
MicroRNA-511-3p Mediated Modulation of the Peroxisome Proliferator-Activated Receptor Gamma (PPARγ) Controls LPS-Induced Inflammatory Responses in Human Monocyte Derived DCs

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Generation of Monocyte-Derived Dendritic Cells (DCs)
2.2. Flow Cytometry Analysis
2.3. Quantification of IDO Activity
2.4. Live/Dead Assay
2.5. RNA Interference
2.6. RNA Isolation and cDNA Synthesis
2.7. Real Time PCR (qRT-PCR)
2.8. Cytokine ELISA
2.9. Statistical Analysis
3. Results
3.1. Changes in miR-511-3p Expression Affect PPARγ Expression and Activity in Human DCs
3.2. PPARγ Activation Modulates Cytokine Production in miR-511-3p-Transfected DCs
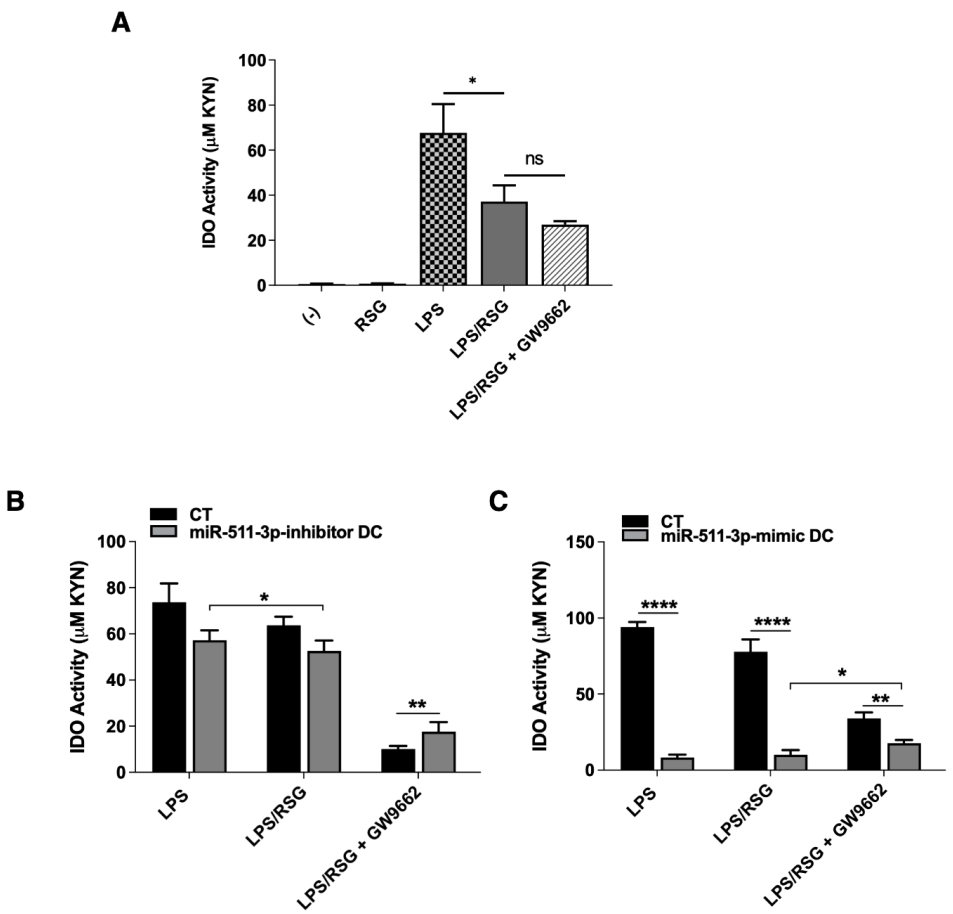
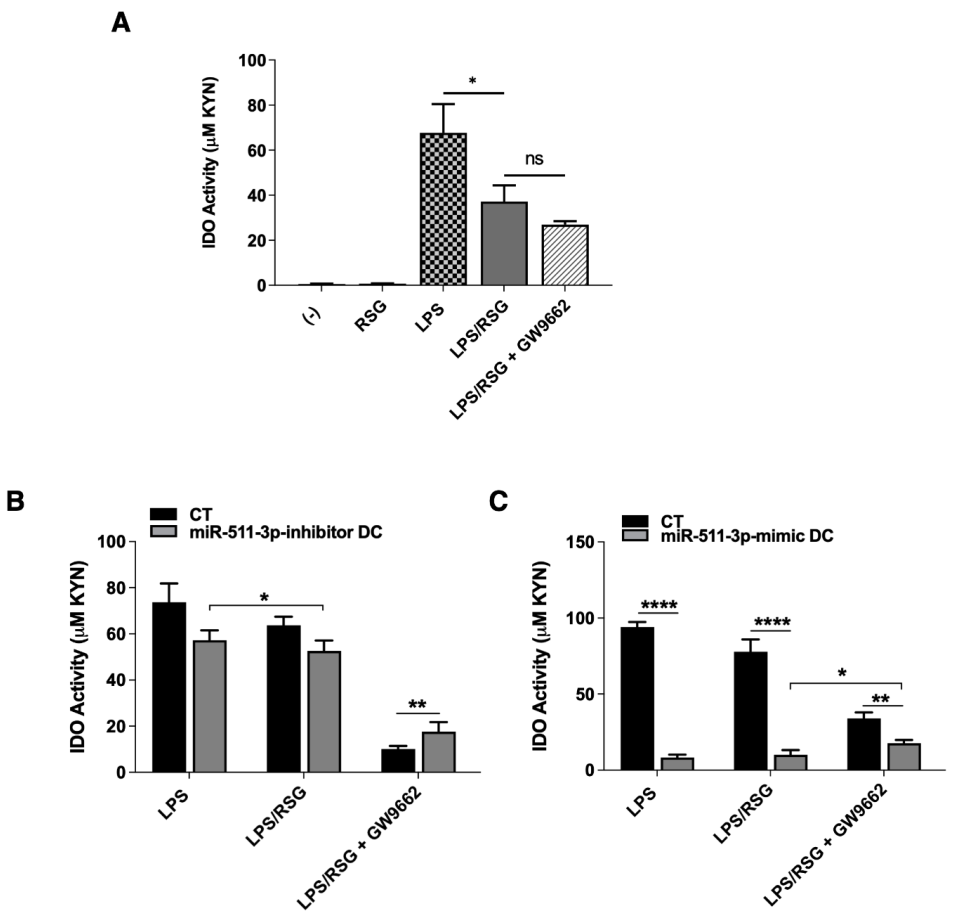
3.3. PPARγ Modulation of LPS-Induced IDO Activity Is Influenced by miR-511-3p Expression in Human DCs
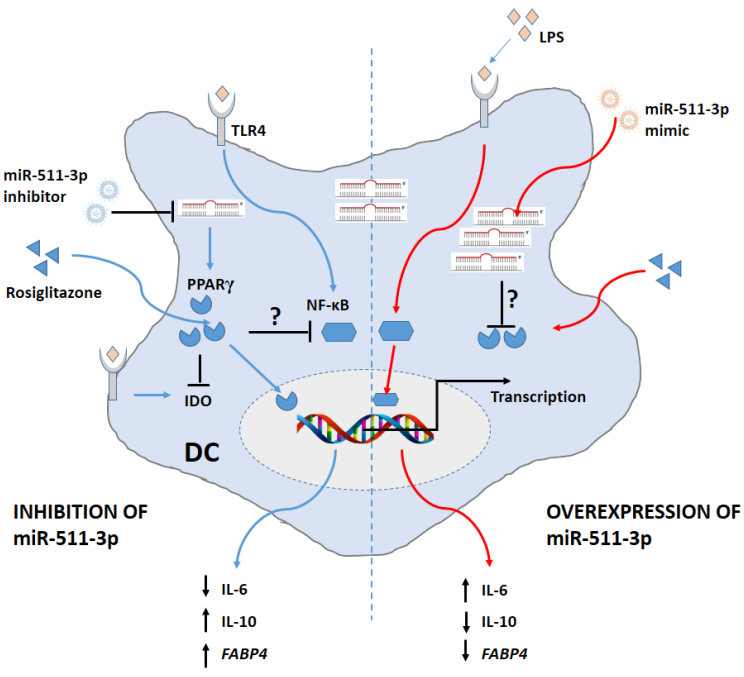
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rosenfeld, Y.; Shai, Y. Lipopolysaccharide (Endotoxin)-host defense antibacterial peptides interactions: Role in bacterial resistance and prevention of sepsis. Biochim. Biophys. Acta (BBA) Biomembr. 2006, 1758, 1513–1522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilroy, D.; Lawrence, T. The resolution of acute inflammation: A ‘tipping point’ in the development of chronic inflammatory diseases. In The Resolution of Inflammation; Rossi, A.G., Sawatzky, D.A., Eds.; Birkhäuser Basel: Basel, Switzerland, 2008; pp. 1–18. [Google Scholar]
- Lawrence, T.; Fong, C. The resolution of inflammation: Anti-inflammatory roles for NF-κB. Int. J. Biochem. Cell Biol. 2010, 42, 519–523. [Google Scholar] [CrossRef]
- Lawrence, T. The Nuclear Factor NF-κB Pathway in Inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef] [Green Version]
- Ahmadian, M.; Suh, J.M.; Hah, N.; Liddle, C.; Atkins, A.R.; Downes, M.; Evans, R.M. PPARgamma signaling and metabolism: The good, the bad and the future. Nat. Med. 2013, 19, 557–566. [Google Scholar] [CrossRef] [Green Version]
- Klotz, L.; Dani, I.; Edenhofer, F.; Nolden, L.; Evert, B.; Paul, B.; Kolanus, W.; Klockgether, T.; Knolle, P.; Diehl, L. Peroxisome proliferator-activated receptor gamma control of dendritic cell function contributes to development of CD4+ T cell anergy. J. Immunol. 2007, 178, 2122–2131. [Google Scholar] [CrossRef] [Green Version]
- Nencioni, A.; Grunebach, F.; Zobywlaski, A.; Denzlinger, C.; Brugger, W.; Brossart, P. Dendritic cell immunogenicity is regulated by peroxisome proliferator-activated receptor gamma. J. Immunol. 2002, 169, 1228–1235. [Google Scholar] [CrossRef] [PubMed]
