SEM. Multiple t-tests corrected using the Holm–Sidak method were used to compare the mean at each time point with the baseline (++: p < 0.01). Two-way ANOVA test followed by Tukey’s test were used to compare the means at each time point between groups. At week 3, statistical significance was present between the LPS + gadobutrol and gadobutrol groups (*: p < 0.05).
3.5. Spontaneous Alternation T-Maze Test
In order to evaluate the cognitive behavior of the tested groups, the spontaneous alternation T-maze test was performed once a week for 4 weeks (Figure 5B). Compared to the baseline values, no evidence of a significant difference in working memory was detected between the means of the different groups computed at each time point (p > 0.05).
3.6. Electrophysiological Assessment
Electromyographic recordings from the gastrocnemius muscle, in response to electrical stimulation of the sciatic nerve, did not show evidence of a difference between the mean amplitude of the evoked CMAPs of the saline and gadobutrol groups (at 1 V: gadobutrol (0.6887 ± 0.1084 [p = 0.27]); at 5 V: saline (1.49 ± 0.4264), gadobutrol (1.332 ± 0.5825 [p = 0.8]); at 7 V: saline (2.492 ± 0.4875), gadobutrol (1.523 ± 0.5929 [p = 0.13])) (Figure 6). However, the gadobutrol group exhibited a lower threshold for muscle activation by responding to a stimulating voltage of 1 V (0.6887 ± 0.1084) while the saline group required a higher voltage of 5 V for activation (1.49 ± 0.4264).
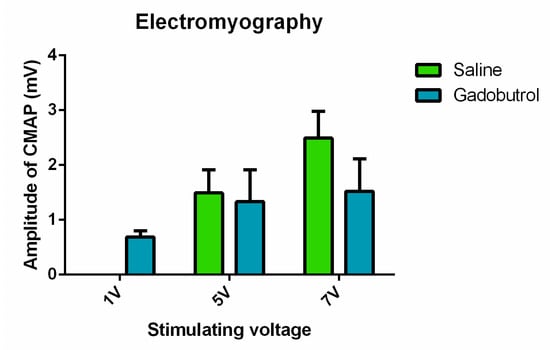
Figure 6.
Electrophysiological assessment of the effect of gadobutrol. Amplitude of the evoked compound muscle action potentials recorded at a voltage of 1 V, 5 V, and 7 V in the saline and gadobutrol groups. Data are represented as the mean ± SEM. For statistical analysis, multiple t-tests were carried out at each voltage.
4. Discussion
This preliminary study is the first to comprehensively evaluate the potential neurotoxic effects of gadobutrol on sensorimotor and cognitive functions using behavioral, molecular, and electrophysiological tests under normal and inflammatory conditions. Our findings showed that repeated intraperitoneal administration of gadobutrol (2.5 mmol/kg), under physiological conditions, resulted in increased Gd concentration in the peripheral and central nervous tissues without any manifestations of cognitive, motor, or sensory impairment. However, when neuronal homeostasis is disrupted by inflammation, the same experimental paradigm results in a significant increase in Gd concentration in the cerebrum (p = 0.007) and the spinal cord (p = 0.006), accompanied by an increase in pain sensitivity to noxious heat stimulation (week 4 p < 0.001).
In the absence of inflammation, the results of the sensorimotor and cognitive behavioral tests are consistent with previous studies demonstrating the safe use of gadobutrol [14,15,16]. Indeed, a recent study has shown that the intravenous administration of various GBCAs over 4 weeks (2.5 mmol/kg) in rats did not lead to histopathological or behavioral evidence of neurotoxicity [9]. The absence of hindpaw hypersensitivity to noxious thermal stimulation following gadobutrol injection indicates that increased Gd concentration in the spinal cord and peripheral nerves, as confirmed by ICP-MS, did not trigger any chemical changes that precipitate peripheral or central sensitization [4]. Similarly, our results showed that gadobutrol, like gadodiamide and gadoterate meglumine, caused no spatial working memory impairment in rats, despite evidence of cytotoxic changes in the hippocampus as indicated by the increase in LDH activity [4]. This outcome is in line with earlier research showing that damage of the hippocampus alone is insufficient to produce global memory impairment [28]. While these results indicate that gadobutrol has penetrated the blood–brain barrier and deposited in the hippocampus, it is difficult to speculate whether Gd is the direct trigger of the cytotoxic changes observed in the hippocampus. A long-term follow-up study is needed to determine whether tissue damage in this anatomic region, as indicated by an increase in LDH, is transient or will worsen over time and cause cognitive impairment.
