
A Comparison of Food and Non-Food Enrichment with Zoo-Housed African Lions (Panthera leo)




Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects and Housing
2.2. Materials
2.3. Design and Procedure
2.4. Statistical Analyses
3. Results
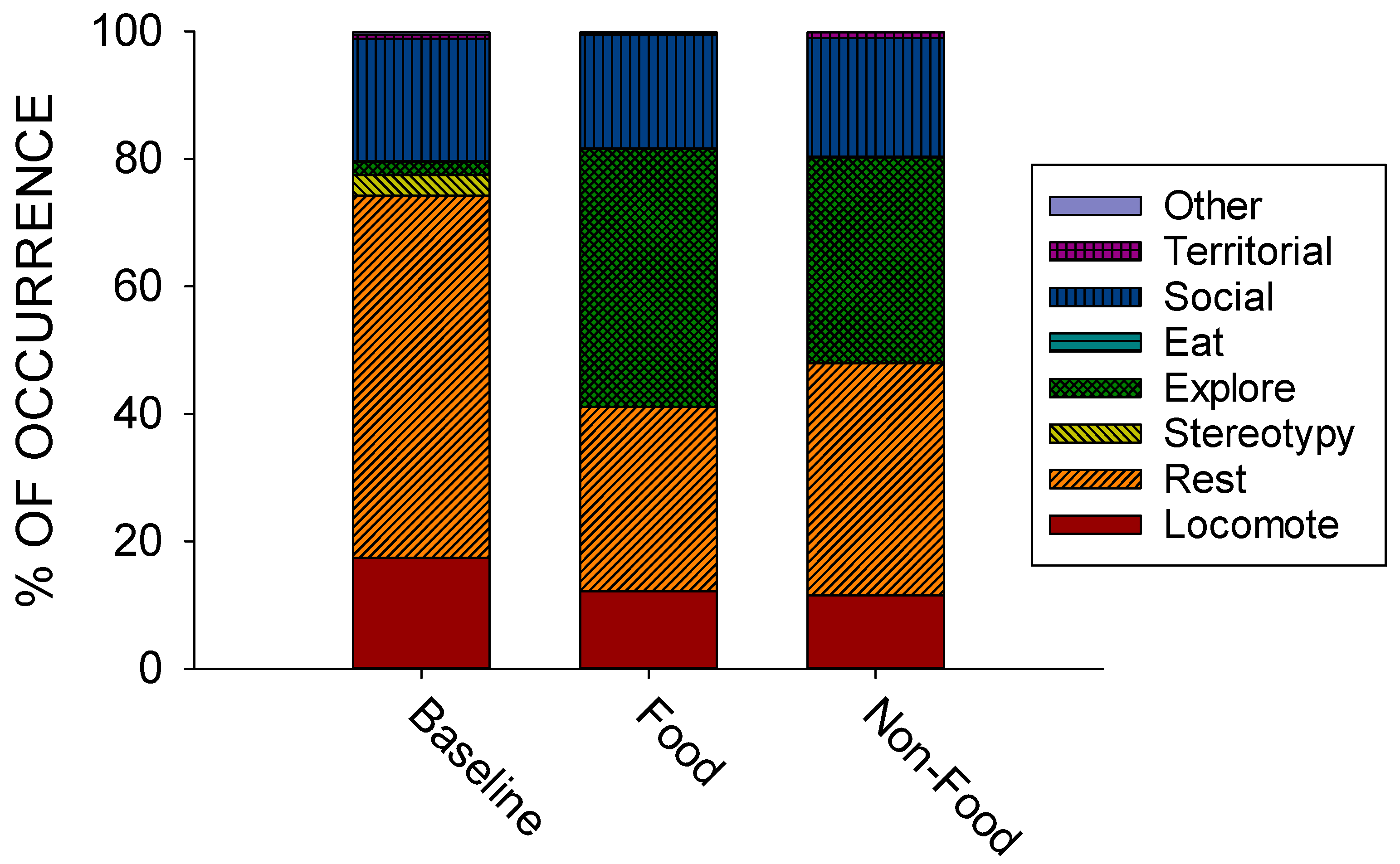
3.1. Significant Differences Between Conditions
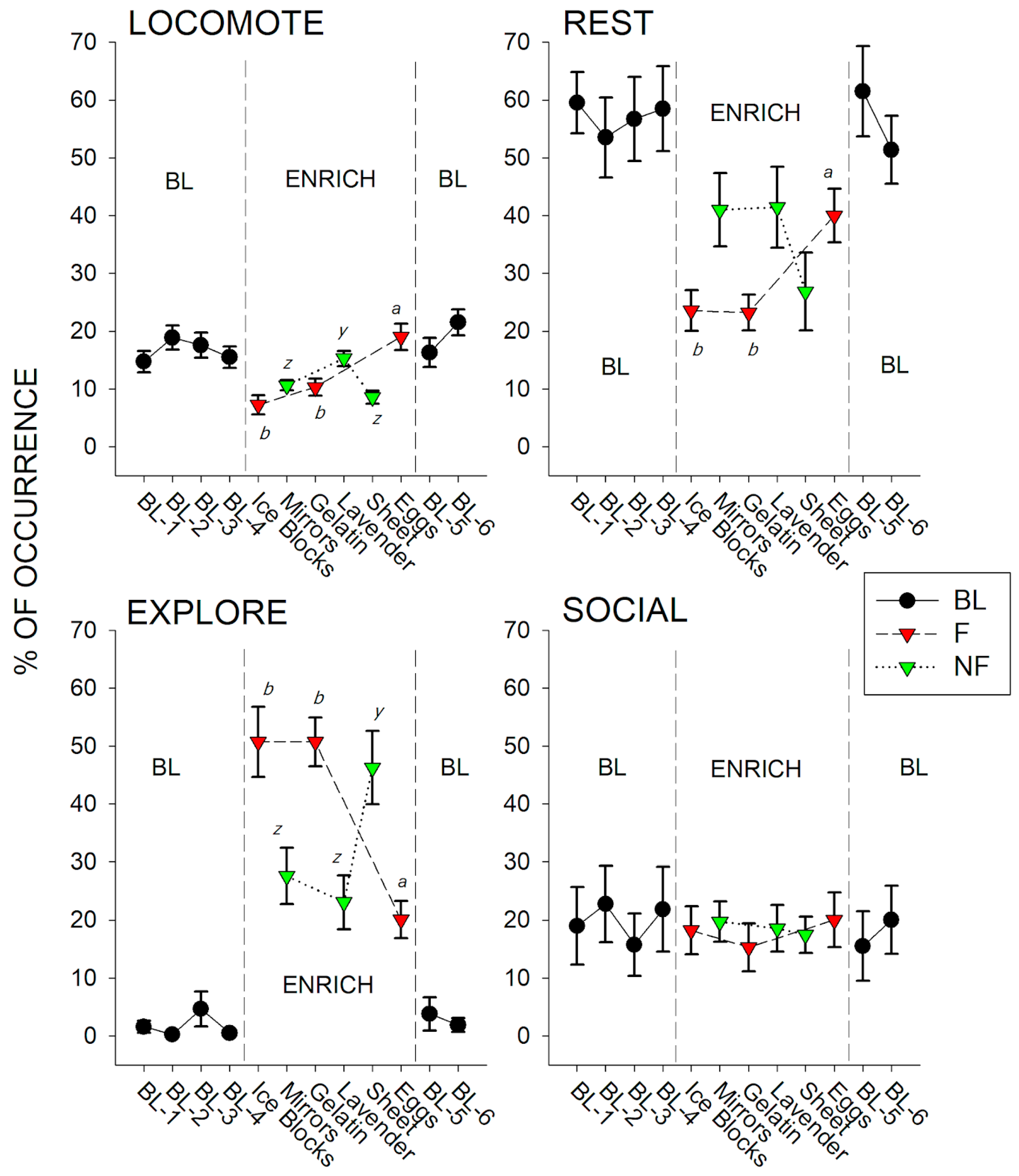
3.2. Significant Differences Between Enrichment Types
3.2.1. Enrichment Type and Locomote
3.2.2. Enrichment Type and Rest
3.2.3. Enrichment Type and Explore
3.2.4. Enrichment Type and Territorial
4. Discussion
4.1. Activity
4.2. Exploration
4.3. Stereotypic Pacing
4.4. Social and Territorial
4.5. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mendyk, R.W.; Augustine, L. Controlled deprivation and enrichment. In Health and Welfare of Captive Reptiles; Springer: Berlin/Heidelberg, Germany, 2023; pp. 323–355. [Google Scholar]
- Clubb, R.; Mason, G.J. Natural behavioural biology as a risk factor in carnivore welfare: How analysing species differences could help zoos improve enclosures. Appl. Anim. Behav. Sci. 2007, 102, 303–328. [Google Scholar] [CrossRef]
- Young, T.; Finegan, E.; Brown, R.D. Effects of summer microclimates on behavior of lions and tigers in zoos. Int. J. Biometeorol. 2013, 57, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Powell, D.M. Preliminary evaluation of environmental enrichment techniques for African lions (Panthera leo). Anim. Welf. 1995, 4, 361–370. [Google Scholar] [CrossRef]
- Hosey, G.; Melfi, V.; Pankhurst, S. Zoo Animals: Behaviour, Management, and Welfare; Oxford University Press: New York, NY, USA, 2013. [Google Scholar]
- Godinez, A.M.; Fernandez, E.J. What is the zoo experience? How zoos impact a visitor’s behaviors, perceptions, and conservation efforts. Front. Psychol. 2019, 10, 469377. [Google Scholar] [CrossRef]
- Mellor, D.J.; Beausoleil, N.J.; Littlewood, K.E.; McLean, A.N.; McGreevy, P.D.; Jones, B.; Wilkins, C. The 2020 five domains model: Including human–animal interactions in assessments of animal welfare. Animals 2020, 10, 1870. [Google Scholar] [CrossRef]
- Mellen, J.; Sevenich MacPhee, M. Philosophy of environmental enrichment: Past, present, and future. Zoo Biol. 2001, 20, 211–226. [Google Scholar] [CrossRef]
- Poole, T.B. Meeting a mammal’s psychological needs: Basic principles. In Second Nature: Environmental Enrichment for Captive Animals; Smithsonian Institution Press: Washington, DC, USA, 1998. [Google Scholar]
