Abstract
In terms of the global burden of disease, helminthiasis is the most common infectious disease in the world. In response to the disease, the human host develops an immunological response that occurs predominantly through the action of T helper 2 (Th2) cells and the interleukins IL-4, IL-5 and IL-13. However, other types of Th cells, such as Th9, are also involved in the defense against helminths, with the IL-9 produced by these cells promoting the induction of mastocytosis and the increased production of IgG1 and IgE, in addition to the increase in intestinal contractility that promotes the expulsion of worms. Together, IL-9 and IL-10, which is also produced by Th9, induce a type 2 inflammatory response characterized by the coordinated actions of innate lymphoid cells, mast cells, basophils and other cells that work together toward a single objective: the reduction of the parasitic burden. This review presents the latest findings on Th9 effector mechanisms in helminthic infections.
1. Introduction
Parasitic infections comprise a group of neglected diseases that are mainly caused by helminths and protozoa that colonize the gastrointestinal tracts of their hosts [1]. Helminthiasis comprises different types of helminth infections that are caused by different species of worms and are the most common infectious diseases in the world, with almost 240 million people infected with Schistosoma spp. [2] and 1.5 billion infected by soil-transmitted helminths, with the Americas particularly affected, with about one-third of the population carrying these parasites [3].
Classically, the immune response against helminths is predominantly due to the action of T helper 2 (Th2) cells and the interleukins IL-4, IL-5 and IL-13. In addition, innate immunity participates through the action of eosinophils, mast cells and basophils—which release their granules to destroy helminths—and activated macrophages, which have a tissue-repairing role through their expression of molecules such as resistin-like molecule (RELM) and arginase 1 [1,4].
The role of IL-9 in the defense against helminths through the induction of mastocytosis and the increased production of IgG1 and IgE, in addition to the increased intestinal contractility that promotes the expulsion of worms, was first discovered in the 1980s [5]. However, only in recent decades has it been shown that IL-9 is not only produced by Th2 cells but is also produced by T cells, such as regulatory T cells (Treg) [6] and CD8+ T cells, including natural killer and gamma-delta (γδ) T cells [7].
Additionally, studies were developed in order to identify which T helper (Th) cells produced IL-9, and it was concluded that IL-9 is produced by a specific group of Th cells that became known as Th9. These cells, characterized by their expression of the transcription factors PU.1 and IRF4 [8], require the induction of TGF-β, a key cytokine that promotes the reprogramming of Th2 cells to the Th9 phenotype [9]. In this review, the important inflammatory role of Th9 cells and IL-9 in the defense against helminths will be discussed.
2. The Participation of Th9 in Inflammation
The inflammatory response, or inflammation, is characterized as a complex cascade of physiological events which have the function of protecting the body’s tissues and organs by acting in such a way as to restrict local damage; however, an excessive response can have deleterious effects [10].
The immune response triggered by the presence of microorganisms and the subsequent tissue damage contribute to the elimination of these agents, as well as favoring the stimulation of repair processes [11]. During the inflammatory process, cellular infiltrate leukocytes may arise as pre- and pro-inflammatory molecules which can be stimulated or inhibited, collaborating in the control of the signs and symptoms of infection [12,13].
Among leukocytes, CD4+ T cells, also known as T helper lymphocytes, differentiate into different Th cell patterns, with different effector functions that drive the profiles of immune responses. CD4+ T cells have transcription factors which, in response to antigenic stimuli, define the signaling pathways responsible for the production of interleukins for each type of T cell [14].
Among the Th cell patterns, Th9 plays an important role in immunity against parasites and autoimmune and inflammatory processes wherein the differentiation of these cells is related to TGF-β and IL-4 signaling pathways, in combination with the participation of the transcription factors IRF4 and PU.1 [9,14,15,16,17,18]. The differentiation process begins with the activation of PU.1 by TGF-β, which triggers the activation of the IL-4 receptor. This induces STAT6, which is required for the secretion of IL-9. GATA-3 and IRF-4 are then activated, leading to the transcription and secretion of IL-9 [19] (Figure 1).
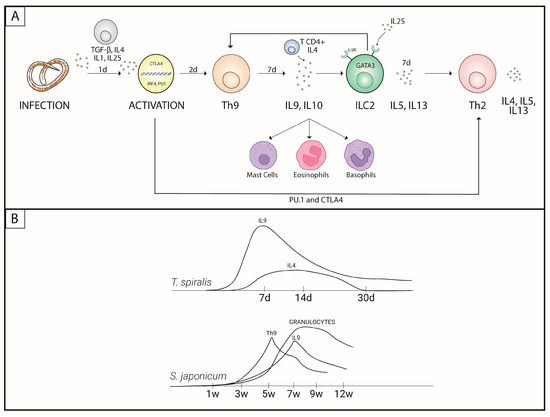
Figure 1.
Th9 response activation process. (A) Participation of Th9 cells in the activation of Th2 cells. (B) Time (in days (d) or weeks (w)) required for triggering the Th9 response in different infections.