Evidently, the concentration of Gd deposited in tissues has been shown to vary depending on the type of GBCA used. For example, results from a previous study demonstrated that intraperitoneal injection of gadobutrol led to lower Gd deposition in the cerebrum and spinal cord by comparison with gadodiamide and gadoterate meglumine [4]. Interestingly, however, gadobutrol resulted in greater Gd concentration in the peripheral nerves than gadoterate meglumine, but was still less than that of gadodiamide [4]. Although increased Gd concentration in the peripheral nerves (trigeminal and sciatic) of gadobutrol-treated rats did not trigger any changes in peripheral sensorimotor function or LDH activity, the EMG recordings clearly indicated that these rats exhibited a decrease in the threshold of nociceptive reflexes compared to the control group, possibly due to the modulatory effect of Gd on voltage-gated channels [17]. Apparently, the observed decrease in threshold of CMAP was not adequate enough to lead to alterations in sensorimotor behavior. Another possibility is that the Gd concentration measured in peripheral nerves was more concentrated in the trigeminal rather than sciatic nerve given the absence of peripheral dysfunction but high Gd concentration in the nerves, as analyzed by ICP-MS. As such, in the peripheral nerves, Gd may be preferentially deposited in cranial nerves rather than spinal cord nerves, which is partly supported by the greater concentration of Gd deposited in the cerebrum compared to the spinal cord. Indeed, Gd has been detected in the trigeminal and oculomotor cranial nerves after the introduction of gadobutrol into the interstitial fluid of the brainstem [29]. It has been proposed that Gd can move across the basement membrane of the brainstem capillaries into the endoneurium of the trigeminal nerve [29].
Our experimental results indicate that the inflammatory changes induced by LPS can possibly make the central nervous system more vulnerable to retaining a higher concentration of Gd following gadobutrol exposure, potentially by affecting the clearance rate or the permeability of the blood–brain barrier. The increase in Gd concentration following inflammation is in line with previous studies [24,30,31]. LPS, an inflammatory agent, is known to activate a network of pro-inflammatory factors (tumor necrosis factor α and IL-18) and perturb CSF-brain homeostasis [25,32,33,34]. The significant increase in the concentration of Gd in the brain and spinal cord of the LPS + gadobutrol group could thus be due to the disruption of the blood–brain barrier or blood–spinal cord barrier mediated by inflammatory and neuroimmune mechanisms [35,36]. The possible leakage of intravenous GBCAs into the subarachnoid space in patients carries the risk of neurotoxic complications such as focal seizures [37]. Therefore, the potential presence of GBCAs in the CSF can cause neurologic deficits. On the other hand, the lesser increase in Gd concentration in the peripheral nerves of the LPS + gadobutrol group prompts the speculation that LPS-induced neuroinflammation has a less disruptive impact on the blood–nerve barrier, and, as a result, Gd is siphoned off into the central nervous system. Previous studies have indeed shown that the degree of barrier disruption may vary based on the LPS dosage, regional impact, and released neurochemical mediators [38]. It is important to note that the sensory sensitivity observed in the LPS + gadobutrol group can be attributed either to LPS-induced inflammation or gadobutrol toxicity. Although our preliminary data are not sufficient to identify the causative factor, the impact of the synergistic effect of LPS-induced inflammation and gadobutrol retention on sensory and motor functions is evident. Taken together, our results suggest that a cautious approach may be warranted when using gadobutrol in patients with inflammatory conditions, as its potential neurotoxicity may be exacerbated under these circumstances.