- Brando, S.; Buchanan-Smith, H.M. The 24/7 approach to promoting optimal welfare for captive wild animals. Behav. Process. 2018, 156, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Brereton, J.; Rose, P. An evaluation of the role of ‘biological evidence’ in zoo and aquarium enrichment practices. Anim. Welf. 2022, 31, 13–26. [Google Scholar] [CrossRef]
- Hoy, J.M.; Murray, P.J.; Tribe, A. Thirty years later: Enrichment practices for captive mammals. Zoo Biol. 2010, 29, 303–316. [Google Scholar] [CrossRef]
- Swaisgood, R.R.; Shepherdson, D.J. Scientific approaches to enrichment and stereotypies in zoo animals: What’s been done and where should we go next? Zoo Biol. 2005, 24, 499–518. [Google Scholar] [CrossRef]
- Izzo, G.N.; Bashaw, M.J.; Campbell, J.B. Enrichment and individual differences affect welfare indicators in squirrel monkeys (Saimiri sciureus). J. Comp. Psychol. 2011, 125, 347. [Google Scholar] [CrossRef] [PubMed]
- Browning, H. The natural behavior debate: Two conceptions of animal welfare. J. Appl. Anim. Welf. Sci. 2020, 23, 325–337. [Google Scholar] [PubMed]
- Mishra, A.K.; Guru, B.C.; Patnaik, A.K. Effect of feeding enrichment on behaviour of captive tigers. Indian Zoo Year Book 2013, 7, 124–133. [Google Scholar]
- Skibiel, A.L.; Trevino, H.S.; Naugher, K. Comparison of several types of enrichment for captive felids. Zoo Biol. 2007, 26, 371–381. [Google Scholar]
- Van Metter, J.E.; Harriger, M.D.; Bolen, R.H. Environmental enrichment utilizing stimulus objects for African lions (Panthera leo leo) and Sumatran tigers (Panthera tigris sumatrae). Bios 2008, 79, 7–16. [Google Scholar] [CrossRef]
- Gardiánová, I.; Bolechová, P. Tigers Stereotypic Pacing and Enrichment. Annu. Res. Rev. Biol. 2014, 4, 1544–1550. [Google Scholar] [CrossRef]
- Markowitz, H.; LaForse, S. Artificial prey as behavioral enrichment devices for felines. Appl. Anim. Behav. Sci. 1987, 18, 31–43. [Google Scholar]
- Phillips, C.J.; Tribe, A.; Lisle, A.; Galloway, T.K.; Hansen, K. Keepers’ rating of emotions in captive big cats, and their use in determining responses to different types of enrichment. J. Vet. Behav. 2017, 20, 22–30. [Google Scholar] [CrossRef]
- Pitsko, L.E. Wild Tigers in Captivity: A Study of the Effects of the Captive Environment on Tiger Behavior. Ph.D. Thesis, Virginia Tech, Blacksburg, VA, USA, 2003. [Google Scholar]
- Law, G.; Kitchener, A.C. Twenty years of the tiger feeding pole: Review and recommendations. Int. Zoo Yearb. 2020, 54, 174–190. [Google Scholar] [CrossRef]
