
Behavioral and Spatial Analysis of a Symphalangus syndactylus Pair in a Controlled Environment

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Pair of Siamangs
2.2. Exhibit
- The outdoor part of the exhibit has three sides and the roof is closed by a wire mesh, while the other side has three glass windows. This section has a trapezoidal shape with dimensions of 10 m × 12 m × 10 m × 14.5 m and a height ranging from 4 m to 6 m;
- The wire meshed tunnel, connecting indoor and outdoor spaces, has a maximum height of 5 m and a total length of 20 m;
- The indoor part of the exhibit has a rectangular shape with approximate dimensions of 3 m × 2 m and a height of 2.5 m.
2.3. Ethogram
- Scratching referred to the use of fingers or nails to scratch various parts of the body, often through quick and repetitive movements.
- For allogrooming (T-N) and (N-T), the notation (T-N) indicated that Tomas was the grooming individual, providing allogrooming to Nina, whereas (N-T) signified the reverse, with Nina grooming Tomas.
- Acrobatic swing: In solitary play, the siamang moved independently on ropes, sometimes hanging upside down or using ropes like a swing, often rapidly and in combination with other play behaviors. In social play, this occurred within a chase session (Chase)
- Chase: Either individuals engaged in playful pursuit, chasing one another or, less frequently, external objects.
- Grapple/Bite: Individuals grasped and rolled together while playfully biting, usually on the ground.
- Grunt/Growl: Playful wrestling was combined with grunting and growling vocalizations, typically occurring on the ground.
- Manipulation play: The individual manipulated an object, often a detached palm frond, while moving on ropes, not for locomotion but as part of play.
- Roll: The individual curled up and rolled by pushing off with its forelimbs.
- Somersault: The individual performed a full-body rotation, typically while in the air, completing a flip.
- Slap: A quick, forceful strike was made with an open hand to another individual.
- Open mouth: The individual opened its mouth widely, approaching the face of another individual while displaying its teeth.
- Singing: For example, SI-AM indicated that the micro-categories reflected the behaviors observed during vocalizations.
- Manipulation: The siamang interacted with an object.
- Catching insects: The individual captured insects using its limbs, often consuming them afterward.
- Look: The individual remained still while observing the surroundings, occasionally turning its head in different directions.
- Hanging rest: The siamang hung motionless for an extended period, either from the mesh or from pole.
- Lie down: The siamang was in a fully reclined position.
- Sit: The individual remained seated without movement, appearing to rest but with open eyes.
- Sleep: The siamang was completely at rest with their eyes closed.
- Feed keeper: This referred to food being provided directly by keepers during feeding sessions. It included specific items offered to the animals as part of their diet in a controlled environment.
- Feed weed: This referred to all food sources that were not part of the provided diet by keepers, but rather those that the animals obtained independently, such as grass from the outdoor area, palm fruits, leaves, and other naturally available plants.
- Touch indicated a light and slow contact made with a limb on another individual.
- Hug indicated that the animals embraced, with one or more limbs fully wrapping around another individual.
- Hang move indicated that the siamang moved slowly while hanging from mesh or pole.
- Walk indicated a slow, bipedal or quadrupedal movement, occurring either on the ground or while balancing on poles.
- Swing indicated a rhythmic, repetitive motion while suspended from ropes or palm fronds.
- Acrobatic move indicated a rapid and agile movement, involving leaps between poles, ropes, palm fronds, or the mesh.
- Defecate indicated all physiological elimination behaviors.
- Sexual behavior included mounting and copulation.
2.4. Data Collection
2.5. Data Analysis
- Singing frequency;
- Spatial analysis of the relative spatial distribution of singing behavior around the exhibit space;
- Time of day in one-hour time slots;
- Duration, subdivided into six durational categories.
