
Does Observer Presence Modify the Behavior and Enclosure Use of Captive Edwards’ Pheasants?



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study site and Species
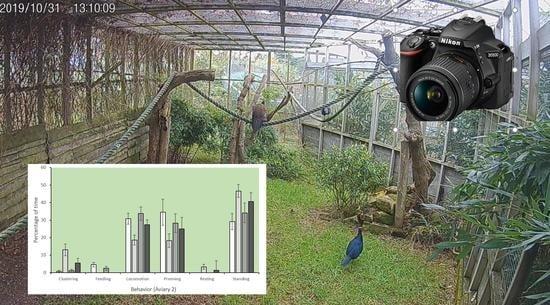
2.2. Behavior
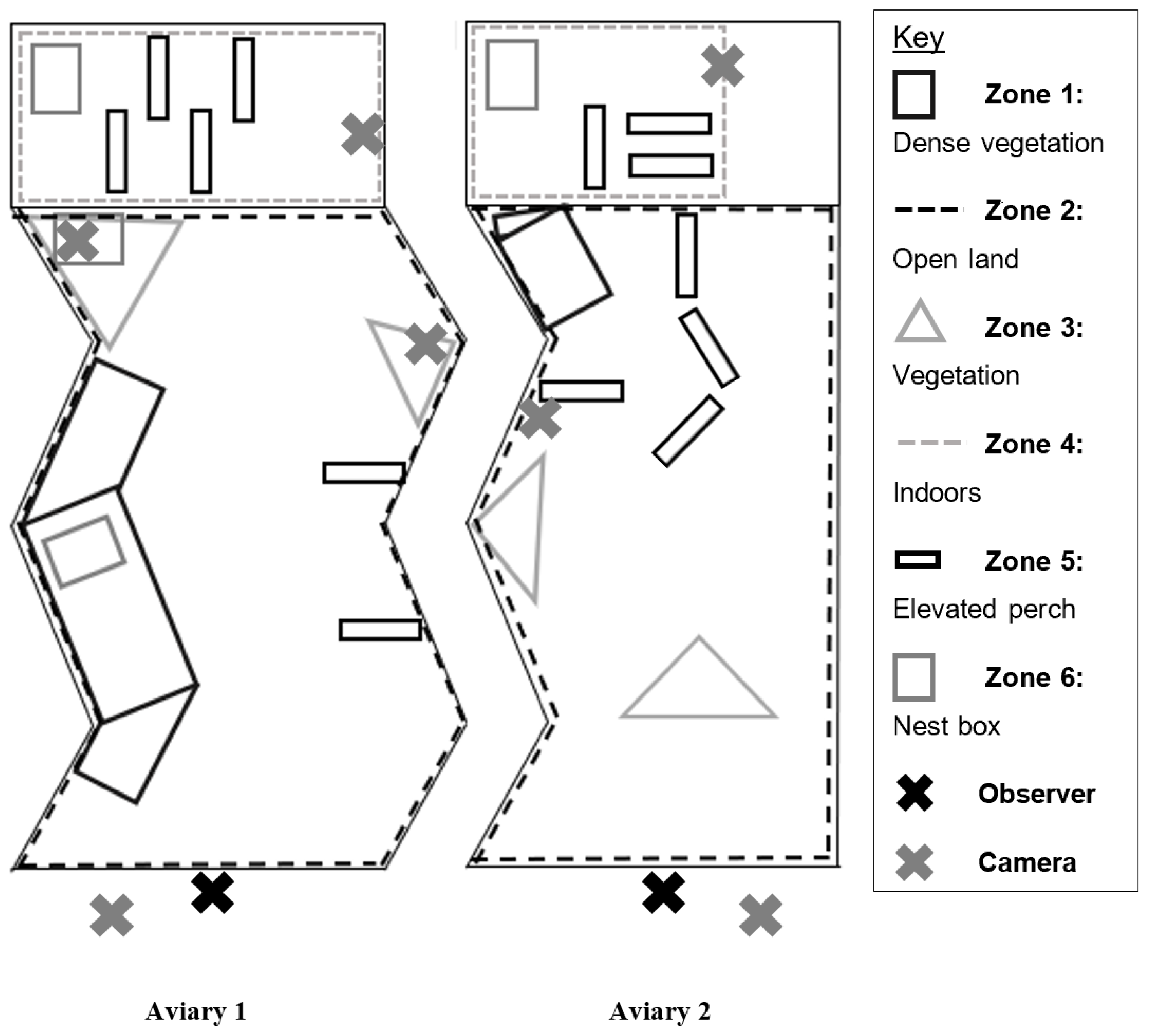
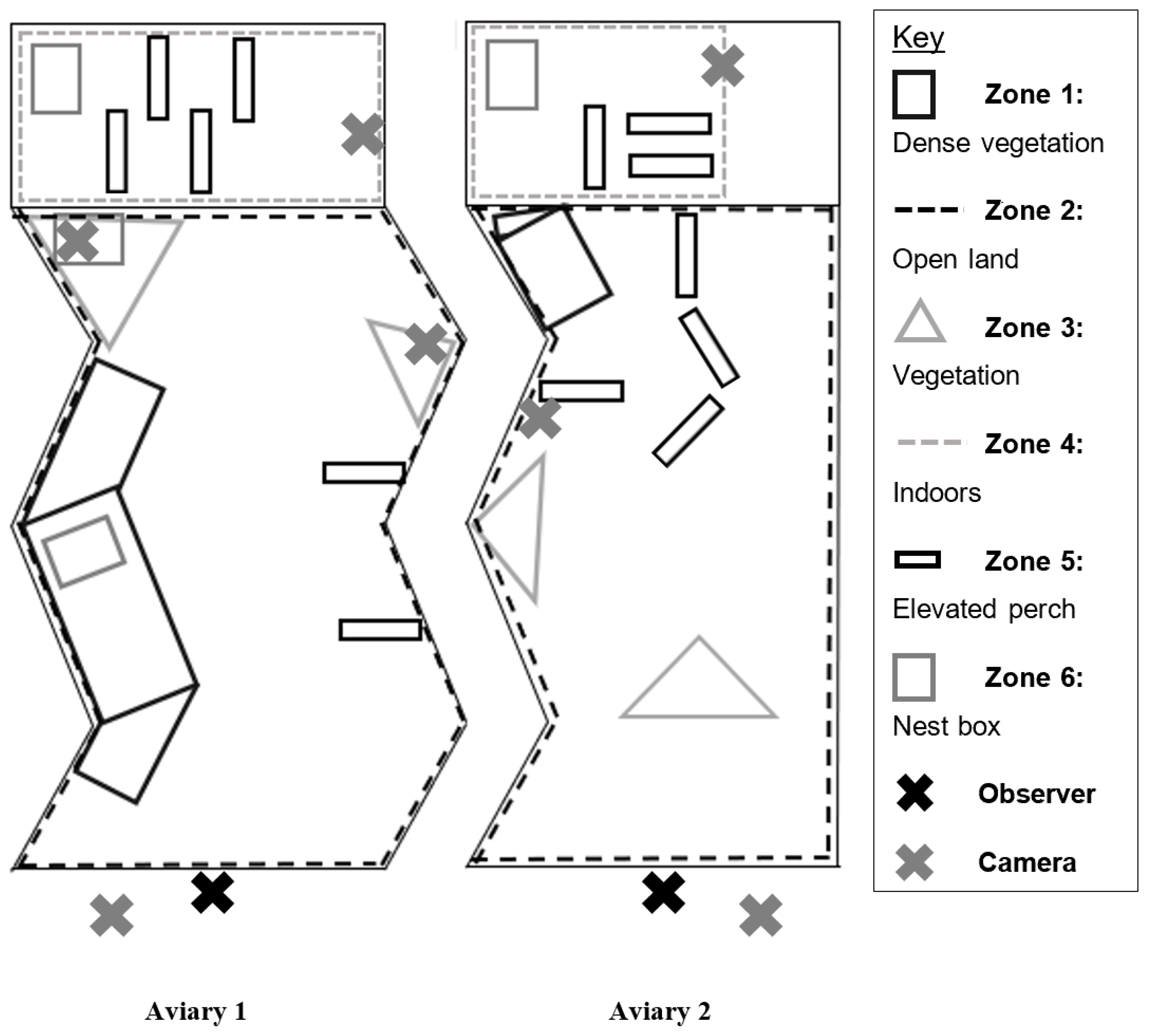
2.3. Enclosure Use
2.4. Data Analysis
3. Results
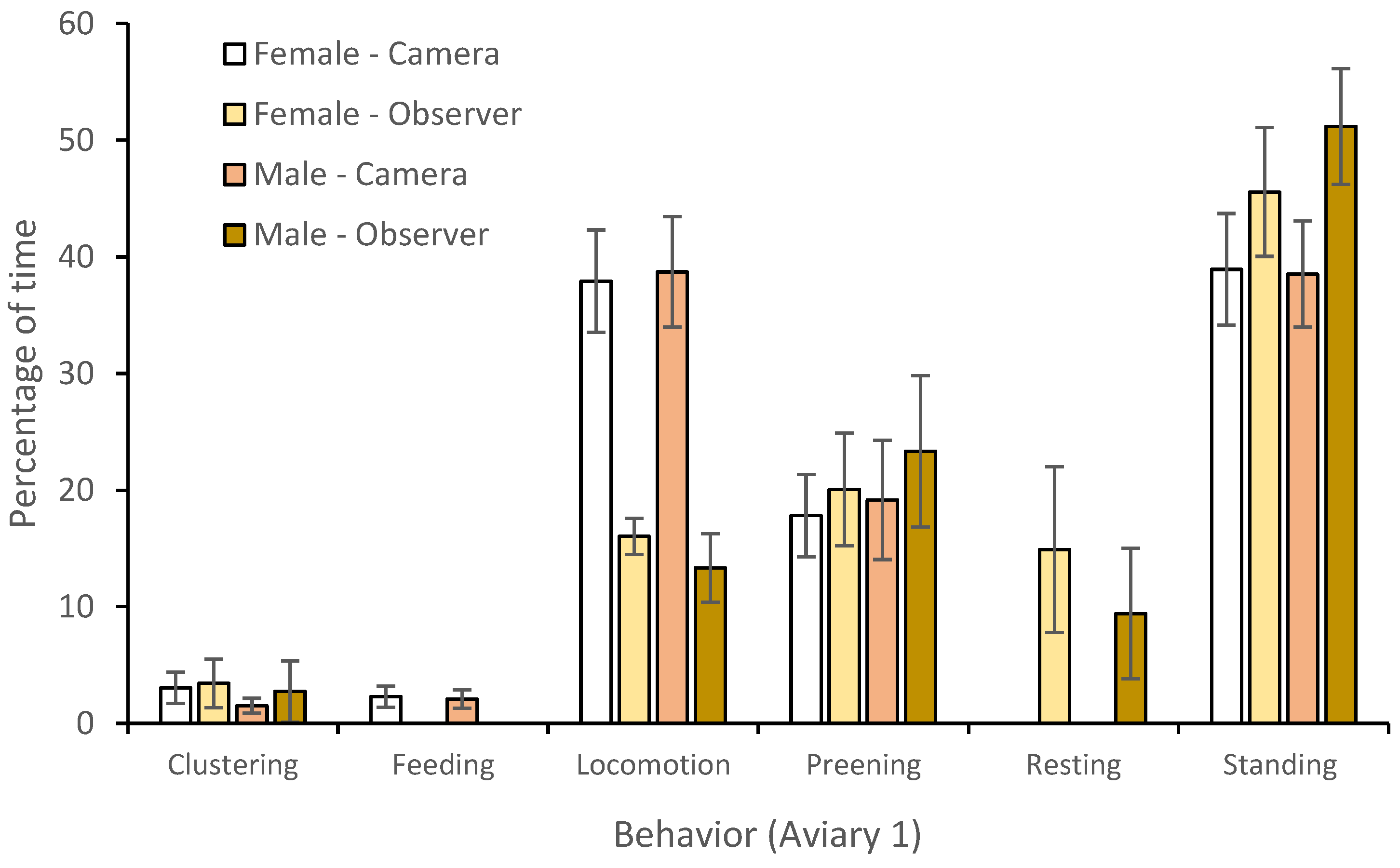
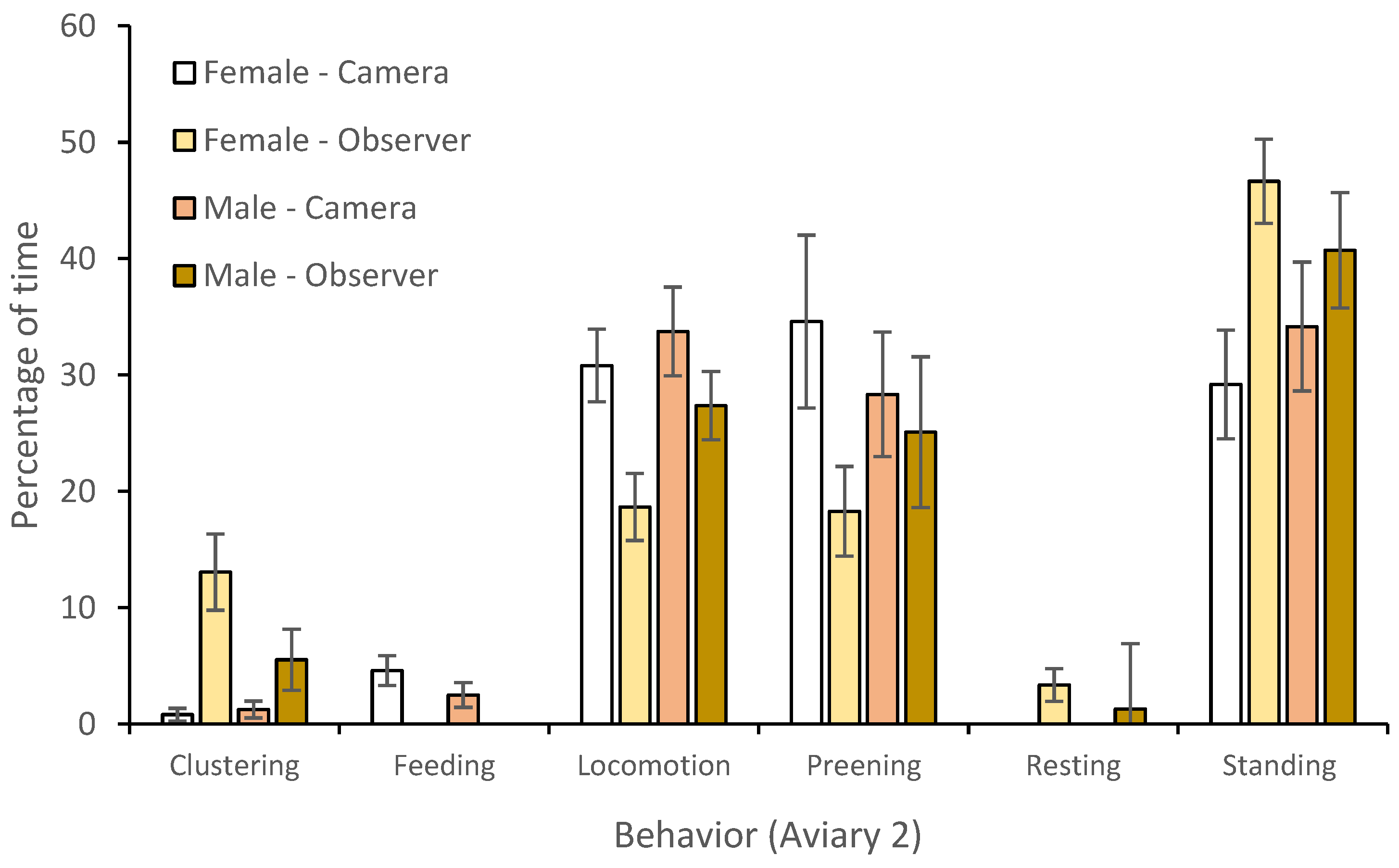
