

Human Gliomedin and Ryanodine 3 Type Receptor Is the Key to Explain the Guillain Barre Syndrome in SARS-CoV-2 and Others Bacterial Related to SARS-CoV-2 Postinfection? A Molecular Mimicry Point of View



Abstract

1. Introduction
2. Materials and Methods
2.1. Review of the Main Microorganisms Related to GBS
2.2. Sequence Alignment—Blast
2.3. Antigenic Protein Analysis
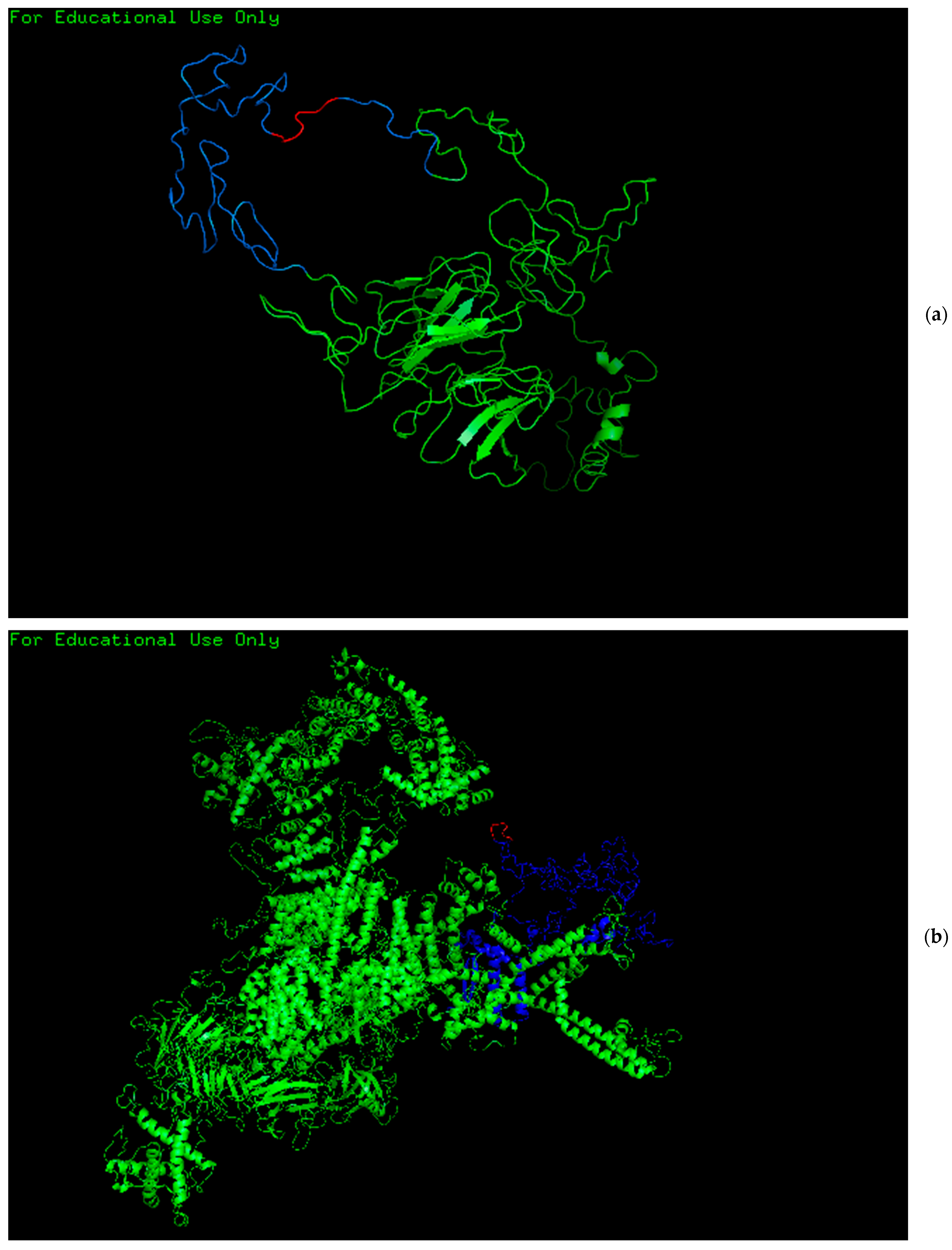
2.4. Protein Modeling
2.5. Prediction of B Cell, Citotoxic T Lymphocytes (CTL) and Helper T Lymphocytes (HTL) Epitopes
2.6. Prediction of the 3D Structures of the Predicted Epitope and HLA-A 0201 Allele Molecular Docking
3. Results
3.1. Sequence Alignment—Blast
3.2. Antigenicity Prediction
3.3. The CTL and HTL Epitopes
3.4. Prediction of the 3D Structures of the Predicted Epitope and HLA-A 0201 and HLA DR52c Allele Molecular Docking
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Esposito, S.; Longo, M.R. Guillain-Barre syndrome. Autoimmun. Rev. 2017, 16, 96–101. [Google Scholar] [CrossRef]
- Willison, H.J.; Jacobs, B.C.; van Doorn, P.A. Guillain-Barre syndrome. Lancet 2016, 388, 717–727. [Google Scholar] [CrossRef]
- Sookaromdee, P.; Wiwanitkit, V. Antibodies to dengue, Zika, Campylobacter jejuni and gangliosides in Guillain-Barre syndrome. Neurol. India 2019, 67, 335. [Google Scholar] [CrossRef]
- Takahashi, R.; Yuki, N. Streptococcal IdeS: Therapeutic potential for Guillain-Barre syndrome. Sci. Rep. 2015, 5, 10809. [Google Scholar] [CrossRef]
- Lewczuk, P.; Padilla-Docal, B.; Rodríguez-Rey, A.; Noris-García, E.; Coifiu-Fanego, R.B.; González-Hernández, M.; Sánchez-Martínez, C.; Dorta-Contreras, A.J. sICAM-1 intrathecal synthesis and release during the acute phase in children suffering from Coxsackie A9 and S. pneumoniae meningoencephalitis. Arq. Neuropsiquiatr. 2008, 66, 504–508. [Google Scholar]
- Restrepo-Jimenez, P.; Rodriguez, Y.; Gonzalez, P.; Chang, C.; Gershwin, M.E.; Anaya, J.M. The immunotherapy of Guillain-Barre syndrome. Expert Opin. Biol. Ther. 2018, 18, 619–631. [Google Scholar] [CrossRef]
- Jo, Y.S.; Choi, J.Y.; Chung, H.; Kim, Y.; Na, S.J. Recurrent Guillain-Barre Syndrome Following Urinary Tract Infection by Escherichia coli. J. Korean Med. Sci. 2018, 33, e29. [Google Scholar] [CrossRef]
- Toscano, G.; Palmerini, F.; Ravaglia, S.; Ruiz, L.; Invernizzi, P.; Cuzzoni, M.G.; Franciotta, D.; Baldanti, F.; Daturi, R.; Postorino, P.; et al. Guillain-Barré Syndrome Associated with SARS-CoV-2. N. Engl. J. Med. 2020, 382, 2574–2576. [Google Scholar] [CrossRef]