This study had some limitations. First, the sample size was relatively small; however, because of the toxic nature of GBCAs, the number of animals was reduced to the minimum needed to achieve the objectives of the study. A larger sample size may enhance the statistical power of our study and enable us to detect a significant difference between the groups. Second, the T-maze test used to evaluate cognitive functions is a non-automated, hand-run task that requires lots of handling by the experimenter, which might have crucially impacted the behavioral responses. Therefore, additional cognitive tests are recommended to confirm the insignificant impact of gadobutrol on cognitive functions. Additionally, since gadobutrol was the only available GBCA at the time of the experiment, it was not possible to compare the results of gadobutrol with other types of GBCAs. Another limitation of this study was that the levels of pro-inflammatory and anti-inflammatory cytokines prior to and following LPS injection were not measured to confirm the presence of inflammatory mediators; regardless, the inflammatory response in the LPS-treated animals was evidenced by increased pain behavior, general sickness, and general behavioral depression [39].
5. Conclusions
Our preliminary findings suggest that repeated administration of gadobutrol results in an increased concentration of Gd in both the peripheral and central nervous tissues, leading to cellular damage, as indicated by a high hippocampal LDH activity. Furthermore, under inflammatory conditions, repeated injections of gadobutrol result in increased heat sensitivity and Gd concentration in the cerebrum and spinal cord, indicating a possible synergistic neurotoxic effect of inflammation and gadobutrol. These results highlight the potential role of inflammation in the neurotoxicity of GBCAs and call for further investigation into the role of inflammatory mediators in exacerbating the neurotoxic effects of retained Gd. These findings are particularly relevant for patients with inflammatory diseases such as multiple sclerosis or arthritis that require multiple MRI exams. It is clear that there is a need for additional research to fully understand the mechanisms of action of gadobutrol as it appears to employ different pathways that can lead to both central and peripheral neurological manifestations.
Author Contributions
M.J.: Data curation; Formal analysis; Roles/Writing—original draft; W.A.-K.: Conceptualization; Project administration; Resources; Software; Supervision; Validation; Visualization; Writing—review & editing. N.L.: Conceptualization; Funding acquisition; Investigation; Methodology; Project administration; Resources; Software; Supervision; Validation; Visualization; Writing—review & editing. All authors have read and agreed to the published version of the manuscript.
Funding
This work was partly funded by the Medical Practice Plan (MPP-WAK18, MPP-NL20) at AUB-FM and the Diana Tamari Sabbagh Scholars Program (DTSSP, Project 9216).
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Animal Care and Use Committee (IACUC) at the American University of Beirut (July 2020).
Informed Consent Statement
Not applicable.
Data Availability Statement
All data can be provided upon reasonable request.