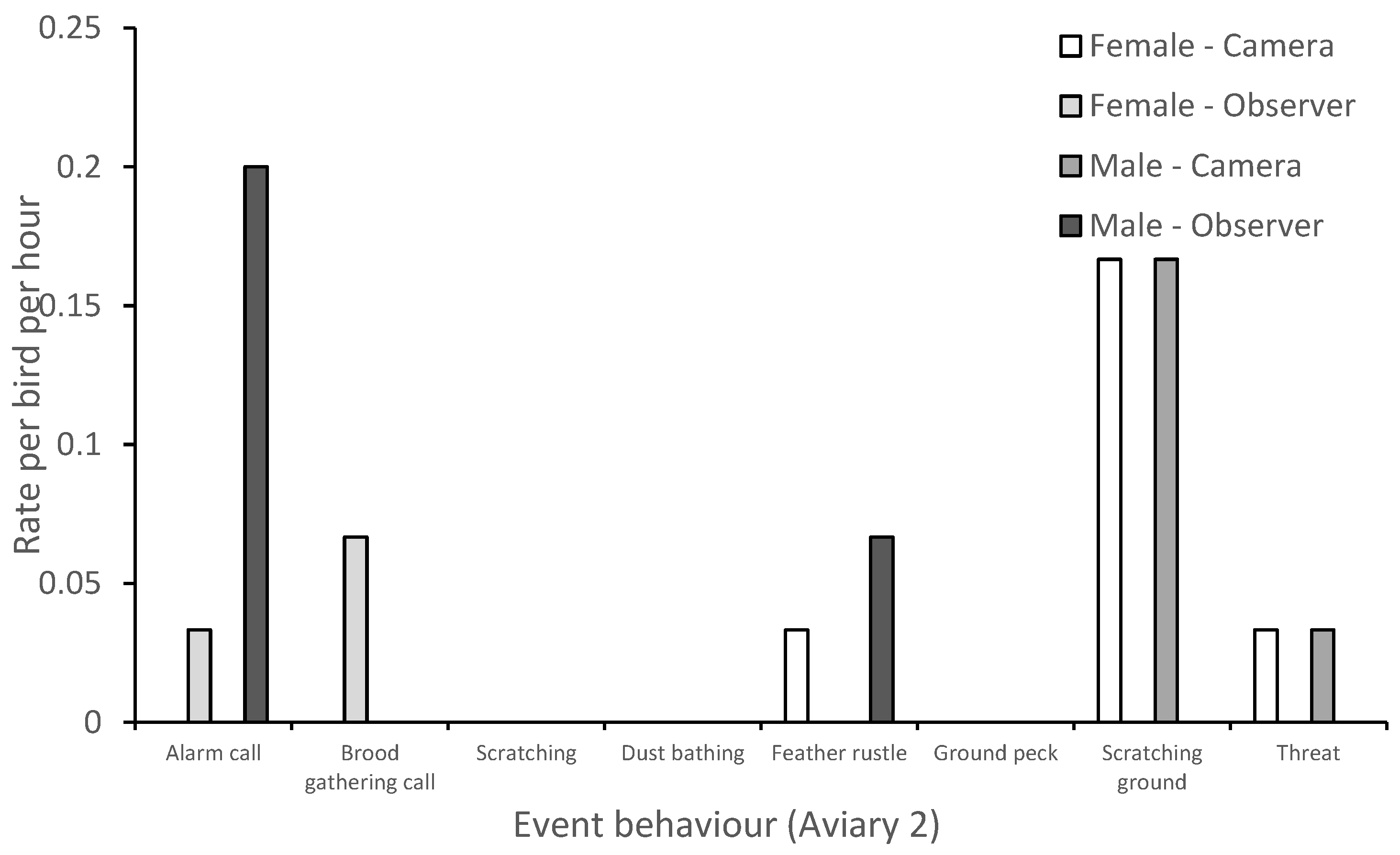
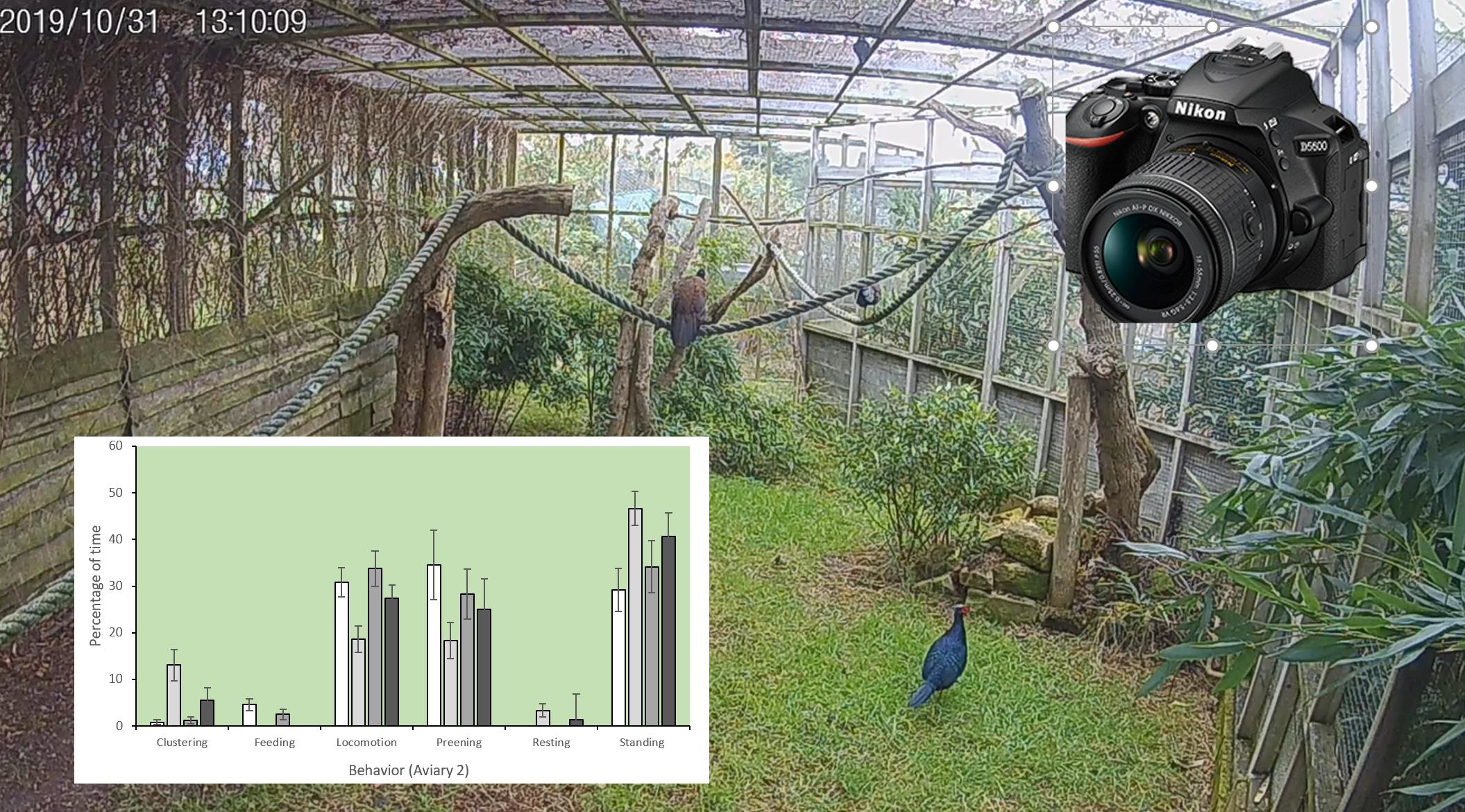
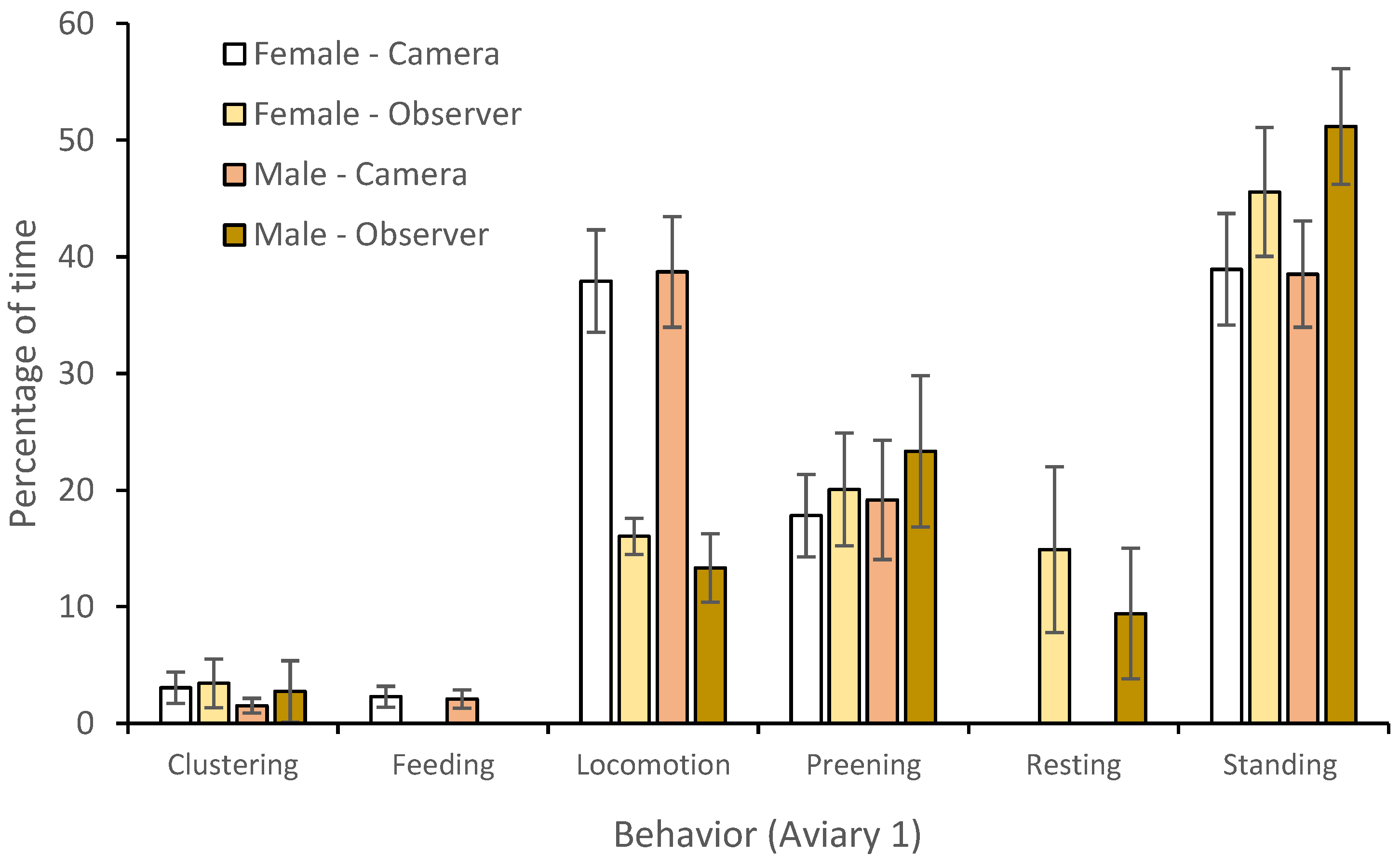
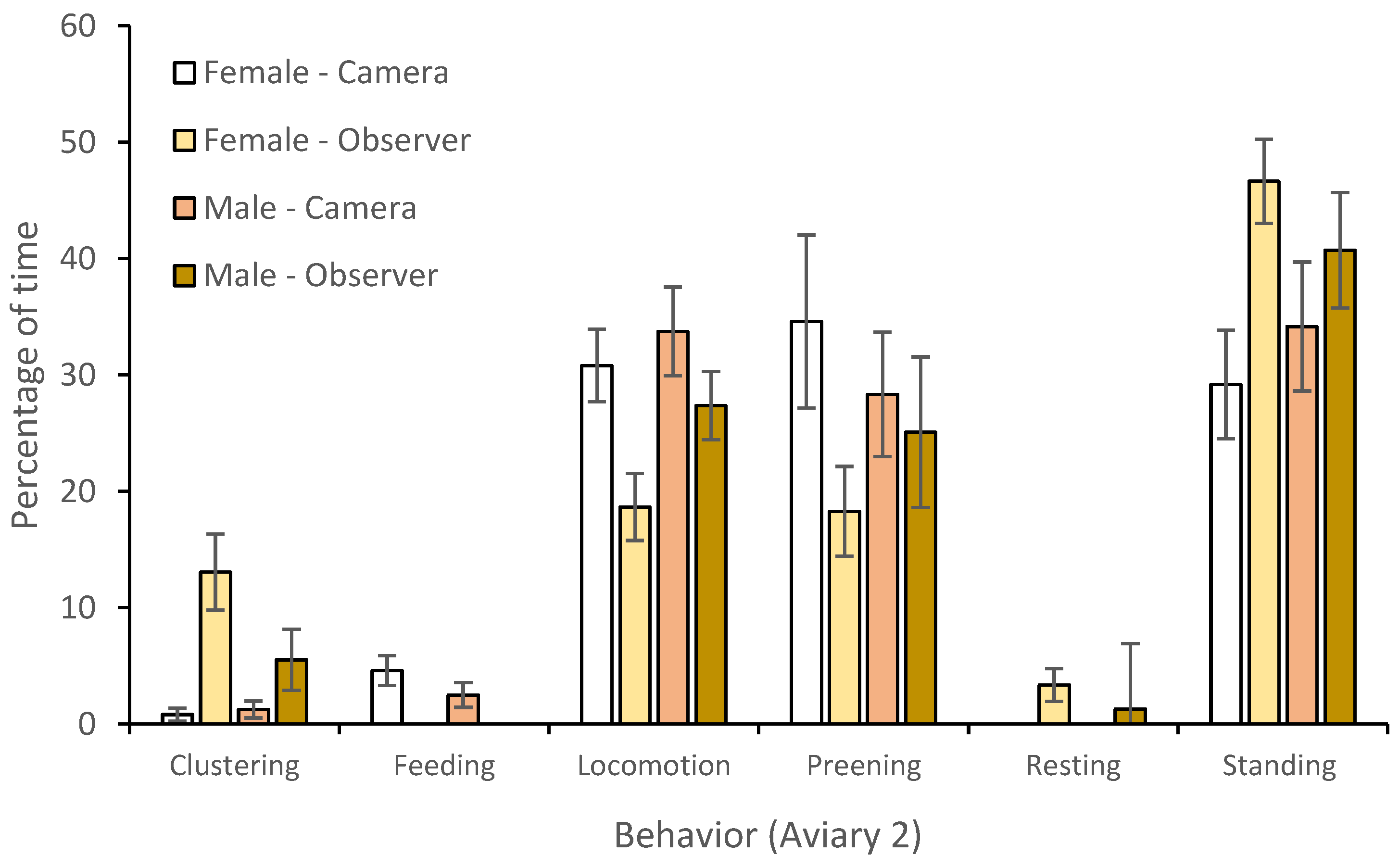
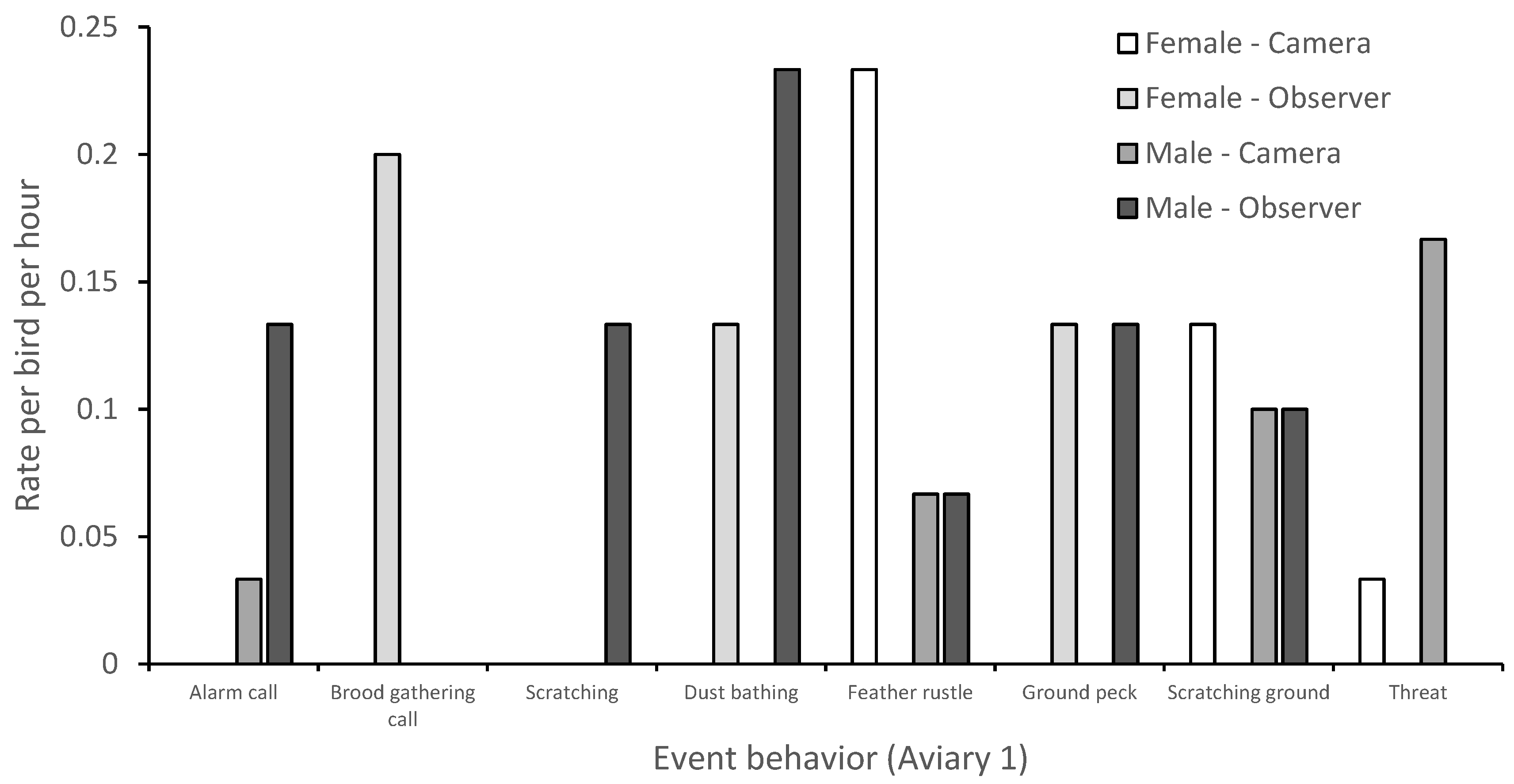
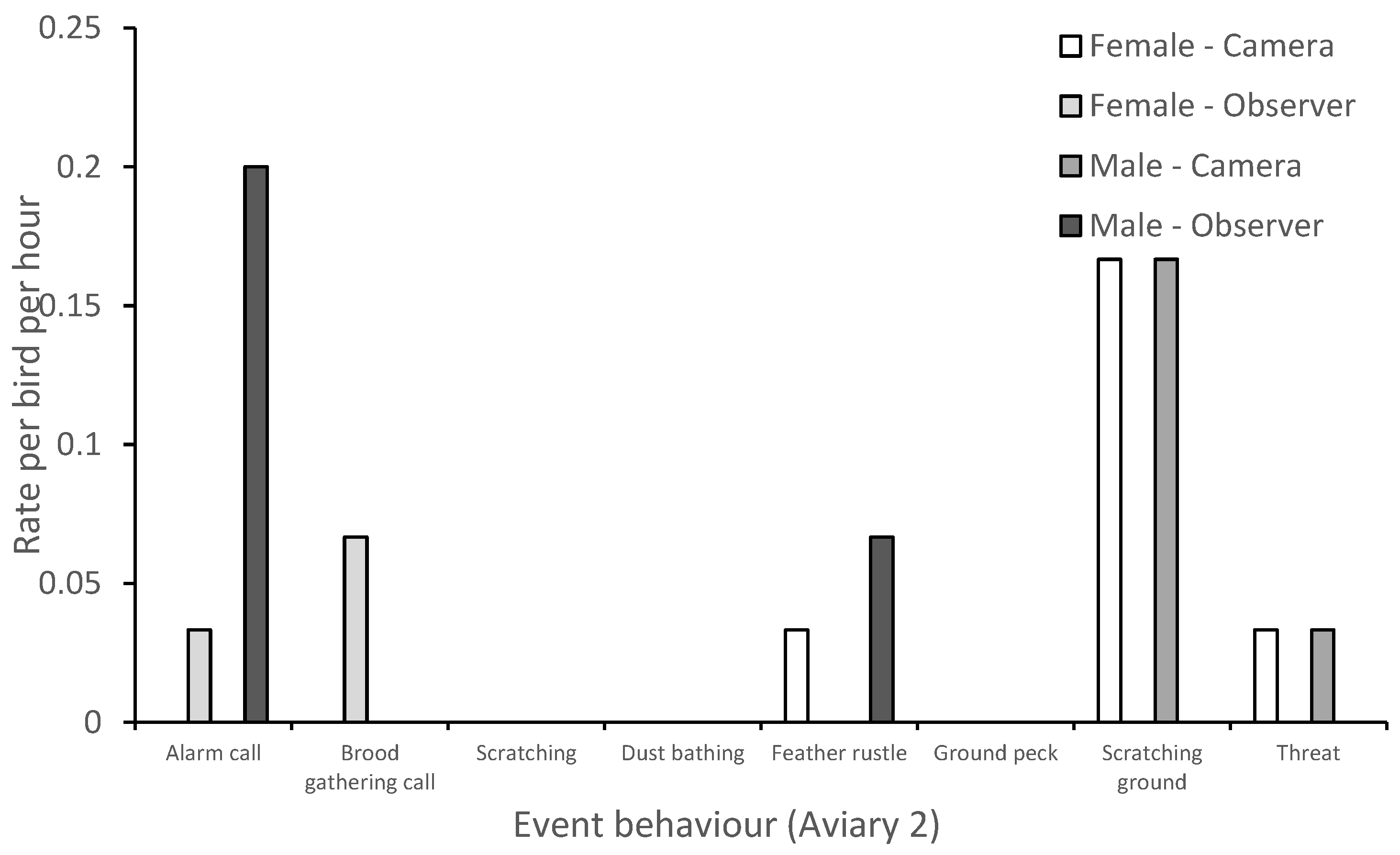
3.1. Behavior
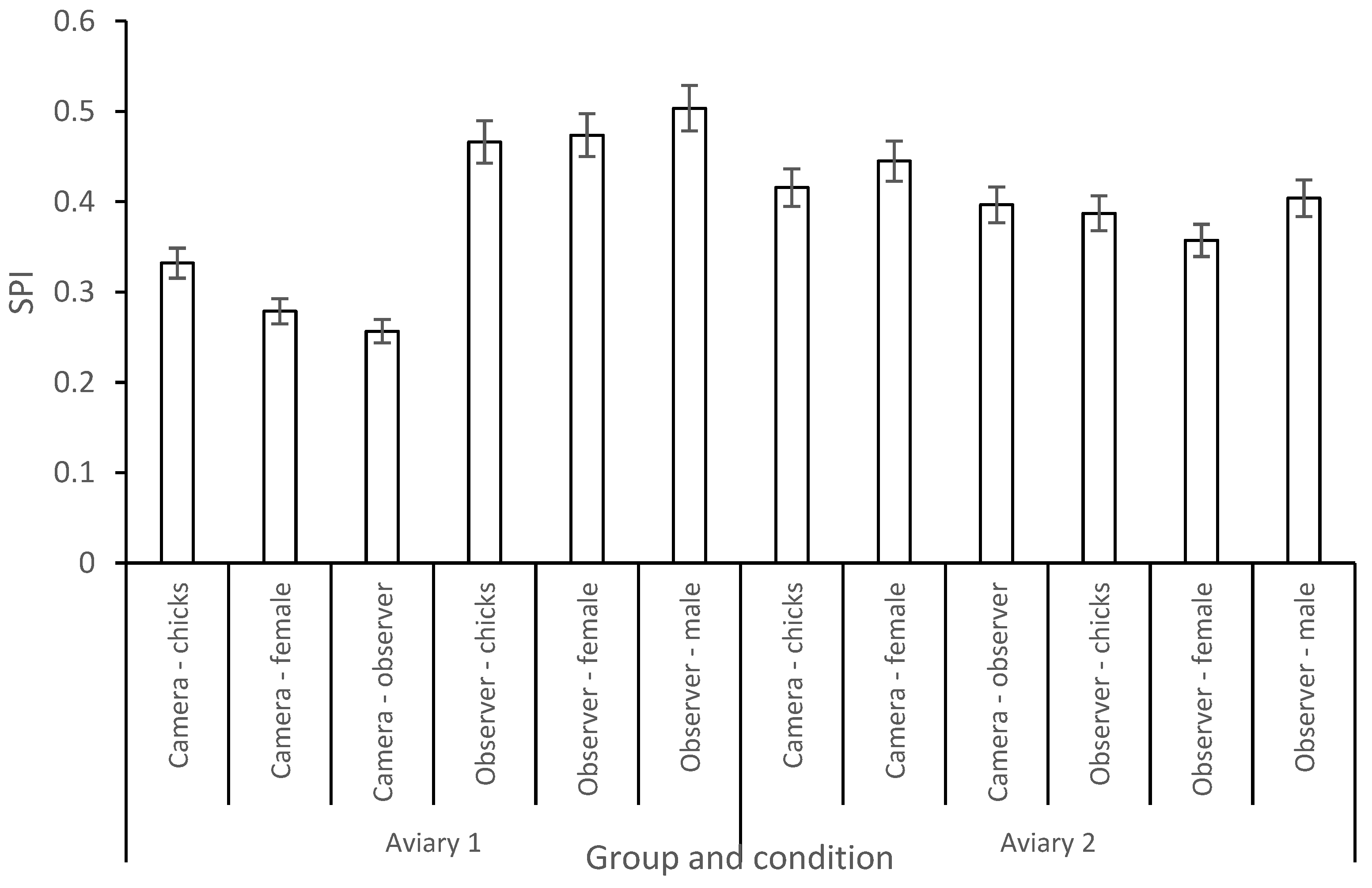
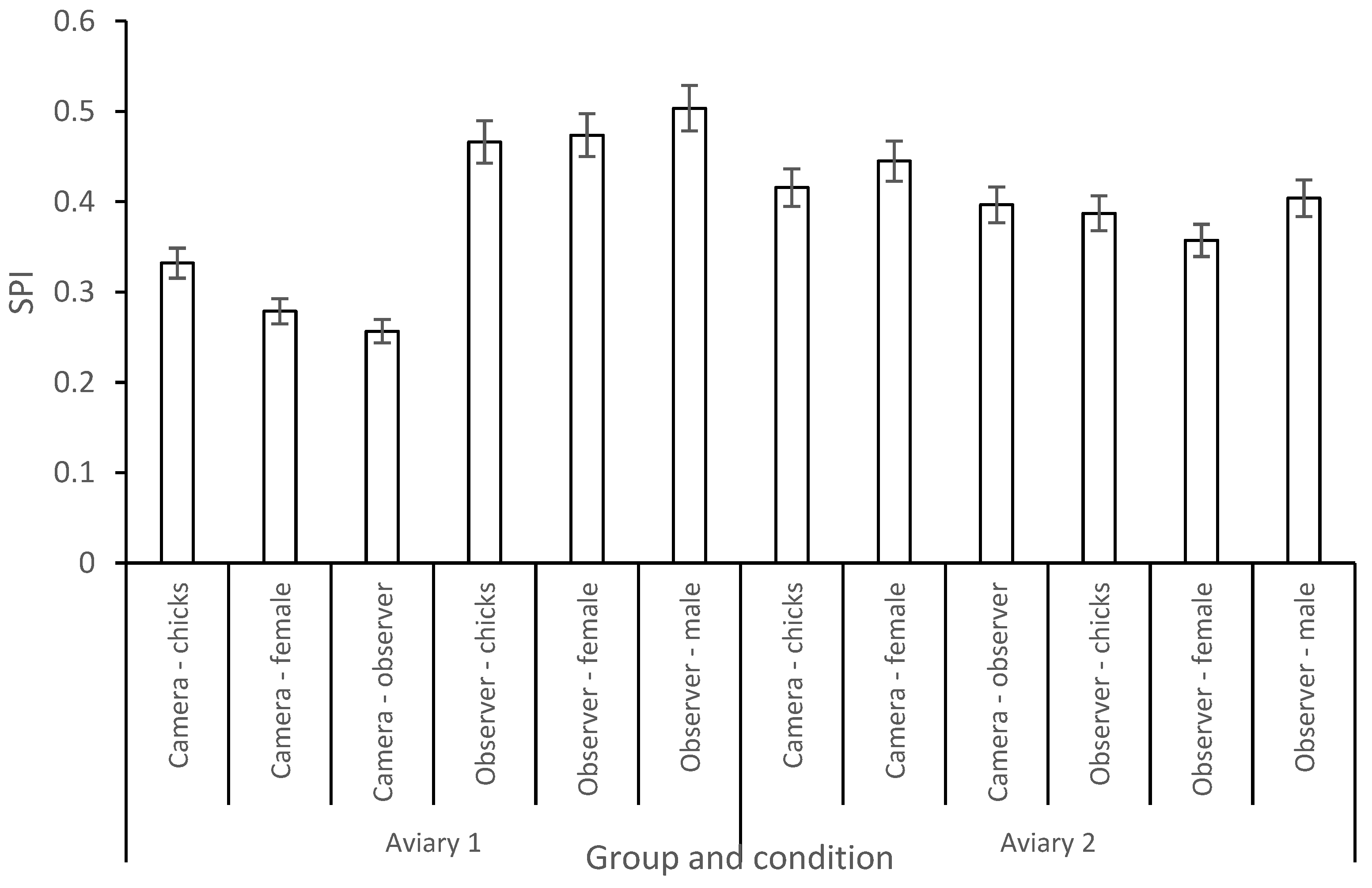
3.2. Enclosure Use
4. Discussion
4.1. Behavior
4.2. Enclosure Use