Initially, IL-9 production was associated with the Th2 response, but studies carried out in mice have redefined IL-9-producing cells as Th9 cells [9,18]. This interleukin acts as a growth factor for mast cells and potentiates the production of inflammatory cytokines by these cells [20] (Figure 1). The secretion of IL-9, induced by TGFβ/IL-4 in CD4+ T cells, depends on the action of a complex of cytokines, such as IL-1β, IL-6, IL-10, IL-21, IL-25, IL-33, IFN-α and IFN-β, as well as the involvement of members of the TNF receptor superfamily (TNFR), which are activated by their ligands OX40L, TL1A and GITRL (capable of increasing IL-9) while being inhibited by IFNγ and IL-27 [21,22].
Studies that analyzed the dynamics of the immune response in allergic processes (similar to helminthic infections in terms of eosinophilic and mast cell responses) showed that the peak cytokine production for both IFN-γ and IL-9 was significantly higher than that of IL-4 [23]. However, Th1 inflammation does not become dominant in all allergic diseases, even when Th2 allergic inflammation is reduced, indicating that the pathogenesis is not entirely due to a Th1/Th2 imbalance [24] and leading to the conclusion that other mechanisms must be involved.
Furthermore, in a study involving allergic rhinitis (AR), exogenous IL-9 augmented IL-4 and exogenous IL-4 augmented IL-9, proving the process of feedback between Th2 and Th9 cytokines [23]. The Th2 responses were greater in AR mice than in controls and were downregulated by anti-IL-9 treatment, suggesting that in the absence of IL-9, the Th2 reactions are inhibited [24].
Some studies carried out in mice demonstrated that the overexpression of IL-9 leads to the development of intestinal mastocytosis and makes the animals resistant to infection by Trichuris muris while also causing the inflammation of airways [5,18,25,26,27]. Due to studies of this type, Th9 cells have been associated with immunological protection against parasitic infections by protozoa and helminths and extracellular pathogens [1].
3. Protective Potential of Th9 in Helminth Infections
3.1. Nematodes
For many years, it was believed that Th2 cells produced IL-9 due to the intrinsic participation of this interleukin in the Th2 response. Since the 1990s, studies have shown that the increases in IL-9 and IL-5 in hosts infected with Trichuris muris confer immunological resistance and prevent the spread of the infection due to a greater ability to expel the parasite [28].
Furthermore, the increases in parasite-specific IgG1 and total IgE, as well as intestinal eosinophils, were inversely proportional to the parasite load unlike parasite-specific IgG2a, which is produced and controlled by IFN-γ stimulation, which, when increased, was unable to control the number of worms [28]. This function is so effective with respect to providing protection against helminths that mice immunized with a synthetic self-adjuvant oleic-vinyl sulfone (OVS) linked to the catalytic region of recombinant serine/threonine phosphatase 2A from the nematode Angiostrongylus costaricensis (rPP2A) had reductions of up to 99% in the number of T. muris eggs [29].
This protective action was also associated with the ability of IL-9 to recruit mast cells [5]. Mast cells are cells originating in the bone marrow which, once mature, are located in mucous membranes or connective tissues. They are classified according to the location and content of their granules, which are composed of different proteases, depending on the environmental stimulus [30]. These cells are implicated in the defense against helminths by inducing a type 2 inflammatory response characterized by the presence of type 2 innate lymphoid cells (ILC2)—especially those producing IL-5, which are Th2, eosinophils, basophils and classically activated macrophages (M2), which are capable of promoting the expulsion of worms [31,32].
This same defense mechanism has been described for experimental infections with Nippostrongylus braziliensis. Additionally, it was reported by Licona-Limón et al. (2013) that IL-9 is the first cytokine expressed in response to the infection by this parasite, having been identified two days post infection, and that it is necessary for the production of IL-5 and IL-13 and increases in eosinophils and basophils in addition to controlling the parasitic load. Furthermore, it has been demonstrated that IL-9 is capable of protecting the host against infections through the stimulation of basophilia and mastocytosis that contribute to the rapid expulsion of adult worms. This is only possible due to the presence of IL-9 receptors (IL-9r mRNA) in these cell types [33].
One of the factors associated with mast cell activity is the enzyme carbonic anhydrase (Car) 1, which is characteristic of the hematopoietic progenitors that are found in peripheral tissues in a steady state and increase after stimulation, such as during infection with Trichinella spiralis [33,34]. In this type of infection, the participation of IL-25, an interleukin of the large IL-17 family, is fundamental for the differentiation of lymphocytes in Th9. This was proven by the increased presence of interleukin-17 receptor B (IL-17RB), the receptor for IL-25 in the gastrointestinal tract of mice infected with T. spiralis [35].
ILC2 induces an increased production of Th2 and Th9 cytokines via MHC-II-dependent interactions and is a primary producer of IL-5 and IL-13 in the early stages of infection [35]. Since IL-9 appears early in infections and stimulates the production of IL-5 and IL-13, it can be inferred that this interleukin is capable of stimulating ILC2 in a process of retrostimulation [33]. This was verified in the studies by Angkasekwinai et al. (2017) and Turner et al. (2013), who described the different roles of IL-9 in stimulating ILCs and vice versa [35,36].