- Angeli, V.; Hammad, H.; Staels, B.; Capron, M.; Lambrecht, B.N.; Trottein, F. Peroxisome proliferator-activated receptor gamma inhibits the migration of dendritic cells: Consequences for the immune response. J. Immunol. 2003, 170, 5295–5301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faveeuw, C.; Fougeray, S.; Angeli, V.; Fontaine, J.; Chinetti, G.; Gosset, P.; Delerive, P.; Maliszewski, C.; Capron, M.; Staels, B.; et al. Peroxisome proliferator-activated receptor gamma activators inhibit interleukin-12 production in murine dendritic cells. FEBS Lett. 2000, 486, 261–266. [Google Scholar] [CrossRef]
- Salazar, F.; Awuah, D.; Negm, O.H.; Shakib, F.; Ghaemmaghami, A.M. The role of indoleamine 2,3-dioxygenase-aryl hydrocarbon receptor pathway in the TLR4-induced tolerogenic phenotype in human DCs. Sci. Rep. 2017, 7, 43337. [Google Scholar] [CrossRef] [PubMed]
- Fallarino, F.; Grohmann, U.; Vacca, C.; Bianchi, R.; Orabona, C.; Spreca, A.; Fioretti, M.C.; Puccetti, P. T cell apoptosis by tryptophan catabolism. Cell Death Differ. 2002, 9, 1069–1077. [Google Scholar] [CrossRef] [PubMed]
- Tas, S.W.; Vervoordeldonk, M.J.; Hajji, N.; Schuitemaker, J.H.N.; van der Sluijs, K.F.; May, M.J.; Ghosh, S.; Kapsenberg, M.L.; Tak, P.P.; de Jong, E.C. Noncanonical NF-κB signaling in dendritic cells is required for indoleamine 2,3-dioxygenase (IDO) induction and immune regulation. Blood 2007, 110, 1540–1549. [Google Scholar] [CrossRef] [Green Version]
- Mbongue, J.C.; Nicholas, D.A.; Torrez, T.W.; Kim, N.-S.; Firek, A.F.; Langridge, W.H.R. The Role of Indoleamine 2, 3-Dioxygenase in Immune Suppression and Autoimmunity. Vaccines 2015, 3, 703–729. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; Xiao, C.; Evans, K.S.; Theivanthiran, T.; DeVito, N.; Holtzhausen, A.; Liu, J.; Liu, X.; Boczkowski, D.; Nair, S.; et al. Paracrine Wnt5a-β-Catenin Signaling Triggers a Metabolic Program that Drives Dendritic Cell Tolerization. Immunity 2018, 48, 147–160.e147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bentwich, I.; Avniel, A.; Karov, Y.; Aharonov, R.; Gilad, S.; Barad, O.; Barzilai, A.; Einat, P.; Einav, U.; Meiri, E.; et al. Identification of hundreds of conserved and nonconserved human microRNAs. Nat. Genet. 2005, 37, 766–770. [Google Scholar] [CrossRef] [PubMed]