- Eloff, F.C. Food ecology of the Kalahari lion Panthera leo vernayi. Koedoe 1984, 27, 249–258. [Google Scholar] [CrossRef]
- Gygax, L. Wanting, liking and welfare: The role of affective states in proximate control of behaviour in vertebrates. Ethology 2017, 123, 689–704. [Google Scholar] [CrossRef]
- Tarou, L.R.; Bashaw, M.J. Maximizing the effectiveness of environmental enrichment: Suggestions from the experimental analysis of behavior. Appl. Anim. Behav. Sci. 2007, 102, 189–204. [Google Scholar] [CrossRef]
- Damasceno, J.; Genaro, G.; Quirke, T.; McCarthy, S.; McKeown, S.; O’Riordan, R. The effects of intrinsic enrichment on captive felids. Zoo Biol. 2017, 36, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Quirke, T.; O’Riordan, R.M. The effect of a randomised enrichment treatment schedule on the behaviour of cheetahs (Acinonyx jubatus). Appl. Anim. Behav. Sci. 2011, 135, 103–109. [Google Scholar] [CrossRef]
- Szokalski, M.S.; Litchfield, C.A.; Foster, W.K. Enrichment for captive tigers (Panthera tigris): Current knowledge and future directions. Appl. Anim. Behav. Sci. 2012, 139, 1–9. [Google Scholar] [CrossRef]
- Regaiolli, B.; Rizzo, A.; Ottolini, G.; Miletto Petrazzini, M.E.; Spiezio, C.; Agrillo, C. Motion illusions as environmental enrichment for zoo animals: A preliminary investigation on lions (Panthera leo). Front. Psychol. 2019, 10, 2220. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, E.J.; Martin, A.L. Animal training, environmental enrichment, and animal welfare: A history of behavior analysis in zoos. J. Zool. Bot. Gard. 2021, 2, 531–543. [Google Scholar] [CrossRef]
- Fernandez, E.J.; Timberlake, W. Selecting and testing environmental enrichment in lemurs. Front. Psychol. 2019, 10, 482182. [Google Scholar] [CrossRef]
- Alligood, C.A.; Dorey, N.R.; Mehrkam, L.R.; Leighty, K.A. Applying behavior-analytic methodology to the science and practice of environmental enrichment in zoos and aquariums. Zoo Biol. 2017, 36, 175–185. [Google Scholar] [CrossRef]
- Podturkin, A.A.; Papaeva, N. Development of an environmental enrichment programme: Case study of white Bengal tiger (Panthera tigris bengalensis) and jaguar (Panthera onca) at Moscow Zoo. J. Zoo Aquar. Res. 2020, 8, 139–145. [Google Scholar]
- Barber, J. Effects of Food Distribution and External Factors on the Activity Budgets of Captive Sun Bears (Helarctos malayanus). Master’s Thesis, Sonoma State University, Rohnert Park, CA, USA, 2018. [Google Scholar]