Angkasekwinai et al. (2017) showed that the increase in IL-25 occurred on the seventh day post-infection and that it induced robust increases in IL-5 and IL-13 but only a moderate increase in IL-9, which was produced by mesenteric lymph node cells from infected mice. ILC2 expresses greater amounts of MHC-II, and their activation potentiates the CD4+ T cell response. However, despite continuing to have the ability to produce IL-4, IL-9 and IL-13 during CD4+ T cell depletion, ILC responds poorly to IL-25, showing that CD4+ T cells stimulate ILCs but are not essential for their activity. Furthermore, the parasite load in mice treated with anti-CD4 remained high after 14 days of infection, again indicating the importance of these cells for controlling parasitism [35,37]. In addition, the study showed that the Th9 response is more efficient in reducing the parasite load than the Th2 response, but it is even more effective in combination. Finally, the study suggested that the IL-25 receptor signal is required for the accumulation of ILC2s during infection which, in turn, may enable CD4+ T cells to exhibit strong Th2 and Th9 antigen-specific cytokine responses against T. spiralis [35,37].
However, Turner et al. (2013) showed that IL-9 is an autocrine amplifier of the function of ILC2s, which act as tissue repair agents after the pulmonary inflammation phase in N. braziliensis infections. ILC2s have IL-9 receptors (IL-9R) which, when activated by their ligands, are responsible for inducing ILC2s to produce IL-5 and IL-13. In IL9r(−/−)mice, the production of these cytokines was strongly compromised from the sixth to the twelfth day post infection, while there were no changes in their production (and in IL-4) by CD4+ T cells. The fact that the production of IL-4 by ILC2 was not compromised shows that IL-9R depletion specifically interferes with the Th9 response [36].
In association with this mechanism, it was described that in infections by Strongyloides stercoralis, the production of IL-9 was directly associated with the infection in a monofunctional way (individually produced) or dual-functional way (co-expressed with IL-10). IL-10, as well as the TGF-β produced by these cells, are also responsible for inducing the production of antigen-specific IL-9. However, this increase did not remain after treatment and the elimination of the parasite, demonstrating that Th9 acts during active infection [38,39]. Rather interestingly, although IL-1 and IL-6 together enabled Th17 differentiation, the combination of IL-1 with TGF-β and IL-4 was able to induce the differentiation of Th9 and Th2 IL9+ in an infection by Brugia malayi [40].
The surprising effect of the induction of Th9 by IL-1 was also reported by other authors as an IL-9 co-stimulator, as it is not able to stimulate the production of IL-9 in an isolated way [41,42,43]. In an infection with T. muris, it was shown that IL-4 requires the co-stimulation of IL-1α or IL-1β to direct the differentiation of helper lymphocytes to the Th9 profile, as characterized by the production of IL-9 and IL-13. Furthermore, without IL-1, the mice were not able to expel the worms, exhibiting high parasite loads 35 days post infection [42].
Concerning TGF-β, it was also shown that alone, neither it nor IL-1 were able to stimulate IL-9 production. As IL-1 is involved in stimulating the Th1 response in Leishmania and Trypanosoma infections and of Th17 in severe schistosomiasis, it is believed that IL-1 acts more as a Th response potentiator than as a polarizer of any specific subpopulation [14,43,44,45]. This highlights the importance of TGF-β/IL-1 and TGF-β/IL-4 in the production of IL-9 [43].
Despite the important role of TGF-β in regulating the expression of Th9, there was no significant reduction in the population of mice expressing T-cell-specific dominant negative TGF-β receptor II [46]. This confirms that TGF-β is not the only cytokine responsible for Th9 differentiation. However, in these individuals, there was an increase in the production of IFN-γ and an exacerbation of the inflammatory process, in addition to a reduced ability to resist infection by Heligmosomoides polygyrus [46].
Rajamanickam et al. (2019) demonstrated that the activity of certain receptors, such as cytotoxic T-lymphocyte-associated antigen 4 (CTLA-4) and programmed death 1 (PD-1), is as important as TGF-β as they are capable of inhibiting the differentiation of the Th1 producers of IFN-γ and IL-2 alone (monofunctional) or combined (dual-functional) and of monofunctional Th17 (producers of IL-17, IL-13 or IL-22) and dual-functional Th17 (producing IFN-γ/IL-17 or IL-17/IL-22). In the context of strongyloidiasis, it has been shown that CTLA-4 increases the expression of IL-4/IL-5-producing Th2 cells, and PD-1 increases the expression of IL-4/IL-5-, IL-4/IL-13- and IL-5/IL-13-producing Th2 cells. Additionally, both factors further increase the IL-9/IL-10-producing Th9 population. Interestingly, these changes did not occur in uninfected individuals, proving that these populations increase only during an active infection [39].
Although the Th2 response is increased after the expression of Th9 cytokines, some studies suggest that Th9 participation is associated with a greater severity of infection and worse prognosis in individuals with helminth infections [40,47,48]. It has been shown that this Th9 subpopulation participates in the formation of lymphedema in patients with lymphatic filariasis, whose response is classically orchestrated by Th1 and Th17 lymphocytes. The parasite antigen was able to modulate the response to increase the expression of IL-9-producing Th2 in infected individuals. However, in those with lymphedema, the Th9 population was higher, demonstrating that Th2 cells are not associated with the pathogenesis of filariasis lymphatic cells, even though they express IL-9 [40].
