
The Effect of Different Irrigation Water Salinity Levels on Nutrients Uptake, Biochemical Content and Growth Response of Blue Panicum, Quinoa and Silage Maize †

Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Physiological and Biochemical Properties
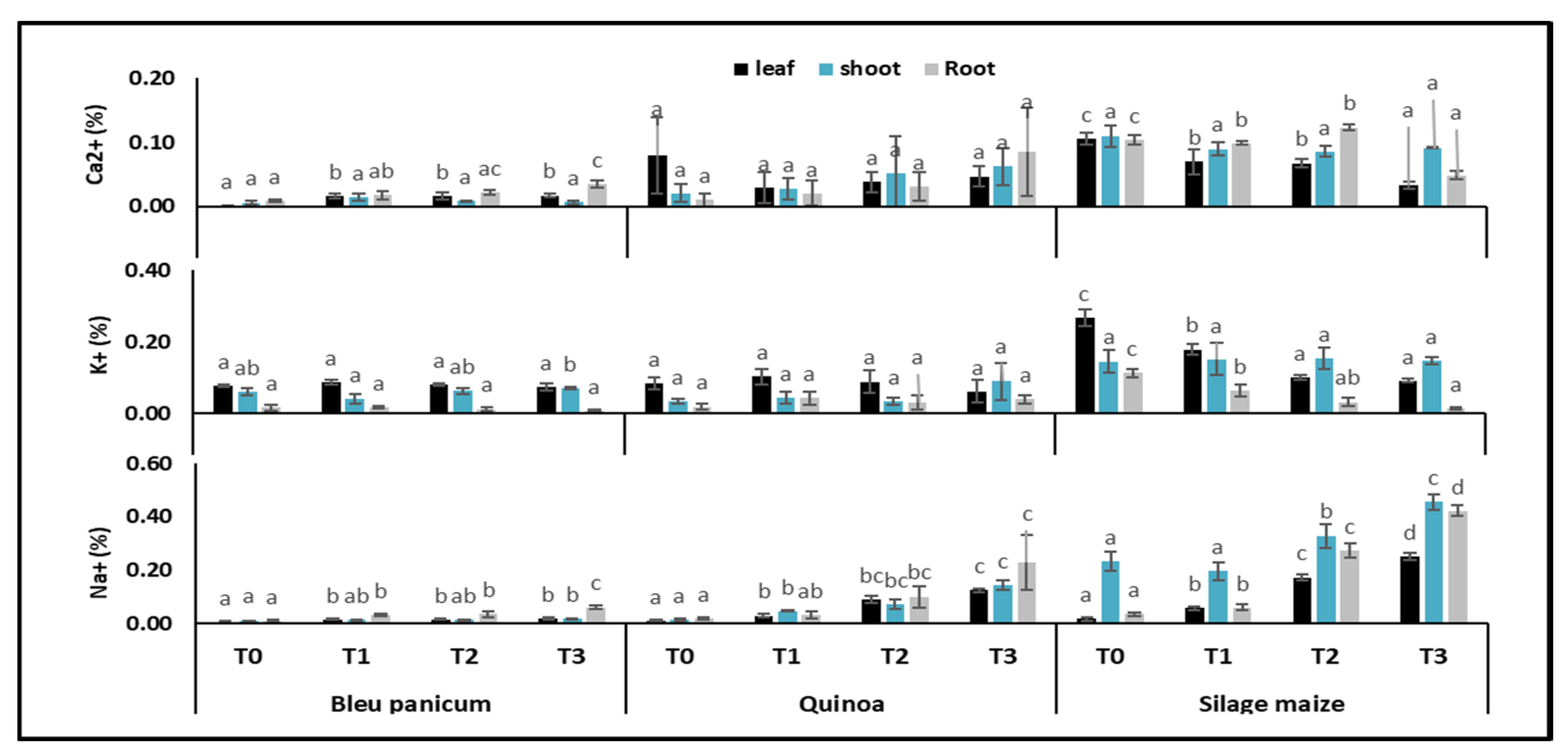
3.2. Plant Mineral Content
4. Discussion
4.1. Physiological and Biochemical Properties
4.2. Plant Mineral Content
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zaman, M.; Shahid, S.A.; Heng, L. Guideline for Salinity Assessment, Mitigation and Adaptation Using Nuclear and Related Techniques; Springer Nature: Cham, Switzerland, 2018; ISBN 3-319-96190-X. [Google Scholar]
- Oumasst, A.; Azougay, S.; Taqarort, N.; Mimouni, A.; Hallam, J. Effets de La Salinité Sur l’absorption Des Nutriments, Les Paramètres Biochimiques et La Croissance Du Bleu Panicum (Panicum Antidotale Retz) et Du Maïs d’ensilage (Zea Mays L). Afr. Mediterr. Agric. J.—Al Awamia 2021, 41–62. [Google Scholar] [CrossRef]
- Koyro, H.-W.; Hussain, T.; Huchzermeyer, B.; Khan, M.A. Photosynthetic and Growth Responses of a Perennial Halophytic Grass Panicum Turgidum to Increasing NaCl Concentrations. Environ. Exp. Bot. 2013, 91, 22–29. [Google Scholar] [CrossRef]
- Atia, A.; Debez, A.; Rabhi, M.; Barhoumi, Z.; Haouari, C.C.; Gouia, H.; Abdelly, C.; Smaoui, A. Salt Tolerance and Potential Uses for Saline Agriculture of Halophytes from the Poaceae. In Sabkha Ecosystems; Gul, B., Böer, B., Khan, M.A., Clüsener-Godt, M., Hameed, A., Eds.; Tasks for Vegetation Science; Springer International Publishing: Cham, Switzerland, 2019; Volume 49, pp. 223–237. ISBN 978-3-030-04416-9. [Google Scholar]
- Hirich, A.; Choukr-Allah, R.; Jacobsen, S.-E. Deficit Irrigation and Organic Compost Improve Growth and Yield of Quinoa and Pea. J. Agron. Crop Sci. 2014, 200, 390–398. [Google Scholar] [CrossRef]