- Arnaud, S.; Budowski, C.; Ng Wing Tin, S.; Degos, B. Post SARS-CoV-2 Guillain-Barré syndrome. Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2020, 131, 1652–1654. [Google Scholar] [CrossRef]
- Finsterer, J.; Scorza, F.A.; Fiorini, A.C. SARS-CoV-2–associated Guillain-Barre syndrome in 62 patients. Eur. J. Neurol. 2021, 2025, 28. [Google Scholar] [CrossRef]
- Wang, D.; Hu, B.; Hu, C.; Zhu, F.; Liu, X.; Zhang, J.; Wang, B.; Xiang, H.; Cheng, Z.; Xiong, Y.; et al. Clinical Characteristics of 138 Hospitalized Patients with 2019 Novel Coronavirus-Infected Pneumonia in Wuhan, China. JAMA Neurol. 2020, 323, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Mao, L.; Jin, H.; Wang, M.; Hu, Y.; Chen, S.; He, Q.; Chang, J.; Hong, C.; Zhou, Y.; Wang, D.; et al. Neurologic Manifestations of Hospitalized Patients With Coronavirus Disease 2019 in Wuhan, China. JAMA Neurol. 2020, 77, 683–690. [Google Scholar] [CrossRef]
- Dreyfus, D.H.; Farina, A.; Farina, G.A. Molecular mimicry, genetic homology, and gene sharing proteomic “molecular fingerprints” using an EBV (Epstein-Barr virus)-derived microarray as a potential diagnostic method in autoimmune disease. Immunol. Res. 2018, 66, 686–695. [Google Scholar] [CrossRef] [PubMed]
- Obando-Pereda, G.A. GAKG-RGEKG an Epitope That Provokes Immune Cross-Reactivity between Prevotella sp. and Human Collagen: Evidence of Molecular Mimicry in Chronic Periodontitis. Autoimmune Dis. 2016, 2016, 5472320. [Google Scholar] [PubMed]
- Rojas, M.; Restrepo-Jiménez, P.; Monsalve, D.M.; Pacheco, Y.; Acosta-Ampudia, Y.; Ramírez-Santana, C.; Leung, P.S.C.; Ansari, A.A.; Gershwin, M.E.; Anaya, J.M. Molecular mimicry and autoimmunity. J. Autoimmun. 2018, 95, 100–123. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, K. Guillain-Barre syndrome during COVID-19 pandemic: An overview of the reports. Neurol. Sci. 2020, 41, 3149–3156. [Google Scholar] [CrossRef]
- Sudo, M.; Miyaji, K.; Spath, P.J.; Morita-Matsumoto, K.; Yamaguchi, Y.; Yuki, N. Polyclonal IgM and IgA block in vitro complement deposition mediated by anti-ganglioside antibodies in autoimmune neuropathies. Int. Immunopharmacol. 2016, 40, 11–15. [Google Scholar] [CrossRef]
- Kuwabara, S. Guillain-Barre syndrome: Epidemiology, pathophysiology and management. Drugs 2004, 64, 597–610. [Google Scholar] [CrossRef]