3.2. Cestodes
Echinococcus spp. are tapeworms that use a predator–prey relationship in their transmission. The two species that cause major public health problems are Echinococcus multilocularis, which causes human alveolar echinococcosis (AE), and Echinococcus granulosus, which causes cystic echinococcosis (CE). Both are cystic diseases that trigger a strong immune response due to the resistance of the encapsulated parasite [49].
AE is a potentially fatal disease that has been extensively studied in its inflammatory context due to the presence of Th17. In addition, the presence of Treg cells indicates that the parasite is capable of inducing a tolerogenic profile to maintain a stable parasite load, promoting the production of the anti-inflammatory cytokines IL-10 and TGF-β as a mechanism of evading the host’s immune response [50]. In patients with AE, increases in mRNA for IL-9 and in the canonical transcription factors of Th9, PU.1 and IRF-4 were identified. Furthermore, the presence of these mRNAs was markedly higher in injured tissues than in peripheral regions and in healthy tissues [51].
The same observation was made in other studies that identified increases in the same mRNAs in patients infected with E. granulosus with CE, with high levels of IL-9, IL-4, IL-10 and TGF-β that decreased after treatment. Despite this, there was a greater contribution of GATA3 than IRF4 to the development of CE, and tge greater circulation of Th9 was positively associated with Th2. This was confirmed by the increased circulation of the aforementioned interleukins in infected patients when compared to the post-treatment period [52,53].
It has also been shown that TGF-β exerts its biological effects by binding to type I and type II membrane receptors that transduce signaling to the nucleus via the phosphorylation of Smads proteins. Smad2 and Smad4 were associated with Th9 differentiation due to their influence on IL-9 production, including in Haemonchus contortus and E. granulosus infections [54,55]. It was shown that TGF-β inhibition down-regulated PU.1, IL-9, IL-10 and Smad3. There was also an increase in IL-9R expression in the cytoplasm of hepatocytes and lymphocyte infiltration in the portal system. Taken together, these data are consistent with increased inflammation and fibrogenesis found in TGF-β/Smad-mediated CE [53].
Additionally, the literature describes that in the differentiation of naïve T cells into Th9 via the IRF4 and Smad3/Smad4 pathways, Th2 cells precede the appearance of Th9 and their cytokines, as if the presence of Th2 were essential for Th0 differentiation into Th9. Abdelaziz et al. (2020) showed that under IL-4 and TGF-β stimulation, there was an increase in Smad3/Smad4, and Th2 cells (defined in this study as IL4+) ceased producing IL4 and began to produce IL9. Furthermore, there was an increase in the expression of IRF4, though there was no change in the expression of GATA3 [56].
This study has some limitations. Initially, it had already been shown that Th9 cells can present a particular transcriptional signature (Th9IL-4+IL-1β) whose differentiation is independent of the TGF-β pathway. This population has intense antitumor cytolytic activity which differentiates it from classic Th9IL-4+TGF-β [57]. Thus, the simple production of IL-4 does not characterize a population as Th2. Furthermore, without a specific stimulus, we cannot affirm that increased levels of IRF4 imply the differentiation of Th2 into Th9.
3.3. Trematodes
Zhan et al. (2017) showed the participation of Th9 in granuloma formation in Schistosoma japonicum infections, with levels of IL-9 in the liver and areas of granulomas being significantly higher than in controls. This is due to increased PU.1 expression, which coincided with Schistosoma eggs reaching the liver and being deposited in the tissue. This study was essential for understanding the participation of Th9 in the immunopathogenesis of S. japonicum infections and highlighted its important inflammatory role [58].
The participation of Th9 in the formation of granulomas is extremely important since this is one of the most effective host responses to infection [58]. The granulomas retain eggs and prevent the circulation of harmful substances to the host, such as soluble egg antigens (SEA). Furthermore, granulomas have the function of promoting the excretion of eggs from the host [59].
Blocking IL-9 reduces the inflammatory process, with a reduction in the deposit of type I and III collagen around the egg and muscle actin in the liver and the consequent formation of granulomas. Zhan et al. (2019) observed that the peak of IL-9 production occurred in the seventh week post infection (shortly after the period of oviposition and granulomatous reaction), long before the peak of IL-4 that occurred only in the twelfth week, demonstrating that the Th2 response begin later than Th9, being consistent with the period in which the chronicity of the infection occurs [60] (Figure 1).
Li et al. (2017) also showed that the peak of CD4+ IL-9+ T cells around the fifth week post infection preceded the formation of the granuloma, which occurred between the fifth and sixth weeks and remained at the same level until the tenth week. This interleukin was able to increase the levels of procollagen-III (PC-III) or factors associated with fibrosis, worsening the inflammatory process and the formation of liver fibrosis [7].
4. Conclusions and Future Directions
The Th9 response is intrinsically dependent on the action of the canonical transcription factors, PU.1 and IRF4, but also on other stimulatory factors, such as CTLA4 and PD-1. Furthermore, the production of IL-9 by Th9 takes place through pathways such as TGF-β/Smad, which induces Th9 differentiation and IL-9 production. Although the difference between the Th2 and Th9 subpopulations has already been proven, the cytokines IL-9 and IL-10 are produced by both by mechanisms that induce a type 2 inflammatory response, which is characterized by the action of ILC2s, mast cells, basophils and other cells that act together for a single objective: the expulsion of worms and the consequent reduction in the parasitic load.