- Awuah, D.; Alobaid, M.; Latif, A.; Salazar, F.; Emes, R.D.; Ghaemmaghami, A.M. The Cross-Talk between miR-511-3p and C-Type Lectin Receptors on Dendritic Cells Affects Dendritic Cell Function. J. Immunol. 2019, 203, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Tserel, L.; Runnel, T.; Kisand, K.; Pihlap, M.; Bakhoff, L.; Kolde, R.; Peterson, H.; Vilo, J.; Peterson, P.; Rebane, A. MicroRNA expression profiles of human blood monocyte-derived dendritic cells and macrophages reveal miR-511 as putative positive regulator of Toll-like receptor 4. J. Biol. Chem. 2011, 286, 26487–26495. [Google Scholar] [CrossRef] [Green Version]
- Karo-Atar, D.; Itan, M.; Pasmanik-Chor, M.; Munitz, A. MicroRNA profiling reveals opposing expression patterns for miR-511 in alternatively and classically activated macrophages. J. Asthma Off. J. Assoc. Care Asthma 2015, 52, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Fleischmann, K.K.; Pagel, P.; von Frowein, J.; Magg, T.; Roscher, A.A.; Schmid, I. The leukemogenic fusion gene MLL-AF9 alters microRNA expression pattern and inhibits monoblastic differentiation via miR-511 repression. J. Exp. Clin. Cancer Res. 2016, 35, 9. [Google Scholar] [CrossRef] [Green Version]
- Salazar, F.; Hall, L.; Negm, O.H.; Awuah, D.; Tighe, P.J.; Shakib, F.; Ghaemmaghami, A.M. The mannose receptor negatively modulates the Toll-like receptor 4-aryl hydrocarbon receptor-indoleamine 2,3-dioxygenase axis in dendritic cells affecting T helper cell polarization. J. Allergy Clin. Immunol. 2016, 137, 1841–1851 e1842. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Nieto, S.; Johal, R.K.; Shakesheff, K.M.; Emara, M.; Royer, P.J.; Chau, D.Y.; Shakib, F.; Ghaemmaghami, A.M. Laminin and fibronectin treatment leads to generation of dendritic cells with superior endocytic capacity. PLoS ONE 2010, 5, e10123. [Google Scholar] [CrossRef] [Green Version]
- Aldajani, W.A.; Salazar, F.; Sewell, H.F.; Knox, A.; Ghaemmaghami, A.M. Expression and regulation of immune-modulatory enzyme indoleamine 2,3-dioxygenase (IDO) by human airway epithelial cells and its effect on T cell activation. Oncotarget 2016, 7, 57606–57617. [Google Scholar] [CrossRef] [Green Version]
- Donaldson, A.R.; Tanase, C.E.; Awuah, D.; Vasanthi Bathrinarayanan, P.; Hall, L.; Nikkhah, M.; Khademhosseini, A.; Rose, F.; Alexander, C.; Ghaemmaghami, A.M. Photocrosslinkable Gelatin Hydrogels Modulate the Production of the Major Pro-inflammatory Cytokine, TNF-α, by Human Mononuclear Cells. Front. Bioeng. Biotechnol. 2018, 6. [Google Scholar] [CrossRef] [PubMed]
- Ghaemmaghami, A.M.; Shakib, F. Human T cells that have been conditioned by the proteolytic activity of the major dust mite allergen Der p 1 trigger enhanced immunoglobulin E synthesis by B cells. Clin. Exp. Allergy 2002, 32, 728–732. [Google Scholar] [CrossRef]