- Brereton, J.E. Challenges and directions in zoo and aquarium food presentation research: A review. J. Zool. Bot. Gard. 2020, 1, 2. [Google Scholar] [CrossRef]
- Kleinlugtenbelt, C.L.; Burkevica, A.; Clauss, M. Large carnivore feeding in European zoos. Zool. Gart. 2023, 91, 9–39. [Google Scholar]
- Farmer, H.; Dayrell, E.; Pullen, K. Encouraging enclosure use in red river hogs using scatter feeding. Shape Enrich. 2006, 15, 11–15. [Google Scholar]
- Haas, S.K.; Hayssen, V.; Krausman, P.R. Panthera leo. Mamm. Species 2005, 762, 1–11. [Google Scholar] [CrossRef]
- De Cuyper, A.; Clauss, M.; Carbone, C.; Codron, D.; Cools, A.; Hesta, M.; Janssens, G.P. Predator size and prey size–gut capacity ratios determine kill frequency and carcass production in terrestrial carnivorous mammals. Oikos 2019, 128, 13–22. [Google Scholar] [CrossRef]
- Clubb, R.; Mason, G. Captivity effects on wide-ranging carnivores. Nature 2003, 425, 473–474. [Google Scholar] [CrossRef] [PubMed]
- Finch, K.; Williams, L.; Holmes, L. Using longitudinal data to evaluate the behavioural impact of a switch to carcass feeding on an Asiatic lion (Panthera leo persica). J. Zoo Aquar. Res. 2020, 8, 283–287. [Google Scholar]
- Mellor, D.J. Updating animal welfare thinking: Moving beyond the “Five Freedoms” towards “a Life Worth Living”. Animals 2016, 6, 21. [Google Scholar] [CrossRef]
- Little, K.A.; Sommer, V. Change of enclosure in langur monkeys: Implications for the evaluation of environmental enrichment. Zoo Biol. 2002, 21, 549–559. [Google Scholar] [CrossRef]
- Lukas, K.E.; Hoff, M.P.; Maple, T.L. Gorilla behavior in response to systematic alternation between zoo enclosures. Appl. Anim. Behav. Sci. 2003, 81, 367–386. [Google Scholar] [CrossRef]
- AZA Lion SSP. Lion Care Manual; Association of Zoos and Aquariums: Silver Spring, MD, USA, 2012. [Google Scholar]
- Höttges, N.; Hjelm, M.; Hård, T.; Laska, M. How does feeding regime affect behaviour and activity in captive African lions (Panthera leo)? J. Zoo Aquar. Res. 2019, 7, 117–125. [Google Scholar]
- Packer, C.; Pusey, A.E.; Eberly, L.E. Egalitarianism in female African lions. Science 2001, 293, 690–693. [Google Scholar] [CrossRef] [PubMed]
- Altman, J.D.; Gross, K.L.; Lowry, S.R. Nutritional and behavioral effects of gorge and fast feeding in captive lions. J. Appl. Anim. Welf. Sci. 2005, 8, 47–57. [Google Scholar] [CrossRef]