Acknowledgments
The authors would like to express their gratitude to the Diana Tamari Sabbagh Scholars Program (DTSSP) for their generous funding of the present work. Also, the authors would like to thank Jimmy Asroui from the Environment Core Laboratory at the American University of Beirut for the inductively coupled plasma-mass spectrometry analysis.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Botta, M. Second coordination sphere water molecules and relaxivity of gadolinium (III) complexes: Implications for MRI contrast agents. Eur. J. Inorg. Chem. 2000, 2000, 399–407. [Google Scholar] [CrossRef]
- Davies, J.; Siebenhandl-Wolff, P.; Tranquart, F.; Jones, P.; Evans, P. Gadolinium: Pharmacokinetics and toxicity in humans and laboratory animals following contrast agent administration. Arch. Toxicol. 2022, 96, 403–429. [Google Scholar] [CrossRef] [PubMed]
- McDonald, R.J.; McDonald, J.S.; Dai, D.; Schroeder, D.; Jentoft, M.E.; Murray, D.L.; Kadirvel, R.; Eckel, L.J.; Kallmes, D.F. Comparison of gadolinium concentrations within multiple rat organs after intravenous administration of linear versus macrocyclic gadolinium chelates. Radiology 2017, 285, 536–545. [Google Scholar]
- Alkhunizi, S.M.; Fakhoury, M.; Abou-Kheir, W.; Lawand, N. Gadolinium Retention in the Central and Peripheral Nervous System: Implications for Pain, Cognition, and Neurogenesis. Radiology 2020, 297, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Erdene, K.; Nakajima, T.; Kameo, S.; Khairinisa, M.A.; Lamid-Ochir, O.; Tumenjargal, A.; Koibuchi, N.; Koyama, H.; Tsushima, Y. Organ retention of gadolinium in mother and pup mice: Effect of pregnancy and type of gadolinium-based contrast agents. Jpn. J. Radiol. 2017, 35, 568–573. [Google Scholar] [CrossRef]
- Khairinisa, M.A.; Takatsuru, Y.; Amano, I.; Erdene, K.; Nakajima, T.; Kameo, S.; Koyama, H.; Tsushima, Y.; Koibuchi, N. The Effect of Perinatal Gadolinium-Based Contrast Agents on Adult Mice Behavior. Investig. Radiol. 2018, 53, 110–118. [Google Scholar] [CrossRef]
- Saupe, N.; Zanetti, M.; Pfirrmann, C.W.; Wels, T.; Schwenke, C.; Hodler, J. Pain and other side effects after MR arthrography: Prospective evaluation in 1085 patients. Radiology 2009, 250, 830–838. [Google Scholar] [CrossRef]
- Semelka, R.C.; Commander, C.W.; Jay, M.; Burke, L.M.; Ramalho, M. Presumed gadolinium toxicity in subjects with normal renal function: A report of 4 cases. Investig. Radiol. 2016, 51, 661–665. [Google Scholar] [CrossRef]
- Ayers-Ringler, J.; McDonald, J.S.; Connors, M.A.; Fisher, C.R.; Han, S.; Jakaitis, D.R.; Scherer, B.; Tutor, G.; Wininger, K.M.; Dai, D.; et al. Neurologic Effects of Gadolinium Retention in the Brain after Gadolinium-based Contrast Agent Administration. Radiology 2022, 302, 676–683. [Google Scholar] [CrossRef]
- Scott, L.J. Gadobutrol: A Review in Contrast-Enhanced MRI and MRA. Clin. Drug Investig. 2018, 38, 773–784. [Google Scholar] [CrossRef]
- Kaunzner, U.W.; Gauthier, S.A. MRI in the assessment and monitoring of multiple sclerosis: An update on best practice. Ther. Adv. Neurol. Disord. 2017, 10, 247–261. [Google Scholar] [CrossRef] [PubMed]
- FDA. Gadavist (Gadobutrol) Injection. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2011/201277s000lbl.pdf (accessed on 1 August 2021).