4.3. Future Directions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miller, L.J.; Mellen, J.D.; Kuczaj, S.A., II. The Importance of Behavioural Research in Zoological Institutions: An Introduction to the Special Issue. Int. J. Comp. Psychol. 2013, 26, 1–4. [Google Scholar] [CrossRef]
- Greggor, A.L.; Blumstein, D.T.; Wong, B.B.M.; Berger-Tal, O. Using animal behaviour in conservation management: A series of systematic reviews and maps. Environ. Evid. 2019, 8, 23. [Google Scholar] [CrossRef]
- Hutchins, M.; Thompson, S.D. Zoo and Aquarium Research: Priority Setting for the Coming Decade. Zoo Biol. 2008, 27, 488–497. [Google Scholar] [CrossRef] [PubMed]
- Berger, A. Activity patterns, chronobiology and the assessment of stress and welfare in zoo and wild animals. Int. Zoo Yearb. 2011, 45, 80–90. [Google Scholar] [CrossRef]
- Mallapur, A.; Sinha, A.; Waran, N. Influence of visitor presence on the behaviour of captive lion-tailed macaques (Macaca silenus) housed in Indian zoos. Appl. Anim. Welf. Sci. 2005, 94, 341–352. [Google Scholar] [CrossRef]
- Carrete, M.; Martínez-Padilla, J.; Rodríguez-Martínez, S.; Rebolo-Ifrán, N.; Palma, A.; Tella, J.L. Heritability of fear of humans in urban and rural populations of a bird species. Sci. Rep. 2016, 6, 31060. [Google Scholar] [CrossRef] [Green Version]
- Cooper, W.E.J.; Frederick, W.G. Optimal flight initiation distance. J. Theor. Biol. 2007, 244, 59–67. [Google Scholar] [CrossRef]
- Mitchell, H.; Hosey, G. Zoo Research Guidelines: Studies on the Effects of Human Visitors on Zoo Animal Behaviour. 2005. Available online: https://winghamwildlifepark.co.uk/wp-content/uploads/2012/12/Visitor-Effects-on-Animal-behaviour.pdf (accessed on 20 December 2021).