- Santos, C.V. Regulation of Chlorophyll Biosynthesis and Degradation by Salt Stress in Sunflower Leaves. Sci. Hortic. 2004, 103, 93–99. [Google Scholar] [CrossRef]
- Roman, V.J. Salt Tolerance Strategies of the Ancient Andean Crop Quinoa; Wageningen University: Wageningen, The Netherlands, 2021. [Google Scholar] [CrossRef]
- Shabala, S. Learning from Halophytes: Physiological Basis and Strategies to Improve Abiotic Stress Tolerance in Crops. Ann. Bot. 2013, 112, 1209–1221. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
| Stem Height (cm) | Aboveground Dry Biomass (g) | Root Dry Biomass (g) | Chlorophyll a (µg mL−1) | Chlorophyll b (µg mL−1) | Carotenoid (µg mL−1) | Proline (mg g−1) | ||
|---|---|---|---|---|---|---|---|---|
| Blue panicum | T0 | 99.25 ± 10.63 ab | 11.71 ± 3.62 a | 4.2 ± 1.24 ab | 29.90 ± 2.69 a | 18.06 ± 4.34 a | 7.08 ± 0.86 a | 0.41 ± 0.18 a |
| T1 | 79.75 ± 5.25 a | 14.34 ± 2.85 a | 5.12 ± 0.88 b | 27.10 ± 5.62 a | 13.55 ± 4.71 a | 6.93 ± 0.70 a | 0.40 ± 0.14 a | |
| T2 | 107.25 ± 8.77 b | 12.69 ± 0.91 a | 3.3 ± 0.62 ab | 29.72 ± 5.66 a | 15.76 ± 2.20 a | 9.19 ± 0.48 b | 0.48 ± 0.14 a | |
| T3 | 98.75 ± 12.84 ab | 10.67 ± 0.51 a | 2.54 ± 0.71 a | 26.06 ± 3.03 a | 12.83 ± 4.26 a | 8.00 ± 1.29 ab | 0.58 ± 0.23 a | |
| Quinoa | T0 | 85.53 ± 6.97 a | 21.55 ± 1.27 a | 2.08 ± 0.39 ab | 21.62 ± 8.67 a | 43.68 ± 20.7 a | 2558.7 ± 1188 a | 0.25 ± 0.11 a |
| T1 | 61.96 ± 19.58 b | 20.98 ± 0.89 a | 2.34 ± 0.58 b | 19.84 ± 3.66 a | 28.47 ± 6.47 a | 1762.24 ± 386 a | 0.35 ± 0.01 a | |
| T2 | 60.16 ± 8.25 b | 21.32 ± 1.12 a | 1.75 ± 0.35 ab | 17.81 ± 4.25 a | 25.35 ± 5.75 a | 1572.33 ± 361 a | 0.35 ± 0.16 a | |
| T3 | 48.53 ± 7.39 c | 20.11 ± 1.48 a | 1.38 ± 0.24 a | 14.33 ± 3.58 a | 19.61 ± 4.33 a | 1225.31 ± 278 a | 0.37 ± 0.16 a | |