Due to the fact that many studies have characterized IL-9 as a cytokine that is produced early in helminthic infections and is responsible for stimulating the production of IL-5 and IL-13, characteristics of the Th2 response, we can speculate that the Th9 response represents the thin line that interconnects the acute and chronic phases of infections, leading the body from an inflammatory response that is unable to contain the spread of the infection to a tolerogenic response that promotes the expulsion of worms and a reduction in the parasitic load, reducing tissue damage and providing protection to hosts. Studies of the dynamics of infections and the characterization of the subpopulations present in different phases and with different parasites could elucidate these questions.
Author Contributions
Conceptualization, Y.L.D.C.O., Y.L.M.d.O., L.L.B. and S.S.D.; investigation, Y.L.D.C.O. and Y.L.M.d.O.; writing—original draft preparation, Y.L.D.C.O., Y.L.M.d.O. and T.M.C.; writing—review and editing, R.T.F., L.L.B. and S.S.D.; supervision, S.S.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Licona-Limón, P.; Arias-Rojas, A.; Olguín-Martínez, E. IL-9 and Th9 in parasite immunity. Semin. Immunopathol. 2017, 39, 29–38. [Google Scholar] [CrossRef]
- World Health Organization. Available online: https://www.who.int/health-topics/schistosomiasis#tab=tab_1 (accessed on 25 April 2023).
- Pan American Health Organization (PAHO). Available online: https://www.paho.org/en/topics/soil-transmitted-helminthiasis (accessed on 25 April 2023).
- Haase, P.; Voehringer, D. Regulation of the humoral type 2 immune response against allergens and helminths. Eur. J. Immunol. 2021, 51, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Faulkner, H.; Renauld, J.C.; Van Snick, J.; Grencis, R.K. Interleukin-9 enhances resistance to the intestinal nematode Trichuris muris. Infect. Immun. 1998, 66, 3832–3840. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.F.; Lind, E.F.; Gondek, D.C.; Bennett, K.A.; Gleeson, M.W.; Pino-Lagos, K.; Scott, Z.A.; Coyle, A.J.; Reed, J.L.; Van Snick, J.; et al. Mast cells are essential intermediaries in regulatory T-cell tolerance. Nature 2006, 442, 997–1002. [Google Scholar] [CrossRef]
- Li, L.; Xie, H.; Wang, M.; Qu, J.; Cha, H.; Yang, Q.; Feng, Y.; Qi, Y.; Qiu, H.; Dong, N.; et al. Characteristics of IL-9 induced by Schistosoma japonicum infection in C57BL/6 mouse liver. Sci. Rep. 2017, 7, 2343. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Gery, I. The unique features of Th9 cells and their products. Crit. Rev. Immunol. 2012, 32, 1–10. [Google Scholar] [CrossRef]
- Veldhoen, M.; Uyttenhove, C.; van Snick, J.; Helmby, H.; Westendorf, A.; Buer, J.; Martin, B.; Wilhelm, C.; Stockinger, B. Transforming growth factor-beta ‘reprograms’ the differentiation of T helper 2 cells and promotes an interleukin 9-producing subset. Nat. Immunol. 2008, 9, 1341–1346. [Google Scholar] [CrossRef]
- Bilate, A.M.B. Inflamação, citosinas, proteínas da fase aguda e implicações terapêuticas. Reumatol. Clín. 2007, 8, 47–51. [Google Scholar]
- Abbas, A.K.; Lichtman, A.H.; Pillai, S. Imunologia Celular e Molecular, 6th ed.; Editora Elsevier: Rio de Janeiro, Brasil, 2008. [Google Scholar]
- Balbino, C.A.; Pereira, L.M.; Curi, R. Mecanismos envolvidos na cicatrização: Uma revisão. Rev. Bras. Farm. 2005, 41, 27–51. [Google Scholar] [CrossRef]
- Chamusca, F.V.; Reis, S.R.A.; Lemaire, D.; Medrado, A.P. Mediadores do efeito sistêmico do processo inflamatório e terapias fotobiomoduladoras: Uma revisão de literatura. Rev. Ciên. Méd. Biol. 2012, 11, 70–78. [Google Scholar] [CrossRef]
- Zheng, B.; Zhang, J.; Chen, H.; Nie, H.; Miller, H.; Gong, Q.; Liu, C. T Lymphocyte-Mediated Liver Immunopathology of Schistosomiasis. Front. Immunol. 2020, 11, 61. [Google Scholar] [CrossRef]