- Iredale, S.K.; Nevill, C.H.; Lutz, C.K. The Influence of Observer Presence on Baboon (Papio spp.) and Rhesus Macaque (Macaca mulatta) Behaviour. Appl. Anim. Behav. Sci. 2009, 122, 53–57. [Google Scholar] [CrossRef] [Green Version]
- Crofoot, M.C.; Lambert, T.D.; Kays, R.; Wikelski, M.C. Does watching a monkey change its behaviour? Quantifying observer effects in habituated wild primates using automated radiotelemetry. Anim. Behav. 2010, 80, 475–480. [Google Scholar] [CrossRef] [Green Version]
- Gray, J. Zoo Ethics: The Challenges of Compassionate Conservation; Cornell University Press: New York, NY, USA, 2017. [Google Scholar]
- Leruste, H.; Bokkers, E.A.M.; Sergent, O.; Wolthius-Fillerup, M.; Van Reenen, C.G.; Lensink, B.J. Effects of the observation method (direct v. from video) and the presence of an observer on behavioural results in veal calves. Animal 2013, 7, 1858–1864. [Google Scholar] [CrossRef]
- Wolfensohn, S.; Shotton, J.; Bowley, H.; Davies, S.; Thompson, S.; Justice, W.S.M. Assessment of Welfare in Zoo Animals: Towards Optimum Quality of Life. Animals 2018, 8, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, M. The impact of human disturbance on birds: A selective review. In Too Close for Comfort: Contensious Issues in Human-Wildlife Encounters; Lunney, D., Munn, A., Meikle, W., Eds.; Royal Zoological Society of New South Wales: Sydney, Australia, 2008; pp. 163–196. [Google Scholar]
- Bračko, A.; King, C.E. Advantages of aviaries and the Aviary Database Project: A new approach to an old housing option for birds. Int. Zoo Yearb. 2014, 48, 166–183. [Google Scholar] [CrossRef]
- McDougall, P.T. Is passive observation of habituated animals truly passive? J. Ethol. 2007, 30, 219–223. [Google Scholar] [CrossRef]
- De Azevedo, C.S.; Fontes, M.; Lima, F.; Da Silva, V.C.A.; Young, R.J.; Rodrigues, M. Visitor Influence on the Behaviour of Captive Greater Rheas (Rhea americana, Rheidae Aves). J. Appl. Anim. Welf. Sci. 2012, 15, 113–125. [Google Scholar] [CrossRef]
- Blanchett, M.K.S.; Finegan, E.; Atkinson, J. The Effects of Increasing Visitor and Noise Levels on Birds Within a Free-Flight Aviary Examined Through Enclosure Use and Behaviour. Anim. Behav. Cogn. 2011, 7, 49–69. [Google Scholar] [CrossRef]
- Morgan, K.N.; Tromborg, C.T. Sources of stress in captivity. Appl. Anim. Welf. Sci. 2007, 102, 262–302. [Google Scholar] [CrossRef]
- BirdLife International. Lophura edwardsi. 2018. Available online: https://www.iucnredlist.org/species/45354985/129928203#population (accessed on 15 February 2022).
- Species360 Zoological Information Management System. Lophura edwardsi. 2021. Available online: https://zims.species360.org/Login.aspx?ReturnUrl=%2f (accessed on 15 February 2022).
- Johnsgard, P.A. Pheasants of the World; Swan Hill Press: Shrewsbury, UK, 1999. [Google Scholar]
- Hennache, A. Husbandry Guidelines Lophura edwardsi; EAZA: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Hennache, A.; Mahood, S.P.; Eames, J.C.; Randi, E. Lophura hatinhensis is an invalid taxon. Forktail 2012, 28, 129–135. [Google Scholar]
- Martin, P.; Bateson, P. Measuring Behaviour: An Introductory Guide, 3rd ed.; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Chen, J.M.; Schütz, K.E.; Tucker, C.B. Technical Note: Comparison of instantaneous sampling and continuous observation of dairy cattle behaviour in freestall housing. J. Dairy Sci. 2016, 99, 8341–8346. [Google Scholar] [CrossRef]
- Dawkins, M.S. Observing Animal Behaviour: Design and Analysis of Quantitative Data; Oxford University Press: Oxford, UK, 2007. [Google Scholar]
- Katajamaa, R.; Larsson, L.H.; Lundberg, P.; Sörensen, I.; Jensen, P. Activity, social and sexual behaviour in Red Junglefowl selected for divergent levels of fear of humans. PLoS ONE 2018, 13, e0204303. [Google Scholar] [CrossRef]
- Zapletal, D.; Suchý, P.; Straková, E.; Vitula, F.; Kuchtík, J. Behaviour patterns of the cage-housed breeding flock of pheasants (Phasianus colchicus). Acta Univ. Agric. Silvic. Mendel. Brun. 2007, 28, 215–220. [Google Scholar] [CrossRef] [Green Version]
- Brereton, J.E. Current directions in animal enclosure use studies. J. Zoo Aquar. Res. 2020, 8, 1–9. [Google Scholar]
- Plowman, A. A note on a modification of the spread of participation index allowing for unequal zones. Appl. Anim. Welf. Sci. 2003, 83, 331–336. [Google Scholar] [CrossRef]
- Lund, A.; Lund, M. Descriptive and Inferential Statistics. 2018. Available online: https://statistics.laerd.com/statistical-guides/descriptive-inferential-statistics.php (accessed on 15 February 2022).