- Port, M.; Idée, J.-M.; Medina, C.; Robic, C.; Sabatou, M.; Corot, C. Efficiency, thermodynamic and kinetic stability of marketed gadolinium chelates and their possible clinical consequences: A critical review. Biometals 2008, 21, 469–490. [Google Scholar] [CrossRef] [PubMed]
- Glutig, K.; Hahn, G.; Kuvvetli, P.; Endrikat, J. Safety of gadobutrol: Results of a non-interventional study of 3710 patients, including 404 children. Acta Radiol. 2019, 60, 873–879. [Google Scholar] [CrossRef] [PubMed]
- Endrikat, J.; Schwenke, C.; Prince, M. Gadobutrol for contrast-enhanced magnetic resonance imaging in elderly patients: Review of the safety profile from clinical trial, post-marketing surveillance, and pharmacovigilance data. Clin. Radiol. 2015, 70, 743–751. [Google Scholar] [CrossRef]
- Ozturk, K.; Nascene, D. Effect of at least 10 serial gadobutrol administrations on brain signal intensity ratios on T1-weighted MRI in children: A matched case-control study. Am. J. Roentgenol. 2021, 217, 753–760. [Google Scholar] [CrossRef] [PubMed]
- Mlinar, B.; Enyeart, J. Block of current through T-type calcium channels by trivalent metal cations and nickel in neural rat and human cells. J. Physiol. 1993, 469, 639–652. [Google Scholar] [CrossRef]
- Green, C.; Jost, G.; Frenzel, T.; Boyken, J.; Schwenke, C.; Pietsch, H. The Effect of Gadolinium-Based Contrast Agents on Longitudinal Changes of Magnetic Resonance Imaging Signal Intensities and Relaxation Times in the Aging Rat Brain. Investig. Radiol. 2022, 57, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Xia, Q.; Yuan, L.; Yang, X.; Wang, K. Impaired mitochondrial function and oxidative stress in rat cortical neurons: Implications for gadolinium-induced neurotoxicity. Neurotoxicology 2010, 31, 391–398. [Google Scholar] [CrossRef]
- Xia, Q.; Feng, X.; Huang, H.; Du, L.; Yang, X.; Wang, K. Gadolinium-induced oxidative stress triggers endoplasmic reticulum stress in rat cortical neurons. J. Neurochem. 2011, 117, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Erdoğan, M.A.; Apaydin, M.; Armagan, G.; Taskiran, D. Evaluation of toxicity of gadolinium-based contrast agents on neuronal cells. Acta Radiol. 2021, 62, 206–214. [Google Scholar] [CrossRef]
- Wermuth, P.J.; Jimenez, S.A. Induction of a type I interferon signature in normal human monocytes by gadolinium-based contrast agents: Comparison of linear and macrocyclic agents. Clin. Exp. Immunol. 2014, 175, 113–125. [Google Scholar] [CrossRef]
- Schmidt-Lauber, C.; Bossaller, L.; Abujudeh, H.H.; Vladimer, G.I.; Christ, A.; Fitzgerald, K.A.; Latz, E.; Gravallese, E.M.; Marshak-Rothstein, A.; Kay, J. Gadolinium-based compounds induce NLRP3-dependent IL-1β production and peritoneal inflammation. Ann. Rheum. Dis. 2015, 74, 2062–2069. [Google Scholar] [CrossRef]
- Weng, T.-I.; Chen, H.J.; Lu, C.-W.; Ho, Y.-C.; Wu, J.-L.; Liu, S.-H.; Hsiao, J.-K. Exposure of macrophages to low-dose gadolinium-based contrast medium: Impact on oxidative stress and cytokines production. Contrast Media Mol. Imaging 2018, 2018, 3535769. [Google Scholar] [CrossRef] [PubMed]
- Bossù, P.; Cutuli, D.; Palladino, I.; Caporali, P.; Angelucci, F.; Laricchiuta, D.; Gelfo, F.; De Bartolo, P.; Caltagirone, C.; Petrosini, L. A single intraperitoneal injection of endotoxin in rats induces long-lasting modifications in behavior and brain protein levels of TNF-α and IL-18. J. Neuroinflammation 2012, 9, 101. [Google Scholar] [CrossRef] [PubMed]