- Ang, C.W.; Jacobs, B.C.; Laman, J.D. The Guillain-Barre syndrome: A true case of molecular mimicry. Trends Immunol. 2004, 25, 61–66. [Google Scholar] [CrossRef]
- Sievers, F.; Higgins, D.G. Clustal Omega for making accurate alignments of many protein sequences. Protein Sci. 2018, 27, 135–145. [Google Scholar] [CrossRef]
- Zhao, W.; Zhang, B.; Yu, J.; He, X.; Yang, F.; Liu, X.; Li, Y.; Liu, Y.; Xie, Q.; Yang, Z. Analysis of the Genome Sequence and Prediction of B-Cell Epitopes of the Envelope Protein of Middle East Respiratory Syndrome-Coronavirus. IEEEACM Trans. Comput. Biol. Bioinform. 2018, 15, 1344–1350. [Google Scholar]
- Yang, J.; Zhang, Y. Protein Structure and Function Prediction Using I-TASSER. Curr. Protoc. Bioinform. 2015. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; A Greenbaum, J.; Jespersen, M.C.; Jurtz, V.; Kim, H.; Sette, A.; Yan, Z.; Nielsen, M.; Andreatta, M.; Dhanda, S.K.; et al. IEDB-AR: Immune epitope database-analysis resource in 2019. Nucleic Acids Res. 2019, 47, W502–W506. [Google Scholar]
- Lamiable, A.; Thévenet, P.; Rey, J.; Vavrusa, M.; Derreumaux, P.; Tufféry, P. PEP-FOLD3: Faster de novo structure prediction for linear peptides in solution and in complex. Nucleic Acids Res. 2016, 44, W449–W454. [Google Scholar] [CrossRef]
- Kurcinski, M.; Badaczewska-Dawid, A.; Kolinski, M.; Kolinski, A.; Kmiecik, S. Flexible docking of peptides to proteins using CABS-dock. Protein Sci. 2020, 29, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Hou, T.; Feng, T.; Liu, H.; Li, D.; Chen, F.; Kang, Y.; Sun, H.; Zhu, F. HawkRank: A new scoring function for protein-protein docking based on weighted energy terms. J. Cheminform. 2017, 9, 66. [Google Scholar]
- Uchibori, A.; Chiba, A. Autoantibodies in Guillain-Barre Syndrome. Brain Nerve 2015, 67, 1347–1357. [Google Scholar]
- Ng, J.K.M.; Doppler, K.; Stengel, H.; Sommer, C.; Appeltshauser, L.; Meinl, E.; Grosskreutz, J. Neurofascin-155 IgM autoantibodies in patients with inflammatory neuropathies. J. Neurol. Neurosurg. Psychiatry 2018, 89, 1145–1151. [Google Scholar]
- Ebrahim Soltani, Z.; Rahmani, F.; Rezaei, N. Autoimmunity and cytokines in Guillain-Barré syndrome revisited: Review of pathomechanisms with an eye on therapeutic options. Eur. Cytokine Netw. 2019, 30, 1–14. [Google Scholar] [CrossRef]