3. Results
3.1. Spatial Analysis and Time Budget
3.2. Singing
4. Discussion
4.1. Time Budget and Spatial Analysis
4.2. Analyses of Singing
4.3. Comparisons Between First and Second Observation Periods
4.4. Singing Comparisons Between the First and Second Observation Periods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Nebasifu, A.A. Ex-Situ Conservation: Primate Protection in the Limbe Wildlife Centre. J. Zool. Stud. 2015, 2, 12–21. [Google Scholar]
- Miranda, R.; Escribano, N.; Casas, M.; Pino-del-Carpio, A.; Villarroya, A. The Role of Zoos and Aquariums in a Changing World. Annu. Rev. Anim. Biosci. 2023, 11, 287–306. [Google Scholar] [CrossRef] [PubMed]
- Wolfensohn, S.; Shotton, J.; Bowley, H.; Davies, S.; Thompson, S.; Justice, W.S.M. Assessment of Welfare in Zoo Animals: Towards Optimum Quality of Life. Animals 2018, 8, 110. [Google Scholar] [CrossRef] [PubMed]
- Sueur, C.; Pelé, M. Importance of Living Environment for the Welfare of Captive Animals: Behaviours and Enrichment. In Proceedings of the Animal Welfare: From Science to Law, Palazzo dell’UNESCO, Parigi, France, 10–11 December 2015. [Google Scholar]
- Gil-Dolz, J.; Ayuso, P.R.; Riba, D.; Crailsheim, D. Neighbors, Pros and Cons: Impact of Intergroup Interactions on the Welfare of Captive Chimpanzee Groups (Pan troglodytes). Ecologies 2024, 5, 279–295. [Google Scholar] [CrossRef]
- Coe, J.C. Naturalizing Habitats for Captive Primates. Zoo Biol. 1989, 8, 117–125. [Google Scholar] [CrossRef]
- Williams, E.; Cabana, F.; Nekaris, K.A.I. Improving Diet and Activity of Insectivorous Primates in Captivity: Naturalizing the Diet of Northern Ceylon Gray Slender Loris, Loris Lydekkerianus Nordicus. Zoo Biol. 2015, 34, 473–482. [Google Scholar] [CrossRef]
- McPhee, M.E.; Carlstead, K. The Importance of Maintaining Natural Behaviors in Captive Mammals. In Wild Mammals in Captivity: Principles and Techniques for Zoo Management, 2nd ed.; Kleiman, D.G., Thompson, K.V., Charlotte, K.B., Eds.; University of Chicago Press: Chicago, IL, USA, 2010; pp. 303–313. ISBN 978-0-226-44010-1. [Google Scholar]