- Muller, E.I.; Souza, J.P.; Muller, C.C.; Muller, A.L.H.; Mello, P.A.; Bizzi, C.A. Microwave-assisted wet digestion with H2O2 at high temperature and pressure using single reaction chamber for elemental determination in milk powder by ICP-OES and ICP-MS. Talanta 2016, 156–157, 232–238. [Google Scholar] [CrossRef]
- Deacon, R.M.; Rawlins, J.N.P. T-maze alternation in the rodent. Nat. Protoc. 2006, 1, 7. [Google Scholar] [CrossRef]
- Aggleton, J.P.; Hunt, P.; Rawlins, J. The effects of hippocampal lesions upon spatial and non-spatial tests of working memory. Behav. Brain Res. 1986, 19, 133–146. [Google Scholar] [CrossRef]
- Varatharaj, A.; Carare, R.O.; Weller, R.O.; Gawne-Cain, M.; Galea, I. Gadolinium enhancement of cranial nerves: Implications for interstitial fluid drainage from brainstem into cranial nerves in humans. Proc. Natl. Acad. Sci. USA 2021, 118, e2106331118. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Hesse, B.; Roman, M.; Stier, D.; Castillo-Michel, H.; Cotte, M.; Suuronen, J.-P.; Lagrange, A.; Radbruch, H.; Paul, F. Increased retention of gadolinium in the inflamed brain after repeated administration of gadopentetate dimeglumine: A proof-of-concept study in mice combining ICP-MS and micro–and Nano–SR-XRF. Investig. Radiol. 2019, 54, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Anderhalten, L.; Silva, R.V.; Morr, A.; Wang, S.; Smorodchenko, A.; Saatz, J.; Traub, H.; Mueller, S.; Boehm-Sturm, P.; Rodriguez-Sillke, Y. Different impact of gadopentetate and gadobutrol on inflammation-promoted retention and toxicity of gadolinium within the mouse brain. Investig. Radiol. 2022, 57, 677–688. [Google Scholar] [CrossRef]
- Fu, H.Q.; Yang, T.; Xiao, W.; Fan, L.; Wu, Y.; Terrando, N.; Wang, T.L. Prolonged neuroinflammation after lipopolysaccharide exposure in aged rats. PLoS ONE 2014, 9, e106331. [Google Scholar] [CrossRef]
- Qin, L.; Wu, X.; Block, M.L.; Liu, Y.; Breese, G.R.; Hong, J.S.; Knapp, D.J.; Crews, F.T. Systemic LPS causes chronic neuroinflammation and progressive neurodegeneration. Glia 2007, 55, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Manouchehrian, O.; Ramos, M.; Bachiller, S.; Lundgaard, I.; Deierborg, T. Acute systemic LPS-exposure impairs perivascular CSF distribution in mice. J. Neuroinflammation 2021, 18, 34. [Google Scholar] [CrossRef] [PubMed]
- Galea, I. The blood–brain barrier in systemic infection and inflammation. Cell. Mol. Immunol. 2021, 18, 2489–2501. [Google Scholar] [CrossRef] [PubMed]
- Montague-Cardoso, K.; Malcangio, M. Changes in blood–spinal cord barrier permeability and neuroimmune interactions in the underlying mechanisms of chronic pain. Pain Rep. 2021, 6, e879. [Google Scholar] [CrossRef]
- Lauer, M.; Lauer, A.; You, S.J.; Kluge, S.; Hattingen, E.; Harter, P.N.; Senft, C.; Wagner, M.; Voss, M. Neurotoxicity of subarachnoid Gd-based contrast agent accumulation: A potential complication of intraoperative MRI? Neurosurg. Focus 2021, 50, E12. [Google Scholar] [CrossRef]
- Banks, W.A.; Gray, A.M.; Erickson, M.A.; Salameh, T.S.; Damodarasamy, M.; Sheibani, N.; Meabon, J.S.; Wing, E.E.; Morofuji, Y.; Cook, D.G. Lipopolysaccharide-induced blood-brain barrier disruption: Roles of cyclooxygenase, oxidative stress, neuroinflammation, and elements of the neurovascular unit. J. Neuroinflammation 2015, 12, 223. [Google Scholar] [CrossRef]
- Bassi, G.S.; Kanashiro, A.; Santin, F.M.; de Souza, G.E.; Nobre, M.J.; Coimbra, N.C. Lipopolysaccharide-induced sickness behaviour evaluated in different models of anxiety and innate fear in rats. Basic Clin. Pharmacol. Toxicol. 2012, 110, 359–369. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).