- Dardalhon, V.; Awasthi, A.; Kwon, H.; Galileos, G.; Gao, W.; Sobel, R.A.; Mitsdoerffer, M.; Strom, T.B.; Elyaman, W.; Ho, I.C.; et al. IL-4 inhibits TGF-beta-induced Foxp3+ T cells and, together with TGF-beta, generates IL-9+ IL-10+ Foxp3(-) effector T cells. Nat. Immunol. 2008, 9, 1347–1355. [Google Scholar] [CrossRef]
- Chang, H.C.; Sehra, S.; Goswami, R.; Yao, W.; Yu, Q.; Stritesky, G.L.; Jabeen, R.; McKinley, C.; Ahyi, A.N.; Han, L.; et al. The transcription factor PU.1 is required for the development of IL-9-producing T cells and allergic inflammation. Nat. Immunol. 2010, 11, 527–534. [Google Scholar] [CrossRef]
- Staudt, V.; Bothur, E.; Klein, M.; Lingnau, K.; Reuter, S.; Grebe, N.; Gerlitzki, B.; Hoffmann, M.; Ulges, A.; Taube, C.; et al. Interferon-regulatory factor 4 is essential for the developmental program of T helper 9 cells. Immunity 2010, 33, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Niedbala, W.; Besnard, A.G.; Nascimento, D.C.; Donate, P.B.; Sonego, F.; Yip, E.; Guabiraba, R.; Chang, H.D.; Fukada, S.Y.; Salmond, R.J.; et al. Nitric oxide enhances Th9 cell differentiation and airway inflammation. Nat. Commun. 2014, 5, 4575. [Google Scholar] [CrossRef]
- Goswami, R.; Jabeen, R.; Yagi, R.; Pham, D.; Zhu, J.; Goenka, S.; Kaplan, M.H. STAT6-dependent regulation of Th9 development. J. Immunol. 2012, 188, 968–975. [Google Scholar] [CrossRef] [PubMed]
- Wiener, Z.; Falus, A.; Toth, S. IL-9 increases the expression of several cytokines in activated mast cells, while the IL-9-induced IL-9 production is inhibited in mast cells of histamine-free transgenic mice. Cytokine 2004, 26, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.T.; Ye, J.J.; Alonso, M.N.; Landrigan, A.; Cheung, R.K.; Engleman, E.; Utz, P.J. Regulation of human Th9 differentiation by type I interferons and IL-21. Immunol. Cell. Biol. 2010, 88, 624–631. [Google Scholar] [CrossRef]
- Meylan, F.; Siegel, R.M. TNF superfamily cytokines in the promotion of Th9 differentiation and immunopathology. Semin. Immunopathol. 2017, 39, 21–28. [Google Scholar] [CrossRef]
- Liu, J.; Harberts, E.; Tammaro, A.; Girardi, N.; Filler, R.B.; Fishelevich, R.; Temann, A.; Licona-Limón, P.; Girardi, M.; Flavell, R.A.; et al. IL-9 regulates allergen-specific Th1 responses in allergic contact dermatitis. J. Investig. Dermatol. 2014, 134, 1903–1911. [Google Scholar] [CrossRef]
- Gu, Z.W.; Wang, Y.X.; Cao, Z.W. Neutralization of interleukin-9 ameliorates symptoms of allergic rhinitis by reducing Th2, Th9, and Th17 responses and increasing the Treg response in a murine model. Oncotarget 2017, 28, 14314–14324. [Google Scholar] [CrossRef]
- Faulkner, H.; Humphreys, N.; Renauld, J.C.; Van Snick, J.; Grencis, R. Interleukin-9 is involved in host protective immunity to intestinal nematode infection. Eur. J. Immunol. 1997, 27, 2536–2540. [Google Scholar] [CrossRef] [PubMed]
- Temann, U.A.; Geba, G.P.; Rankin, J.A.; Flavell, R.A. Expression of interleukin 9 in the lungs of transgenic mice causes airway inflammation, mast cell hyperplasia, and bronchial hyperresponsiveness. J. Exp. Med. 1998, 188, 1307–1320. [Google Scholar] [CrossRef] [PubMed]
- Temann, U.A.; Ray, P.; Flavell, R.A. Pulmonary overexpression of IL-9 induces Th2 cytokine expression, leading to immune pathology. J. Clin. Investig. 2002, 109, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Else, K.J.; Finkelman, F.D.; Maliszewski, C.R.; Grencis, R.K. Cytokine-mediated regulation of chronic intestinal helminth infection. J. Exp. Med. 1994, 179, 347–351. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Samblas, M.; García-Rodríguez, J.J.; Trelis, M.; Bernal, D.; Lopez-Jaramillo, F.J.; Santoyo-Gonzalez, F.; Vilchez, S.; Espino, A.M.; Bolás-Fernández, F.; Osuna, A. Self-adjuvanting C18 lipid vinil sulfone-PP2A vaccine: Study of the induced immunomodulation against. Open Biol. 2017, 7, 170031. [Google Scholar] [CrossRef]
- Pennock, J.L.; Grencis, R.K. The mast cell and gut nematodes: Damage and defence. Chem. Immunol. Allergy 2006, 90, 128–140. [Google Scholar] [CrossRef]