- Yanuar, A. The Population Distribution and Abundance of Siamangs (Symphalangus syndactylus) and Agile Gibbons (Hylobates agilis) in West Central Sumatra, Indonesia. In The Gibbons; Springer: New York, NY, USA, 2009; pp. 453–465. ISBN 978-0-387-88603-9. [Google Scholar]
- Malone, N.; Fuentes, A.; White, F.J. Variation in the Social Systems of Extant Hominoids: Comparative Insight into the Social Behavior of Early Hominins. Int. J. Primatol. 2012, 33, 1251–1277. [Google Scholar] [CrossRef]
- Hankinson, E. Spatial Distribution and Density of the Lar Gibbon Hylobates Lar and Siamang Symphalangus Syndactylus in Relation to Canopy Structure and Disturbance in a Lowland Dipterocarp Forest, Sumatra. Master’s Thesis, Bournemouth University, Poole, UK, 2016. [Google Scholar]
- O’Brien, T.G.; Kinnaird, M.F.; Nurcahyo, A.; Prasetyaningrum, M.; Iqbal, M. Fire, Demography and the Persistence of Siamang (Symphalangus syndactylus: Hylobatidae) in a Sumatran Rainforest. Anim. Conserv. 2003, 6, 115–121. [Google Scholar] [CrossRef]
- Nijman, V.; Geissmann, T.; Traeholt, C.; Roos, C.; Nowak, M.G. Symphalangus Syndactylus. In The IUCN Red List of Threatened Species 2020; IUCN: Gland, Switzerland, 2015; E.T39779A17967873. [Google Scholar]
- Langbein, J.; Nawroth, C. Editorial: Captive Animal Behavior: Individual Differences in Learning and Cognition, and Implications on Animal Welfare. Front. Vet. Sci. 2022, 9, 1102122. [Google Scholar] [CrossRef]
- Chivers, D.J. Communication Within and Between Family Groups of Siamang (Symphalangus syndactylus). Behaviour 1976, 57, 116–135. [Google Scholar] [CrossRef]
- Lappan, S. Male Care of Infants in a Siamang (Symphalangus syndactylus) Population Including Socially Monogamous and Polyandrous Groups. Behav. Ecol. Sociobiol. 2008, 62, 1307–1317. [Google Scholar] [CrossRef]
- Lappan, S.; Morino, L. Mating in the Presence of a Competitor: Audience Effects May Promote Male Social Tolerance in Polyandrous Siamang (Symphalangus syndactylus) Groups. Behaviour 2014, 151, 1067–1089. [Google Scholar] [CrossRef]
- Morino, L. Social Correlates of Androgen Levels in a Facultatively Monogamous Ape (Symphalangus syndactylus): A Test of the Challenge Hypothesis. Behav. Ecol. Sociobiol. 2015, 69, 243–251. [Google Scholar] [CrossRef]