- Koch, M.; Maertens, B.; Franzke, C.-W.; Hopkins, D.; Keene, D.R.; Bruckner-Tuderman, L.; Greenspan, D.S. Cleavage and oligomerization of gliomedin, a transmembrane collagen required for node of ranvier formation. J. Biol. Chem. 2007, 282, 10647–10659. [Google Scholar]
- Meissner, G. The structural basis of ryanodine receptor ion channel function. J. Gen. Physiol. 2017, 149, 1065–1089. [Google Scholar] [CrossRef] [PubMed]
- Kira, J.I.; Yamasaki, R.; Ogata, H. Anti-neurofascin autoantibody and demyelination. Neurochem. Int. 2019, 130, 104360. [Google Scholar] [CrossRef] [PubMed]
- Devaux, J.J. Antibodies to gliomedin cause peripheral demyelinating neuropathy and the dismantling of the nodes of Ranvier. Am. J. Pathol. 2012, 181, 1402–1413. [Google Scholar] [CrossRef] [PubMed]
- Querol, L.; Lleixà, C. Novel Immunological and Therapeutic Insights in Guillain-Barré Syndrome and CIDP. Neurotherapeutics 2021, 18, 2222–2235. [Google Scholar] [CrossRef]
- Nascimento, O.J.M.; da Silva, I.R.F. Guillain-Barre syndrome and Zika virus outbreaks. Curr. Opin. Neurol. 2017, 30, 500–507. [Google Scholar] [CrossRef]
- Cickovski, T.; Mondal, A.M.; Stebliankin, V.; Balbin, C.A.; Baral, P.; Chapagain, P.; Sobhan, M.; Siltberg-Liberles, J.; Nunez-Castilla, J.; Narasimhan, G.; et al. Potential Autoimmunity Resulting from Molecular Mimicry between SARS-CoV-2 Spike and Human Proteins. Viruses 2022, 14, 1415. [Google Scholar]
- Blundell, T.L.; Thomas, S.E.; Bannerman, B.P.; Vedithi, S.C.; Copoiu, L.; Hala, S.; Jamasb, A.R.; van Tonder, A.J.; Alsulami, A.F.; Beaudoin, C.A.; et al. Predicted structural mimicry of spike receptor-binding motifs from highly pathogenic human coronaviruses. Comput. Struct. Biotechnol. J. 2021, 19, 3938–3953. [Google Scholar]
- Rich, K.D.; Srivastava, S.; Muthye, V.R.; Wasmuth, J.D. Identification of potential molecular mimicry in pathogen-host interactions. PeerJ 2023, 11, e16339. [Google Scholar] [CrossRef]
- Bigdeli, A.; Ghaderi-Zefrehei, M.; Lesch, B.J.; Behmanesh, M.; Arab, S.S. Bioinformatics analysis of myelin-microbe interactions suggests multiple types of molecular mimicry in the pathogenesis of multiple sclerosis. PLoS ONE 2024, 19, e0308817. [Google Scholar] [CrossRef]
- Reynisson, B.; Alvarez, B.; Paul, S.; Peters, B.; Nielsen, M. NetMHCpan-4.1 and NetMHCIIpan-4.0: Improved predictions of MHC antigen presentation by concurrent motif deconvolution and integration of MS MHC eluted ligand data. Nucleic Acids Res. 2020, 48, W449–W454. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PUBMED Ref. | Protein | Epitope | Length |