- Ricote, M.; Li, A.C.; Willson, T.M.; Kelly, C.J.; Glass, C.K. The peroxisome proliferator-activated receptor-gamma is a negative regulator of macrophage activation. Nature 1998, 391, 79–82. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Ting, A.T.; Seed, B. PPAR-gamma agonists inhibit production of monocyte inflammatory cytokines. Nature 1998, 391, 82–86. [Google Scholar] [CrossRef]
- Szatmari, I.; Gogolak, P.; Im, J.S.; Dezso, B.; Rajnavolgyi, E.; Nagy, L. Activation of PPARγ Specifies a Dendritic Cell Subtype Capable of Enhanced Induction of iNKT Cell Expansion. Immunity 2004, 21, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Shi, L.; Xin, W.; Xu, J.; Xu, J.; Li, Q.; Xu, Z.; Wang, J.; Wang, G.; Yao, W.; et al. Activation of PPARγ inhibits pro-inflammatory cytokines production by upregulation of miR-124 in vitro and in vivo. Biochem. Biophys. Res. Commun. 2017, 486, 726–731. [Google Scholar] [CrossRef]
- Appel, S.; Mirakaj, V.; Bringmann, A.; Weck, M.M.; Grünebach, F.; Brossart, P. PPAR-γ agonists inhibit toll-like receptor-mediated activation of dendritic cells via the MAP kinase and NF-κB pathways. Blood 2005, 106, 3888–3894. [Google Scholar] [CrossRef]
- Asada, K.; Sasaki, S.; Suda, T.; Chida, K.; Nakamura, H. Antiinflammatory Roles of Peroxisome Proliferator–activated Receptor γ in Human Alveolar Macrophages. Am. J. Respir. Crit. Care Med. 2004, 169, 195–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, C.M.; Hale, P.T.; Carlin, J.M. The Role of IFN-γ and TNF-α-Responsive Regulatory Elements in the Synergistic Induction of Indoleamine Dioxygenase. J. Interferon Cytokine Res. Off. J. Int. Soc. Interferon Cytokine Res. 2005, 25, 20–30. [Google Scholar] [CrossRef]
- Robinson, C.M.; Shirey, K.A.; Carlin, J.M. Synergistic transcriptional activation of indoleamine dioxygenase by IFN-gamma and tumor necrosis factor-alpha. J. Interferon Cytokine Res 2003, 23, 413–421. [Google Scholar] [CrossRef]
- Fujigaki, S.; Saito, K.; Sekikawa, K.; Tone, S.; Takikawa, O.; Fujii, H.; Wada, H.; Noma, A.; Seishima, M. Lipopolysaccharide induction of indoleamine 2,3-dioxygenase is mediated dominantly by an IFN-gamma-independent mechanism. Eur. J. Immunol. 2001, 31, 2313–2318. [Google Scholar] [CrossRef]
- Varga, T.; Nagy, L. Nuclear receptors, transcription factors linking lipid metabolism and immunity: The case of peroxisome proliferator-activated receptor gamma. Eur. J. Clin. Investig. 2008, 38, 695–707. [Google Scholar] [CrossRef]
- Díaz-Delfín, J.; Morales, M.; Caelles, C. Hypoglycemic Action of Thiazolidinediones/Peroxisome Proliferator–Activated Receptor γ by Inhibition of the c-Jun NH2-Terminal Kinase Pathway. Diabetes 2007, 56, 1865–1871. [Google Scholar] [CrossRef] [PubMed]