- Morino, L.; Pasquaretta, C.; Sueur, C.; MacIntosh, A.J.J. Communication Network Reflects Social Instability in a Wild Siamang (Symphalangus syndactylus) Population. Int. J. Primatol. 2021, 42, 618–639. [Google Scholar] [CrossRef]
- Dewi, D.P.; Iskandar, E.; Perwitasari-Farajallah, D. The Feeding Behavior and Food Preferences of Siamang (Symphalangus syndactylus Raffles, 1821) at Taman Safari Indonesia Bogor. IOP Conf. Ser. Earth Environ. Sci. 2023, 1271, 012048. [Google Scholar] [CrossRef]
- Vaglio, S.; Kaburu, S.S.K.; Pearce, R.; Bryant, L.; McAuley, A.; Lott, A.; Sheppard, D.J.; Smith, S.; Tompkins, B.E.; Elwell, E.J.; et al. Effects of Scent Enrichment on Behavioral and Physiological Indicators of Stress in Zoo Primates. Am. J. Primatol. 2021, 83, e23247. [Google Scholar] [CrossRef]
- Hosey, G.; Ward, S.; Melfi, V. The Effect of Visitors on the Behaviour of Zoo-Housed Primates: A Test of Four Hypotheses. Appl. Anim. Behav. Sci. 2023, 263, 105938. [Google Scholar] [CrossRef]
- Sacchet, E.; Milesi, A.; Preatoni, D.; Castiglioni, R. Symphalangus syndactylus (Raffles, 1821) in Ambiente Controllato. Comportamento Vocale e Sociale, uso Dello Spazio e Prossemica. Bachelor’s Thesis, Università Studiorum Insubriae, Varese, Italy, 2020. [Google Scholar]
- Mittermeier, R.A.; Rylands, A.B.; Wilson, D.E. Handbook of the Mammals of the World; Lynx: Barcelona, Spain, 2013; Volume 3. [Google Scholar]
- Poletti, S.; Castiglioni, R.; Gamba, M. Comportamento Sociale e Vocale di una Coppia di Siamanghi (Symphalangus syndactylus) in Ambiente Controllato. Master’s Thesis, Università degli Studi di Torino, Turin, Italy, 2020. [Google Scholar]
- Harrison, N.J.; Hill, R.A.; Alexander, C.; Marsh, C.D.; Nowak, M.G.; Abdullah, A.; Slater, H.D.; Korstjens, A.H. Sleeping Trees and Sleep-Related Behaviours of the Siamang (Symphalangus syndactylus) in a Tropical Lowland Rainforest, Sumatra, Indonesia. Primates 2021, 62, 63–75. [Google Scholar] [CrossRef]
- Orgeldinger, M. Protective and Territorial Behavior in Captive Siamangs (Hylobates syndactylus). Zoo Biol. 1997, 16, 309–325. [Google Scholar] [CrossRef]
- Castiglioni, R.; Sacchet, E.; Milesi, A.; Preatoni, D. Singing Behavior of Siamang Group Living in Captivity. In Proceedings of the XII Convegno Nazionale della Ricerca nei Parchi, Bussolengo, Spain, 7–9 October 2022. [Google Scholar]