|---|---|---|---|
| NP_861454.2 | Gliomedin | KGEKGD | 213–218 |
| NP_001027.3 | Ryanodine Receptor 3 | KGEKGD | 4269–4274 |
| WP_038434285.1 | S. pyogenes | KGEKGD | 163–168/201–206 |
| WP_050096714.1 | S. pneumoniae | KGEKGD | 600–605/615–620/651–656/669–674/759–764/789–794/819–824/947–952/1238–1243/1253–1258/1518–1523 |
| WP_096855986.1 | E. coli | KGEKGD | 330–305 |
| EAH8259323.1 | C. jejuni | KGEKGD | 32–37 |
| WP_041175161.1 | H.influenzae | KGEKG | 607–612 |
| Gliomedin | Ryanodine Type 3 Receptor | E. coli | ||||||
|---|---|---|---|---|---|---|---|---|
| Position | Residue | Index | Position | Residue | Index | Position | Residue | Index |
| 213 | Lys | 1.73 | 4269 | Lys | 1.65 | 330 | Lys | 3.4 |
| 214 | Gly | 1.74 | 4270 | Gly | 2.3 | 331 | Gly | 2.86 |
| 215 | Glu | 1.78 | 4271 | Glu | 3 | 332 | Glu | 2.83 |
| 216 | Lys | 2.12 | 4272 | Lys | 2.5 | 333 | Lys | 2.8 |
| 217 | Gly | 2.86 | 4273 | Gly | 2.4 | 334 | Gly | 2.57 |
| 218 | Asp | 3.4 | 4274 | Asp | 1.9 | 335 | Asp | 2.74 |
| S. neumoniae | ||||||||
| Position | Residue | Index | Position | Residue | Index | Position | Residue | Index |
| 600 | Lys | 2 | 759 | Lys | 2.7 | 1238 | Lys | 1.1 |
| 601 | Gly | 2.04 | 760 | Gly | 2.74 | 1239 | Gly | 1.64 |
| 602 | Glu | 2.08 | 761 | Glu | 2.58 | 1240 | Glu | 1.78 |
| 603 | Lys | 2.12 | 762 | Lys | 2.62 | 1241 | Lys | 2.32 |
| 604 | Gly | 2.46 | 763 | Gly | 2.66 | 1242 | Gly | 2.66 |
| 605 | Asp | 3.4 | 764 | Asp | 3.4 | 1243 | Asp | 3.4 |
| 615 | Lys | 1.5 | 789 | Lys | 1.5 | 1253 | Lys | 2.7 |
| 616 | Gly | 1.84 | 790 | Gly | 1.84 | 1254 | Gly | 2.74 |
| 617 | Glu | 1.92 | 791 | Glu | 1.98 | 1255 | Glu | 2.78 |
| 618 | Lys | 2.32 | 792 | Lys | 2.32 | 1256 | Lys | 2.68 |
| 619 | Gly | 2.66 | 793 | Gly | 2.66 | 1257 | Gly | 2.66 |
| 620 | Asp | 3.4 | 794 | Asp | 3.4 | 1258 | Asp | 3.4 |
| 651 | Lys | 2.7 | 819 | Lys | 1.5 | 1518 | Lys | 2.11 |
| 652 | Gly | 2.74 | 820 | Gly | 1.84 | 1519 | Gly | 2.18 |
| 653 | Glu | 2.58 | 821 | Glu | 1.98 | 1520 | Glu | 2.25 |
| 654 | Lys | 2.62 | 822 | Lys | 2.32 | 1521 | Lys | 2.32 |
| 655 | Gly | 2.66 | 823 | Gly | 2.66 | 1522 | Gly | 2.66 |
| 656 | Asp | 3.4 | 824 | Asp | 3.4 | 1523 | Asp | 3.4 |
| 669 | Lys | 1.5 | 947 | Lys | 1.1 | |||
| 670 | Gly | 1.84 | 948 | Gly | 1.4 | |||
| 671 | Glu | 1.98 | 949 | Glu | 1.78 | |||