- Landreth, G. Therapeutic use of agonists of the nuclear receptor PPARgamma in Alzheimer’s disease. Curr. Alzheimer Res. 2007, 4, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.D.; Lichtenstein, G.R.; Deren, J.J.; Sands, B.E.; Hanauer, S.B.; Katz, J.A.; Lashner, B.; Present, D.H.; Chuai, S.; Ellenberg, J.H.; et al. Rosiglitazone for active ulcerative colitis: A randomized placebo-controlled trial. Gastroenterology 2008, 134, 688–695. [Google Scholar] [CrossRef] [Green Version]
- Jennewein, C.; von Knethen, A.; Schmid, T.; Brüne, B. MicroRNA-27b Contributes to Lipopolysaccharide-mediated Peroxisome Proliferator-activated Receptor γ (PPARγ) mRNA Destabilization. J. Biol. Chem. 2010, 285, 11846–11853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samokhvalov, V.; Vriend, J.; Jamieson, K.; Akhnokh, M.; Manne, R.; Falck, J.; Seubert, J. PPARγ signaling is required for mediating eets protective effects in neonatal cardiomyocytes exposed to LPS. Front. Pharmacol. 2014, 5. [Google Scholar] [CrossRef]
- Zhou, M.; Wu, R.; Dong, W.; Jacob, A.; Wang, P. Endotoxin downregulates peroxisome proliferator-activated receptor-γ via the increase in TNF-α release. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 294, R84–R92. [Google Scholar] [CrossRef]
- von Bubnoff, D.; Bieber, T. The indoleamine 2,3-dioxygenase (IDO) pathway controls allergy. Allergy 2012, 67, 718–725. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.E.; Astola, N.; Cribbs, A.P.; Goddard, M.E.; Park, I.; Green, P.; Davies, A.H.; Williams, R.O.; Feldmann, M.; Monaco, C. Indoleamine 2,3-dioxygenase-1 is protective in atherosclerosis and its metabolites provide new opportunities for drug development. Proc. Natl. Acad. Sci. USA 2015, 112, 13033–13038. [Google Scholar] [CrossRef] [Green Version]
- Curti, A.; Trabanelli, S.; Salvestrini, V.; Baccarani, M.; Lemoli, R.M. The role of indoleamine 2,3-dioxygenase in the induction of immune tolerance: Focus on hematology. Blood 2009, 113, 2394–2401. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.Y.; Gonzalez-Martin, A.; Miletic, A.V.; Lai, M.; Knight, S.; Sabouri-Ghomi, M.; Head, S.R.; Macauley, M.S.; Rickert, R.C.; Xiao, C. Transfection of microRNA Mimics Should Be Used with Caution. Front. Genet. 2015, 6, 340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chawla, A.; Barak, Y.; Nagy, L.; Liao, D.; Tontonoz, P.; Evans, R.M. PPAR-γ dependent and independent effects on macrophage-gene expression in lipid metabolism and inflammation. Nat. Med. 2001, 7, 48. [Google Scholar] [CrossRef]