- Nagashima, H.; Mahlakõiv, T.; Shih, H.Y.; Davis, F.P.; Meylan, F.; Huang, Y.; Harrison, O.J.; Yao, C.; Mikami, Y.; Urban, J.F.; et al. Neuropeptide CGRP Limits Group 2 Innate Lymphoid Cell Responses and Constrains Type 2 Inflammation. Immunity 2019, 51, 682–695.e686. [Google Scholar] [CrossRef]
- Inclan-Rico, J.M.; Hernandez, C.M.; Henry, E.K.; Federman, H.G.; Sy, C.B.; Ponessa, J.J.; Lemenze, A.D.; Joseph, N.; Soteropoulos, P.; Beaulieu, A.M.; et al. Trichinella spiralis-induced mastocytosis and erythropoiesis are simultaneously supported by a bipotent mast cell/erythrocyte precursor cell. PLoS Pathog. 2020, 16, e1008579. [Google Scholar] [CrossRef]
- Licona-Limón, P.; Henao-Mejia, J.; Temann, A.U.; Gagliani, N.; Licona-Limón, I.; Ishigame, H.; Hao, L.; Herbert, D.R.; Flavell, R.A. Th9 Cells Drive Host Immunity against Gastrointestinal Worm Infection. Immunity 2013, 39, 744–757. [Google Scholar] [CrossRef]
- Henry, E.K.; Sy, C.B.; Inclan-Rico, J.M.; Espinosa, V.; Ghanny, S.S.; Dwyer, D.F.; Soteropoulos, P.; Rivera, A.; Siracusa, M.C. Carbonic anhydrase enzymes regulate mast cell-mediated inflammation. J. Exp. Med. 2016, 213, 1663–1673. [Google Scholar] [CrossRef] [PubMed]
- Angkasekwinai, P.; Sodthawon, W.; Jeerawattanawart, S.; Hansakon, A.; Pattanapanyasat, K.; Wang, Y.H. ILC2s activated by IL-25 promote antigen-specific Th2 and Th9 functions that contribute to the control of Trichinella spiralis infection. PLoS ONE 2017, 12, e0184684. [Google Scholar] [CrossRef]
- Turner, J.E.; Morrison, P.J.; Wilhelm, C.; Wilson, M.; Ahlfors, H.; Renauld, J.C.; Panzer, U.; Helmby, H.; Stockinger, B. IL-9-mediated survival of type 2 innate lymphoid cells promotes damage control in helminth-induced lung inflammation. J. Exp. Med. 2013, 210, 2951–2965. [Google Scholar] [CrossRef] [PubMed]
- Angkasekwinai, P.; Srimanote, P.; Wang, Y.H.; Pootong, A.; Sakolvaree, Y.; Pattanapanyasat, K.; Chaicumpa, W.; Chaiyaroj, S.; Dong, C. Interleukin-25 (IL-25) promotes efficient protective immunity against Trichinella spiralis infection by enhancing the antigen-specific IL-9 response. Infect. Immun. 2013, 81, 3731–3741. [Google Scholar] [CrossRef] [PubMed]
- Anuradha, R.; Munisankar, S.; Bhootra, Y.; Jagannathan, J.; Dolla, C.; Kumaran, P.; Nutman, T.B.; Babu, S. IL-10- and TGFβ-mediated Th9 Responses in a Human Helminth Infection. PLoS Negl. Trop. Dis. 2016, 10, e0004317. [Google Scholar] [CrossRef]
- Rajamanickam, A.; Munisankar, S.; Dolla, C.; Nutman, T.B.; Babu, S. Cytotoxic T-Lymphocyte-Associated Antigen 4 (CTLA-4)- and Programmed Death 1 (PD-1)-Mediated Regulation of Monofunctional and Dual Functional CD4. Infect. Immun. 2019, 87, e00469-19. [Google Scholar] [CrossRef]
- Anuradha, R.; George, P.J.; Hanna, L.E.; Chandrasekaran, V.; Kumaran, P.; Nutman, T.B.; Babu, S. IL-4-, TGF-β-, and IL-1-dependent expansion of parasite antigen-specific Th9 cells is associated with clinical pathology in human lymphatic filariasis. J. Immunol. 2013, 191, 2466–2473. [Google Scholar] [CrossRef]
- Schmitt, E.; Beuscher, H.U.; Huels, C.; Monteyne, P.; van Brandwijk, R.; van Snick, J.; Ruede, E. IL-1 serves as a secondary signal for IL-9 expression. J. Immunol. 1991, 147, 3848–3854. [Google Scholar] [CrossRef]
- Helmby, H.; Grencis, R.K. Interleukin 1 plays a major role in the development of Th2-mediated immunity. Eur. J. Immunol. 2004, 34, 3674–3681. [Google Scholar] [CrossRef]
- Uyttenhove, C.; Brombacher, F.; Van Snick, J. TGF-β interactions with IL-1 family members trigger IL-4-independent IL-9 production by mouse CD4(+) T cells. Eur. J. Immunol. 2010, 40, 2230–2235. [Google Scholar] [CrossRef]
- Poveda, C.; Fresno, M.; Gironès, N.; Martins-Filho, O.A.; Ramírez, J.D.; Santi-Rocca, J.; Marin-Neto, J.A.; Morillo, C.A.; Rosas, F.; Guhl, F. Cytokine profiling in Chagas disease: Towards understanding the association with infecting Trypanosoma cruzi discrete typing units (a BENEFIT TRIAL sub-study). PLoS ONE 2014, 9, e91154. [Google Scholar] [CrossRef] [PubMed]