- Hamilton, J.; Fuller, G.; Allard, S. Evaluation of the impact of behavioral opportunities on four zoo-housed Aardvarks (Orycteropus afer). Animals 2020, 10, 1433. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Pride | Lion | Sex/Birth Year | Relation |
|---|---|---|---|
| Pride 1 (Priscilla’s pride) | Priscilla | Female/1998 | Pride leader |
| Rah | Male/2004 | Son of Priscilla Castrated | |
| Leah | Female/2005 | Daughter of Priscilla | |
| Pride 2 (Kiara’s pride) | Kiara | Female/1998 | Former pride leader |
| Mishka | Female/2004 | Current pride leader, Daughter of Kiara | |
| Sakura | Male/2005 | Son of Kiara, castrated | |
| Kahn | Male/2005 | Son of Kiara, castrated | |
| Toa | Male/2009 | Son of Kiara, vasectomized during study | |
| Tawhiri | Male/2009 | Son of Kiara, castrated | |
| Kairangi | Male/2009 | Son of Kiara, castrated | |
| Tua | Female/2009 | Daughter of Mishka | |
| Tama | Female/2009 | Daughter of Mishka |
| Class | Behaviour | Definition |
|---|---|---|
| Locomote | Walking | Moving slowly |
| Running | Moving fast | |
| Jumping | At least two paws leave the ground | |
| Rolling/stretching | Extended reach while lying down | |
| Rest | Sitting | Backside on ground, while upright |
| Lying Down | At least one side on ground | |
| Standing | Upright with no movement | |
| Bathing | Non-drinking contact with water | |
| Stereotypy | Pacing | Repetitive movement; at least one back-and-forth loop |
| Explore | Sniffing | Head lifted while inhaling |
| Digging | Using paws to move dirt | |
| Interacting with object | Any contact with moveable objects (other than study enrichment) | |
| Interacting with enrichment | Contact with experimental enrichment item, including eating of food-based enrichment items | |
| Eat | Eating | Mouth contact with edible food items, excluding consumption of enrichment items |
| Drinking | Mouth contact with water (non-bathing) | |
| Social | Interacting with other lion | Any contact with another lion |
| Vocalising | Sound made by a lion, typically directed at another lion | |
| Stalking | Crouched hunting position directed at another lion | |
| Grooming | Licking/pawing body of self or conspecific | |
| Territorial | Scratch marking | Claw contact with non-movable structure |
| Scent marking | Urine directed at a non-movable structure | |
| Rubbing | Face/body contact with non-movable structure | |
| Other | Urinate/defecate | Non-directed excretion |
| Out of sight | Not visible | |
| Other | Behaviour that does not meet above criteria |
| Days | Condition | Type | Description |
|---|---|---|---|