- Celinski, K.; Dworzanski, T.; Korolczuk, A.; Piasecki, R.; Slomka, M.; Madro, A.; Fornal, R. Effects of peroxisome proliferator-activated receptors-gamma ligands on dextran sodium sulphate-induced colitis in rats. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2011, 62, 347–356. [Google Scholar]
- Pisanu, A.; Lecca, D.; Mulas, G.; Wardas, J.; Simbula, G.; Spiga, S.; Carta, A.R. Dynamic changes in pro- and anti-inflammatory cytokines in microglia after PPAR-gamma agonist neuroprotective treatment in the MPTPp mouse model of progressive Parkinson’s disease. Neurobiol. Dis. 2014, 71, 280–291. [Google Scholar] [CrossRef]
- Rajaram, M.V.; Brooks, M.N.; Morris, J.D.; Torrelles, J.B.; Azad, A.K.; Schlesinger, L.S. Mycobacterium tuberculosis activates human macrophage peroxisome proliferator-activated receptor gamma linking mannose receptor recognition to regulation of immune responses. J. Immunol. 2010, 185, 929–942. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer2 | Sequence (5′-3′) |
|---|---|---|
| GAPDH | Forward Reverse | GAGTCAACGGATTTGGTCGT GACAAGCTTCCCGTTCTCAG |
| PPARG | Forward Reverse | TGTGAAGCCCATTGAAGACA ACTCAGGGTGGTTCAGCTTC |
| FABP4 | Forward Reverse | GGATGGAAAATCAACCACCA GGAAGTGACGCCTTTCATGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Awuah, D.; Ruisinger, A.; Alobaid, M.; Mbadugha, C.; Ghaemmaghami, A.M. MicroRNA-511-3p Mediated Modulation of the Peroxisome Proliferator-Activated Receptor Gamma (PPARγ) Controls LPS-Induced Inflammatory Responses in Human Monocyte Derived DCs. Immuno 2022, 2, 104-117. https://doi.org/10.3390/immuno2010008
Awuah D, Ruisinger A, Alobaid M, Mbadugha C, Ghaemmaghami AM. MicroRNA-511-3p Mediated Modulation of the Peroxisome Proliferator-Activated Receptor Gamma (PPARγ) Controls LPS-Induced Inflammatory Responses in Human Monocyte Derived DCs. Immuno. 2022; 2(1):104-117. https://doi.org/10.3390/immuno2010008
Chicago/Turabian StyleAwuah, Dennis, Alisa Ruisinger, Meshal Alobaid, Chidimma Mbadugha, and Amir M. Ghaemmaghami. 2022. "MicroRNA-511-3p Mediated Modulation of the Peroxisome Proliferator-Activated Receptor Gamma (PPARγ) Controls LPS-Induced Inflammatory Responses in Human Monocyte Derived DCs" Immuno 2, no. 1: 104-117. https://doi.org/10.3390/immuno2010008
APA StyleAwuah, D., Ruisinger, A., Alobaid, M., Mbadugha, C., & Ghaemmaghami, A. M. (2022). MicroRNA-511-3p Mediated Modulation of the Peroxisome Proliferator-Activated Receptor Gamma (PPARγ) Controls LPS-Induced Inflammatory Responses in Human Monocyte Derived DCs. Immuno, 2(1), 104-117. https://doi.org/10.3390/immuno2010008