- Pearson, E.L.; Davis, J.M.; Litchfield, C.A. A Case Study of Orangutan and Siamang Behavior Within a Mixed-Species Zoo Exhibit. J. Appl. Anim. Welf. Sci. 2010, 13, 330–346. [Google Scholar] [CrossRef]
- Liebal, K.; Pika, S.; Tomasello, M. Social Communication in Siamangs (Symphalangus syndactylus): Use of Gestures and Facial Expressions. Primates 2004, 45, 41–57. [Google Scholar] [CrossRef] [PubMed]
- Zulamri Morning Call of Siamang (Symphalangus syndactyllus) in Subayang River, Riau—Indonesia. Int. J. Ecophysiol. 2019, 1, 125–130. [CrossRef]
- Chamove, A.S.; Hosey, G.R.; Schaetzel, P. Visitors Excite Primates in Zoos. Zoo Biol. 1988, 7, 359–369. [Google Scholar] [CrossRef]
- Sherwen, S.L.; Hemsworth, P.H. The Visitor Effect on Zoo Animals: Implications and Opportunities for Zoo Animal Welfare. Animals 2019, 9, 366. [Google Scholar] [CrossRef]
- Maple, T.L.; Perdue, B.M. Wellness as Welfare. In Zoo Animal Welfare; Maple, T., Perdue, B.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 49–67. ISBN 978-3-642-35955-2. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| Sex | Date of Birth | Date of Acquisition | Local ID | Origin |
|---|---|---|---|---|
| ♀ | 12 March 1997 | 31 March 1999 | Nina | Planete Sauvage Parc Zoologique, Port-Saint-Père, France |
| ♂ | 2 June 2000 | 13 April 2005 | Tomas | Zoo de Besancon, France |
| Macro-Category | Micro-Category |
|---|---|
| Autogrooming (AUTOGR) | - Scratch (SC) |
| Allogrooming (ALLOGR) | Nina–Tomas (N-T) Tomas–Nina (T-N) |
| Play (PL) [Solitary (PLS); With (PLW)] | Acrobatic swing (AS) Chase (CH)/Grapple/Bite Grunt (GB)/Growl (GG) Manipulation play (MP) Roll (RO) Somersault (SM) |
| Conflict (CO) | Slap (SP) Open mouth (OM) |
| Sing (SI) | Sing followed by another behavioral model (e.g., SI-AM) |
| Manipulation (MA) | - Catch insect (CI) |
| Rest (RE) | Look (RE) Hanging resting (HN RE) Lie down (LD) Sit (ST) Sleep (SL) |
| Drink (DK) | - |
| Feed (FE) | Feed keeper (FE K) Feed weed (FE W) |
| Body contact (BC) | Touch (TC) Hug (HG) |
| Move (MO) | Hanging moving (HN MO) Walk (WA) Swing (SW) Acrobatic move (AM) |
| Other (OT) | Defecate (DE) Sexual behavior (SX) |