| 672 | Lys | 2.32 | 950 | Lys | 2.12 | |||
| 673 | Gly | 2.66 | 951 | Gly | 2.46 | |||
| 674 | Asp | 3.4 | 952 | Asp | 3.4 | |||
| S. pyogenes | C. jejuni | |||||||
| Position | Residue | Index | Position | Residue | Index | Position | Residue | Index |
| 163 | Lys | 1.1 | 202 | Lys | 1.1 | 31 | Lys | 0.75 |
| 164 | Gly | 1.44 | 203 | Gly | 1.44 | 32 | Gly | 1.3 |
| 165 | Glu | 1.78 | 204 | Glu | 1.78 | 33 | Glu | 1.3 |
| 166 | Lys | 2.12 | 205 | Lys | 2.12 | 34 | Lys | 1.3 |
| 167 | Gly | 2.86 | 206 | Gly | 2.66 | 35 | Gly | 1.3 |
| 168 | Asp | 3.4 | 207 | Asp | 3.4 | 36 | Asp | 1.5 |
| H. influenzae | ||||||||
| Position | Residue | Index | ||||||
| 607 | Lys | 0.9 | ||||||
| 608 | Gly | 0.9 | ||||||
| 609 | Glu | 0.9 | ||||||
| 610 | Lys | 0.9 | ||||||
| 611 | Gly | 1.3 | ||||||
| S/N | Core Peptide | Length | Alleles MHCI | Alleles MHCII | Antigenicity | Conservancy | Toxicity | Allergenicity |
|---|---|---|---|---|---|---|---|---|
| 1 | KGEKGD | 6 | HLA-A*30:01 HLA-B*15:01 | HLA-DQA1*05:01/DQB1*03:01 HLA-DRB3*01:01 HLA-DRB4*01:01 | 2.8561 | 100 | Non-toxic | allergen |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Obando-Pereda, G.A.; Ponce-Soto, L.A. Human Gliomedin and Ryanodine 3 Type Receptor Is the Key to Explain the Guillain Barre Syndrome in SARS-CoV-2 and Others Bacterial Related to SARS-CoV-2 Postinfection? A Molecular Mimicry Point of View. Immuno 2025, 5, 28. https://doi.org/10.3390/immuno5030028
Obando-Pereda GA, Ponce-Soto LA. Human Gliomedin and Ryanodine 3 Type Receptor Is the Key to Explain the Guillain Barre Syndrome in SARS-CoV-2 and Others Bacterial Related to SARS-CoV-2 Postinfection? A Molecular Mimicry Point of View. Immuno. 2025; 5(3):28. https://doi.org/10.3390/immuno5030028
Chicago/Turabian StyleObando-Pereda, Gustavo Alberto, and Luis Alberto Ponce-Soto. 2025. "Human Gliomedin and Ryanodine 3 Type Receptor Is the Key to Explain the Guillain Barre Syndrome in SARS-CoV-2 and Others Bacterial Related to SARS-CoV-2 Postinfection? A Molecular Mimicry Point of View" Immuno 5, no. 3: 28. https://doi.org/10.3390/immuno5030028
APA StyleObando-Pereda, G. A., & Ponce-Soto, L. A. (2025). Human Gliomedin and Ryanodine 3 Type Receptor Is the Key to Explain the Guillain Barre Syndrome in SARS-CoV-2 and Others Bacterial Related to SARS-CoV-2 Postinfection? A Molecular Mimicry Point of View. Immuno, 5(3), 28. https://doi.org/10.3390/immuno5030028