| Silage maize | T0 | 90.45 ± 4.96 c | 10.87 ± 0.79 c | 4.8 ± 0.65 d | 30.14 ± 2.89 c | 29.11 ± 6.06 c | 5.30 ± 1.80 ab | 0.71 ± 0.17 a |
| T1 | 85.35 ± 5.96 c | 8.32 ± 0.57 b | 3.622 ± 0.5 c | 28.39 ± 3.37 c | 18.51 ± 3.50 b | 7.48 ± 0.73 b | 0.62 ± 0.25 a | |
| T2 | 72.1 ± 4.09 b | 8.7 ± 1.18 b | 1.94 ± 0.57 b | 16.61 ± 2.17 b | 7.40 ± 1.77 a | 3.09 ± 0.52 a | 0.60 ± 0.08 a | |
| T3 | 56.95 ± 4.77 a | 4.61 ± 0.82 a | 0.64 ± 0.14 a | 8.84 ± 2.83 a | 6.64 ± 6.95 a | 2.72 ± 1.63 a | 0.33 ± 0.11 a | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oumasst, A.; Azougay, S.; Mimouni, A.; Hallam, J. The Effect of Different Irrigation Water Salinity Levels on Nutrients Uptake, Biochemical Content and Growth Response of Blue Panicum, Quinoa and Silage Maize. Environ. Sci. Proc. 2022, 16, 60. https://doi.org/10.3390/environsciproc2022016060
Oumasst A, Azougay S, Mimouni A, Hallam J. The Effect of Different Irrigation Water Salinity Levels on Nutrients Uptake, Biochemical Content and Growth Response of Blue Panicum, Quinoa and Silage Maize. Environmental Sciences Proceedings. 2022; 16(1):60. https://doi.org/10.3390/environsciproc2022016060
Chicago/Turabian StyleOumasst, Assma, Sara Azougay, Abdelaziz Mimouni, and Jamal Hallam. 2022. "The Effect of Different Irrigation Water Salinity Levels on Nutrients Uptake, Biochemical Content and Growth Response of Blue Panicum, Quinoa and Silage Maize" Environmental Sciences Proceedings 16, no. 1: 60. https://doi.org/10.3390/environsciproc2022016060
APA StyleOumasst, A., Azougay, S., Mimouni, A., & Hallam, J. (2022). The Effect of Different Irrigation Water Salinity Levels on Nutrients Uptake, Biochemical Content and Growth Response of Blue Panicum, Quinoa and Silage Maize. Environmental Sciences Proceedings, 16(1), 60. https://doi.org/10.3390/environsciproc2022016060