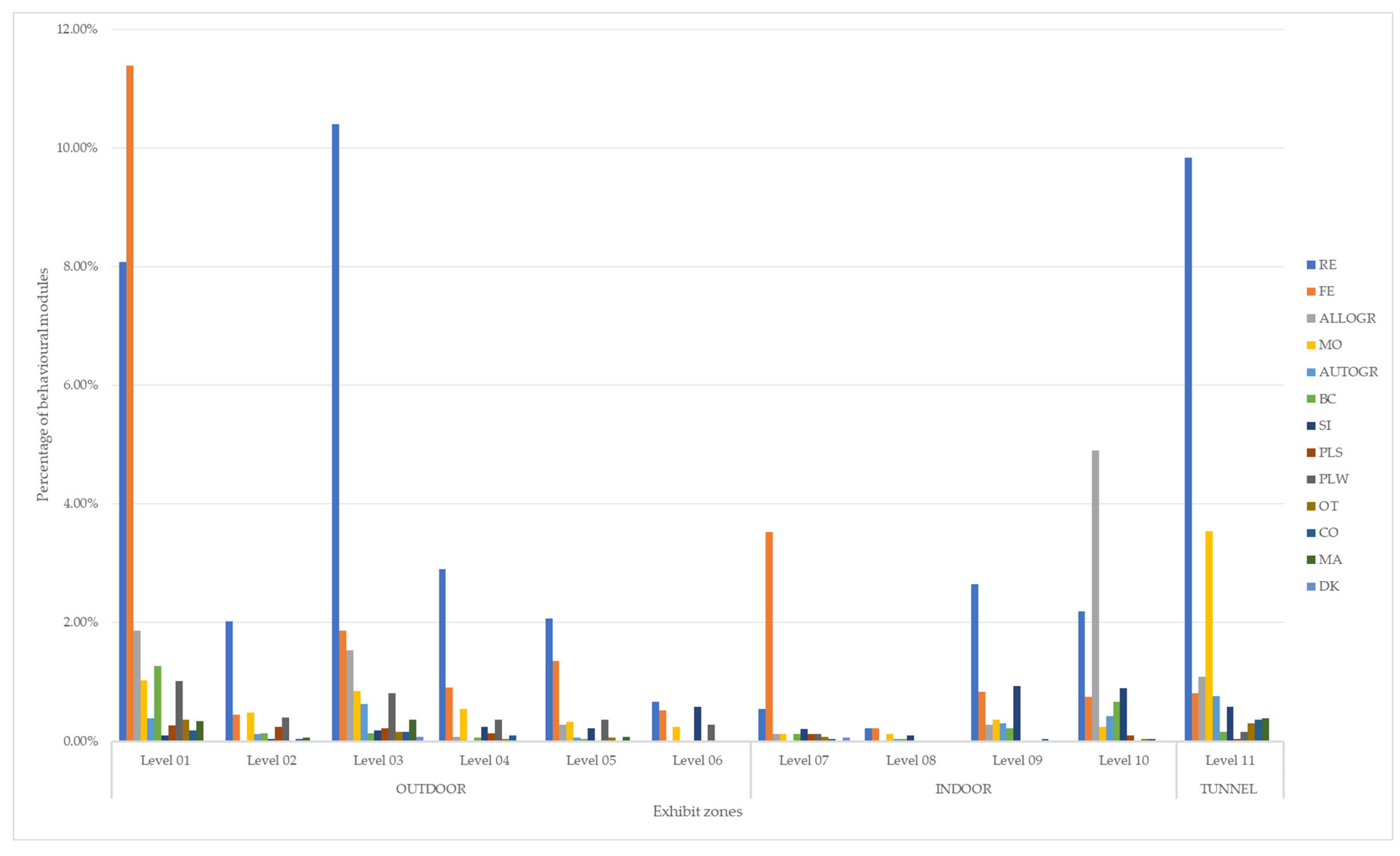
| LEVELS | RE | FE | ALLOGR | MO | AUTOGR | BC | SI | PLS | PLW | OT | CO | MA | DK | TOTAL |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| OUTDOOR | 26.12% | 16.48% | 3.76% | 3.47% | 1.21% | 1.66% | 1.37% | 0.87% | 3.23% | 0.65% | 0.48% | 0.87% | 0.08% | 60.26% |
| Level 01 | 8.08% | 11.39% | 1.86% | 1.03% | 0.38% | 1.27% | 0.10% | 0.26% | 1.01% | 0.36% | 0.18% | 0.34% | 0.00% | 26.28% |
| Level 02 | 2.02% | 0.44% | 0.00% | 0.48% | 0.12% | 0.14% | 0.04% | 0.24% | 0.40% | 0.00% | 0.04% | 0.06% | 0.00% | 4.00% |
| Level 03 | 10.40% | 1.86% | 1.54% | 0.85% | 0.63% | 0.14% | 0.18% | 0.22% | 0.81% | 0.16% | 0.16% | 0.36% | 0.08% | 17.39% |
| Level 04 | 2.89% | 0.91% | 0.08% | 0.55% | 0.02% | 0.06% | 0.24% | 0.14% | 0.36% | 0.04% | 0.10% | 0.02% | 0.00% | 5.41% |
| Level 05 | 2.06% | 1.35% | 0.28% | 0.32% | 0.06% | 0.04% | 0.22% | 0.00% | 0.36% | 0.06% | 0.00% | 0.08% | 0.00% | 4.85% |
| Level 06 | 0.67% | 0.53% | 0.00% | 0.24% | 0.00% | 0.00% | 0.59% | 0.00% | 0.28% | 0.02% | 0.00% | 0.00% | 0.00% | 2.32% |
| INDOOR | 5.60% | 5.31% | 5.29% | 0.85% | 0.79% | 1.05% | 2.12% | 0.24% | 0.12% | 0.12% | 0.12% | 0.02% | 0.06% | 21.70% |
| Level 07 | 0.55% | 3.52% | 0.12% | 0.12% | 0.02% | 0.12% | 0.20% | 0.12% | 0.12% | 0.08% | 0.04% | 0.00% | 0.06% | 5.07% |
| Level 08 | 0.22% | 0.22% | 0.00% | 0.12% | 0.04% | 0.04% | 0.10% | 0.00% | 0.00% | 0.00% | 0.00% | 0.02% | 0.00% | 0.77% |
| Level 09 | 2.65% | 0.83% | 0.28% | 0.36% | 0.30% | 0.22% | 0.93% | 0.02% | 0.00% | 0.00% | 0.04% | 0.00% | 0.00% | 5.64% |
| Level 10 | 2.18% | 0.75% | 4.89% | 0.24% | 0.42% | 0.67% | 0.89% | 0.10% | 0.00% | 0.04% | 0.04% | 0.00% | 0.00% | 10.22% |
| TUNNEL | 9.84% | 0.81% | 1.09% | 3.54% | 0.77% | 0.16% | 0.59% | 0.04% | 0.16% | 0.30% | 0.36% | 0.38% | 0.00% | 18.04% |
| Level 11 | 9.84% | 0.81% | 1.09% | 3.54% | 0.77% | 0.16% | 0.59% | 0.04% | 0.16% | 0.30% | 0.36% | 0.38% | 0.00% | 18.04% |
| TOTAL | 41.56% | 22.61% | 10.14% | 7.86% | 2.77% | 2.87% | 4.08% | 1.15% | 3.52% | 1.07% | 0.97% | 1.27% | 0.14% | 100.00% |
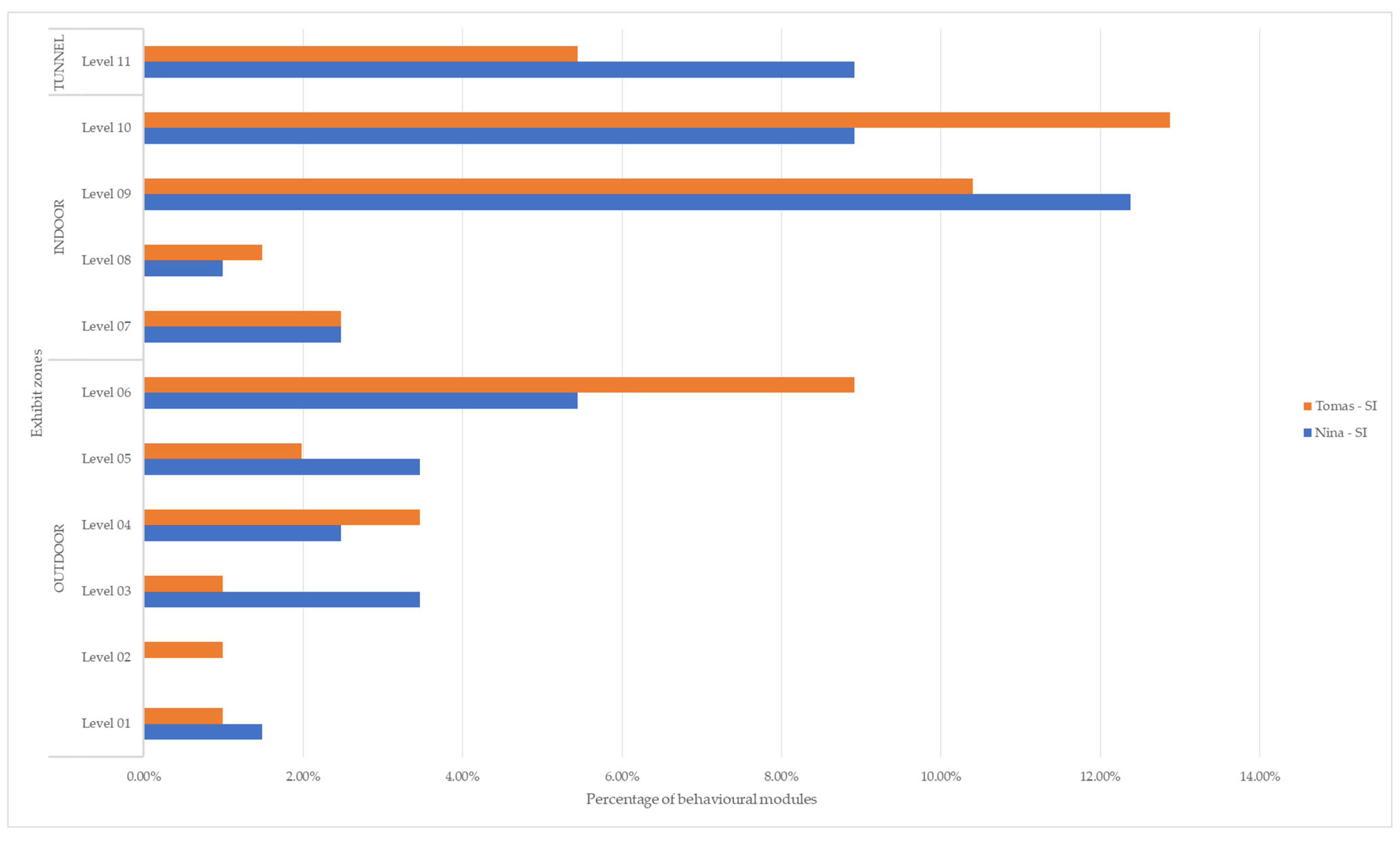
| LEVELS | Nina | Tomas | TOTAL |
|---|---|---|---|
| OUTDOOR | 16.34% | 17.33% | 33.66% |
| Level 01 | 1.49% | 0.99% | 2.48% |