- Amorim, E.A.D.S.; de França, Á.; Pereira, V.R.A.; Brelaz-de-Castro, M.C.A. IL-1 family and Cutaneous Leishmaniasis: A poorly understood relationship. Cytokine Growth Factor Rev. 2021, 57, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, L.A.; Maizels, R.M. Cutting edge: In the absence of TGF-β signaling in T cells, fewer CD103+ regulatory T cells develop, but exuberant IFN-γ production renders mice more susceptible to helminth infection. J. Immunol. 2012, 189, 1113–1117. [Google Scholar] [CrossRef] [PubMed]
- Anuradha, R.; Munisankar, S.; Bhootra, Y.; Dolla, C.; Kumaran, P.; Nutman, T.B.; Babu, S. Modulation of CD4+ and CD8+ T cell function and cytokine responses in Strongyloides stercoralis infection by interleukin-27 (IL-27) and IL-37. Infect. Immun. 2017, 85, e00500-17. [Google Scholar] [CrossRef]
- Sibi, J.M.; Mohan, V.; Munisankar, S.; Babu, S.; Aravindhan, V. Augmented Innate and Adaptive Immune Responses Under Conditions of Diabetes-Filariasis Comorbidity. Front. Immunol. 2021, 12, 716515. [Google Scholar] [CrossRef]
- Woolsey, I.D.; Miller, A.L. Echinococcus granulosus sensu lato and Echinococcus multilocularis: A review. Res. Vet. Sci. 2021, 135, 517–522. [Google Scholar] [CrossRef]
- Tuxun, T.; Wang, J.H.; Lin, R.Y.; Shan, J.Y.; Tai, Q.W.; Li, T.; Zhang, J.H.; Zhao, J.M.; Wen, H. Th17/Treg imbalance in patients with liver cystic echinococcosis. Parasite Immunol. 2012, 34, 520–527. [Google Scholar] [CrossRef]
- Tuxun, T.; Apaer, S.; Ma, H.Z.; Zhang, H.; Aierken, A.; Lin, R.Y.; Wen, H. The Potential Role of Th9 Cell Related Cytokine and Transcription Factors in Patients with Hepatic Alveolar Echinococcosis. J. Immunol. Res. 2015, 2015, 895416. [Google Scholar] [CrossRef]
- Pang, N.; Zhang, F.; Li, S.; Zhu, Y.; Zhang, C.; An, M.; Wang, H.; Mamuti, W.; Ding, J.; Fan, H. TGF-β/Smad signaling pathway positively up-regulates the differentiation of Interleukin-9-producing CD4. J. Infect. 2018, 76, 406–416. [Google Scholar] [CrossRef]
- Pang, N.; Zhang, F.; Ma, X.; Zhang, Z.; Zhao, H.; Xin, Y.; Wang, S.; Zhu, Y.; Wen, H.; Ding, J. Th9/IL-9 profile in human echinococcosis: Their involvement in immune response during infection by Echinococcus granulosus. Mediat. Inflamm. 2014, 2014, 781649. [Google Scholar] [CrossRef]
- Wang, A.; Pan, D.; Lee, Y.H.; Martinez, G.J.; Feng, X.H.; Dong, C. Cutting edge: Smad2 and Smad4 regulate TGF-β-mediated Il9 gene expression via EZH2 displacement. J. Immunol. 2013, 191, 4908–4912. [Google Scholar] [CrossRef] [PubMed]
- Memon, M.A.; Naqvi, M.A.; Xin, H.; Meng, L.; Hasan, M.W.; Haseeb, M.; Lakho, S.A.; Aimulajiang, K.; Bu, Y.; Xu, L.; et al. Immunomodulatory dynamics of excretory and secretory products on Th9 immune response during Haemonchus contortus infection in goat. PLoS Negl. Trop. Dis. 2020, 14, e0008218. [Google Scholar] [CrossRef] [PubMed]
- Abdelaziz, M.H.; Wang, H.; Cheng, J.; Xu, H. Th2 cells as an intermediate for the differentiation of naïve T cells into Th9 cells, associated with the Smad3/Smad4 and IRF4 pathway. Exp. Ther. Med. 2020, 19, 1947–1954. [Google Scholar] [CrossRef] [PubMed]
- Xue, G.; Jin, G.; Fang, J.; Lu, Y. IL-4 together with IL-1β induces antitumor Th9 cell differentiation in the absence of TGF-β signaling. Nat. Commun. 2019, 10, 1376. [Google Scholar] [CrossRef]
- Zhan, T.; Zhang, T.; Wang, Y.; Wang, X.; Lin, C.; Ma, H.; Duan, Z.; Li, C.; Xu, J.; Xia, C. Dynamics of Th9 cells and their potential role in immunopathogenesis of murine schistosomiasis. Parasit. Vectors 2017, 10, 305. [Google Scholar] [CrossRef]
- Costain, A.H.; MacDonald, A.S.; Smits, H.H. Schistosome Egg Migration: Mechanisms, Pathogenesis and Host Immune Responses. Front. Immunol. 2018, 20, 3042. [Google Scholar] [CrossRef] [PubMed]
- Zhan, T.; Ma, H.; Jiang, S.; Zhong, Z.; Wang, X.; Li, C.; Yu, D.; Liu, L.; Xu, J.; Xia, C. Interleukin-9 blockage reduces early hepatic granuloma formation and fibrosis during Schistosoma japonicum infection in mice. Immunology 2019, 158, 296–303. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).