- Melfi, V. There are big gaps in our knowledge, and thus approach, to zoo animal welfare: A case for evidence-based zoo animal management. Zoo Biol. 2009, 28, 574–588. [Google Scholar] [CrossRef] [PubMed]
- World Pheasant Association. A Philosophy for the Reintroduction of Pheasants. WPA News 2015, 96, 12–13. [Google Scholar]
- Davey, G. Visitors’ effects on the welfare of animals in the zoo: A review. J. Appl. Anim. Welf. Sci. 2007, 10, 169–183. [Google Scholar] [CrossRef]
- European Conservation Breeding Group. Edwards’s Pheasant. WPA News 2014, 95, 12–13. [Google Scholar]
- Christiansen, F.; Rasmussen, M.H.; Lusseau, D. Inferring activity budgets in wild animals to estimate the consequences of disturbance. Behav. Ecol. 2013, 24, 1415–1425. [Google Scholar] [CrossRef] [Green Version]
- Baugh, A.T.; Davidson, S.C.; Hau, M.; Van Oers, K. Temporal dynamics of the HPA axis linked to exploratory behavior in a wild European songbird (Parus major). Gen. Comp. Endocrin. 2017, 250, 104–112. [Google Scholar] [CrossRef]
- Goodenough, A.E.; McDonald, K.; Moody, K.; Wheeler, C. Are “visitor effects” overestimated? Behaviour in captive lemurs is mainly driven by co-variation with time and weather. J. Zoo Aquar. Res. 2019, 7, 59–66. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aviary | Sex | Life Stage | Hatch Location |
|---|---|---|---|
| 1 | Male | Adult | Private Collection |
| 1 | Female | Adult | Private Collection |
| 1 | Female | Juvenile | Sparsholt College |
| 1 | Female | Juvenile | Sparsholt College |
| 1 | Female | Juvenile | Sparsholt College |
| 1 | Female | Juvenile | Sparsholt College |
| 1 | Male | Juvenile | Sparsholt College |
| 1 | Male | Juvenile | Sparsholt College |
| 2 | Male | Adult | Private Collection |
| 2 | Female | Adult | Private Collection |
| 2 | Female | Juvenile | Sparsholt College |
| 2 | Female | Juvenile | Sparsholt College |
| Behavior | Description |
|---|---|
| State behaviors | |
| Clustering | Birds get together and remain near to one another. |
| Feeding | Individual feeds from a trough, scratches litter back with feet and pecks the ground or pecks plants which are elevated from the ground. Includes the act of ingesting water or manipulating a water source. |
| Locomotion | Bird takes two or more steps, moving from one point to another. This action may be performed at a slow or fast pace, with head held forward. Behavior also includes flying; individual flaps wings to gain height from the ground and has all limbs off the ground for a period longer than two seconds or glides with wings stationary in mid-flight. |
| Preening | Cleaning of the feathers; moving beak across the body and individually through feathers (perhaps trimming or re-arranging feathers) in a standing or lying position. |
| Resting | Individual shows a relaxed posture, standing, sitting or lying down. Head may be close to the body. Includes sleeping and eyes closed for longer than two seconds. |
| Standing | Bird remains still in an upright position with legs extended. |
| Out of sight | Bird is not visible or is partially obstructed from observer’s vision. |
| Event behaviors | |
| Alarm call | Occurs when a threat or unusual sound/sight is present. Individuals repeat the call back to one another at approximately a ½ second apart until call slowly diminishes. |
| Brood gathering call | Can occur alongside a brood caution call. Vocal sounds include the parent clucking, barking, and “kee kee” noises. Chicks are silent and begin cheeping while returning to mother. |
| Chase | Aggressor runs toward opponent and opponent flees. |
| Dust bathing | Individual rolls in or covers body in dust/ground substrate to bathe. |
| Feather rustle | Bird shakes body laterally and causes feathers to appear larger and untidy. |
| Ground peck | Individual pecks ground with beak, once or more than once. |
| Scratching ground | Individual moves feet back and forth to scrape the ground, enclosure walls or feed trough. |
| Threat | Aggressor steps forward and lunges towards opponent. Opponent may flee but aggressor will not chase. Alternatively, aggressor may flap wings at opponent and may also produce a hoarse “krrrrah” sound. |
| Wing flapping | Bird extends wings and repeatedly moves them back and forth whilst standing, sitting, or lying down. |
| Behavior | R2 (P) | Predictor | SE Predictor | X2 | DF | p Value |
|---|---|---|---|---|---|---|
| Clustering | 25.74 | Model | 0.529 | 81.48 | 7 | <0.001 * |
| Temperature | 0.013 | 0.76 | 1 | 0.383 | ||
| Visitor presence (min) | 0.015 | 12.01 | 1 | 0.001 * | ||
| Keeper presence (min) | 0.004 | 0.35 | 1 | 0.557 | ||
| Individual bird | 0.109 | 1.54 | 3 | <0.214 | ||
| Observation type | 0.071 | 69.05 | 1 | <0.001 * | ||
| Feeding | 41.91 | Model | 0.636 | 18.83 | 7 | 0.004 |
| Temperature | 0.052 | 1.85 | 1 | 0.174 | ||
| Visitor presence (min) | 0.127 | 1.78 | 1 | 0.183 | ||
| Keeper presence (min) | 0.038 | 4.64 | 1 | 0.031 * | ||
| Individual bird | 0.283 | 0.08 | 3 | 0.774 | ||
| Observation type | 1.09 | 12.02 | 1 | 0.002 * | ||
| Locomotion | 36.98 | Model | 0.136 | 135.99 | 7 | <0.001 * |
| Temperature | 0.001 | 0.05 | 1 | 0.828 | ||
| Visitor presence (min) | 0.0158 | 1.76 | 1 | 0.185 | ||
| Keeper presence (min) | 0.005 | 15.71 | 1 | <0.001 * | ||
| Individual bird | 0.065 | 33.43 | 3 | <0.001 * | ||
| Observation type | 0.102 | 30.35 | 1 | <0.001 * | ||
| Preening | 6.07 | Model | 0.153 | 41.04 | 7 | <0.001 * |
| Temperature | 0.013 | 0.49 | 1 | 0.485 | ||
| Visitor presence (min) | 0.015 | 0.06 | 1 | 0.809 | ||
| Keeper presence (min) | 0.004 | 8.62 | 1 | 0.003 * | ||
| Individual bird | 0.109 | 28.63 | 3 | <0.001 * | ||
| Observation type | 0.071 | 2.66 | 1 | 0.265 | ||
| Resting | 42.27 | Model | 0.842 | 56.26 | 7 | <0.001 * |
| Temperature | 0.039 | 10.96 | 1 | 0.001 * | ||
| Visitor presence (min) | 0.307 | 21.72 | 1 | <0.001 * | ||
| Keeper presence (min) | 0.009 | 1.34 | 1 | 0.246 | ||
| Individual bird | 0.185 | 1.98 | 3 | 0.159 | ||
| Observation type | 842 | 34.84 | 1 | <0.001 * | ||
| Standing | 16.32 | Model | 0.122 | 65.06 | 7 | <0.001 * |
| Temperature | 0.122 | 6.27 | 1 | 0.012 * | ||
| Visitor presence (min) | 0.009 | 1.45 | 1 | 0.229 | ||
| Keeper presence (min) | 0.011 | 4.55 | 1 | 0.033 * | ||
| Individual bird | 0.003 | 28.46 | 3 | <0.001 * | ||
| Observation type | 0.050 | 2.84 | 1 | 0.241 |
| R2 | Predictor | SE Predictor | DF | p Value |
|---|---|---|---|---|
| 5.5 | Model | 0.001 | 9 | 0.014 * |
| Temperature | 0.005 | 1 | 0.013 * | |
| Visitor presence (min) | 0.005 | 1 | 0.686 | |
| Keeper presence (min) | 0.001 | 1 | 0.215 | |
| Individual bird | 0.039 | 5 | 0.498 | |
| Observation type | 0.031 | 1 | 0.001 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoy, R.A.; Brereton, J.E. Does Observer Presence Modify the Behavior and Enclosure Use of Captive Edwards’ Pheasants? J. Zool. Bot. Gard. 2022, 3, 147-157. https://doi.org/10.3390/jzbg3020012
Hoy RA, Brereton JE. Does Observer Presence Modify the Behavior and Enclosure Use of Captive Edwards’ Pheasants? Journal of Zoological and Botanical Gardens. 2022; 3(2):147-157. https://doi.org/10.3390/jzbg3020012
Chicago/Turabian StyleHoy, Rhiannon Amy, and James Edward Brereton. 2022. "Does Observer Presence Modify the Behavior and Enclosure Use of Captive Edwards’ Pheasants?" Journal of Zoological and Botanical Gardens 3, no. 2: 147-157. https://doi.org/10.3390/jzbg3020012
APA StyleHoy, R. A., & Brereton, J. E. (2022). Does Observer Presence Modify the Behavior and Enclosure Use of Captive Edwards’ Pheasants? Journal of Zoological and Botanical Gardens, 3(2), 147-157. https://doi.org/10.3390/jzbg3020012