| Level 02 | 0.00% | 0.99% | 0.99% |
| Level 03 | 3.47% | 0.99% | 4.46% |
| Level 04 | 2.48% | 3.47% | 5.94% |
| Level 05 | 3.47% | 1.98% | 5.45% |
| Level 06 | 5.45% | 8.91% | 14.36% |
| INDOOR | 24.75% | 27.23% | 51.98% |
| Level 07 | 2.48% | 2.48% | 4.95% |
| Level 08 | 0.99% | 1.49% | 2.48% |
| Level 09 | 12.38% | 10.40% | 22.77% |
| Level 10 | 8.91% | 12.87% | 21.78% |
| TUNNEL | 8.91% | 5.45% | 14.36% |
| Level 11 | 8.91% | 5.45% | 14.36% |
| TOTAL | 50.00% | 50.00% | 100.00% |
| Time Slots | TOTAL |
|---|---|
| 9:00–9:55 | 8.91% |
| 10:00–10:55 | 23.76% |
| 11:00–11:55 | 36.63% |
| 12:00–12:55 | 15.84% |
| 1:00–1:55 | 5.94% |
| 2:00–2:55 | 8.91% |
| 3:00–3:55 | 0.00% |
| 4:00–4:55 | 0.00% |
| 5:00–5:55 | 0.00% |
| Duration (minutes) | Total |
|---|---|
| 1–5 | 0.00% |
| 6–10 | 9.09% |
| 11–15 | 31.81% |
| 16–20 | 50.00% |
| 21–25 | 9.09% |
| 26–30 | 0.00% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carpino, C.; Castiglioni, R.; Sacchet, E.; Milesi, A.; Marano, L.; Leonetti, F.L.; Romano, V.; Giglio, G.; Sperone, E. Behavioral and Spatial Analysis of a Symphalangus syndactylus Pair in a Controlled Environment. J. Zool. Bot. Gard. 2025, 6, 23. https://doi.org/10.3390/jzbg6020023
Carpino C, Castiglioni R, Sacchet E, Milesi A, Marano L, Leonetti FL, Romano V, Giglio G, Sperone E. Behavioral and Spatial Analysis of a Symphalangus syndactylus Pair in a Controlled Environment. Journal of Zoological and Botanical Gardens. 2025; 6(2):23. https://doi.org/10.3390/jzbg6020023
Chicago/Turabian StyleCarpino, Chiara, Roberta Castiglioni, Elisa Sacchet, Arianna Milesi, Luca Marano, Francesco Luigi Leonetti, Viviana Romano, Gianni Giglio, and Emilio Sperone. 2025. "Behavioral and Spatial Analysis of a Symphalangus syndactylus Pair in a Controlled Environment" Journal of Zoological and Botanical Gardens 6, no. 2: 23. https://doi.org/10.3390/jzbg6020023
APA StyleCarpino, C., Castiglioni, R., Sacchet, E., Milesi, A., Marano, L., Leonetti, F. L., Romano, V., Giglio, G., & Sperone, E. (2025). Behavioral and Spatial Analysis of a Symphalangus syndactylus Pair in a Controlled Environment. Journal of Zoological and Botanical Gardens, 6(2), 23. https://doi.org/10.3390/jzbg6020023