| 1 to 4 | Baseline | N/A | No additional enrichment used beyond daily lion truck feed |
| 5 | Food Enrichment | Ice Blocks | 4 L of fresh blood mixed with water for 4 kg frozen blocks. Pride 1 received 3 blocks and pride 2 received 5 blocks |
| 6 | Non-Food Enrichment | Mirrors | 2 mirrors per pride were tied at head height outside of enclosures |
| 7 to 10 | Not Tested | N/A | Varied enrichment, not directly tested |
| 11 | Food Enrichment | Gelatine | 3 L fresh blood mixed with 125 g of gelatine (frozen moulds). Pride 1 received 3 moulds and pride 2 received 6 moulds |
| 12 | Non-Food Enrichment | Lavender | 20 fresh lavender branches per exhibit scattered in each enclosure |
| 13 | Non-Food Enrichment | Clean Sheet | Non-scented, clean sheet tied to a tree/trunk, one in each exhibit |
| 14 | Food Enrichment | Eggs | 10 hard-boiled eggs per exhibit scattered in each enclosure |
| 15 to 16 | Baseline | N/A | No additional enrichment used. |
| Behavior Class | Baseline | Food | Non-Food |
|---|---|---|---|
| Locomote | 17.42 (±0.88) a | 12.19 (±1.34) b | 11.50 (±0.79) b |
| Rest | 56.86 (±2.71) a | 28.93 (±2.51) b | 36.44 (±3.92) b |
| Stereotypy | 3.178 (±1.29) | 0 | 0 |
| Explore | 2.12 (±0.74) a | 40.51 (±3.57) b | 32.31 (±3.43) b |
| Eat | 0.12 (±0.06) | 0.03 (±0.03) | 0.17 (±0.09) |
| Social | 19.15 (±2.52) | 17.85 (±2.46) | 18.60 (±2.01) |
| Territorial | 0.61 (±0.07) | 0.47 (±0.10) a | 0.98 (±0.16) b |
| Other | 0.54 (±0.29) | 0.03 (±0.03) | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cremers, P.; Norman, M.; Brando, S.; Fernandez, E.J. A Comparison of Food and Non-Food Enrichment with Zoo-Housed African Lions (Panthera leo). J. Zool. Bot. Gard. 2025, 6, 25. https://doi.org/10.3390/jzbg6020025
Cremers P, Norman M, Brando S, Fernandez EJ. A Comparison of Food and Non-Food Enrichment with Zoo-Housed African Lions (Panthera leo). Journal of Zoological and Botanical Gardens. 2025; 6(2):25. https://doi.org/10.3390/jzbg6020025
Chicago/Turabian StyleCremers, Peggy, Max Norman, Sabrina Brando, and Eduardo J. Fernandez. 2025. "A Comparison of Food and Non-Food Enrichment with Zoo-Housed African Lions (Panthera leo)" Journal of Zoological and Botanical Gardens 6, no. 2: 25. https://doi.org/10.3390/jzbg6020025
APA StyleCremers, P., Norman, M., Brando, S., & Fernandez, E. J. (2025). A Comparison of Food and Non-Food Enrichment with Zoo-Housed African Lions (Panthera leo). Journal of Zoological and Botanical Gardens, 6(2), 25. https://doi.org/10.3390/jzbg6020025