
Allometric Model for Predicting Root Biomass of Field Crops in the Salt-Affected Clay Soil: Novel Approach †



{kind=link}
{kind=link}
Abstract
:1. Introduction
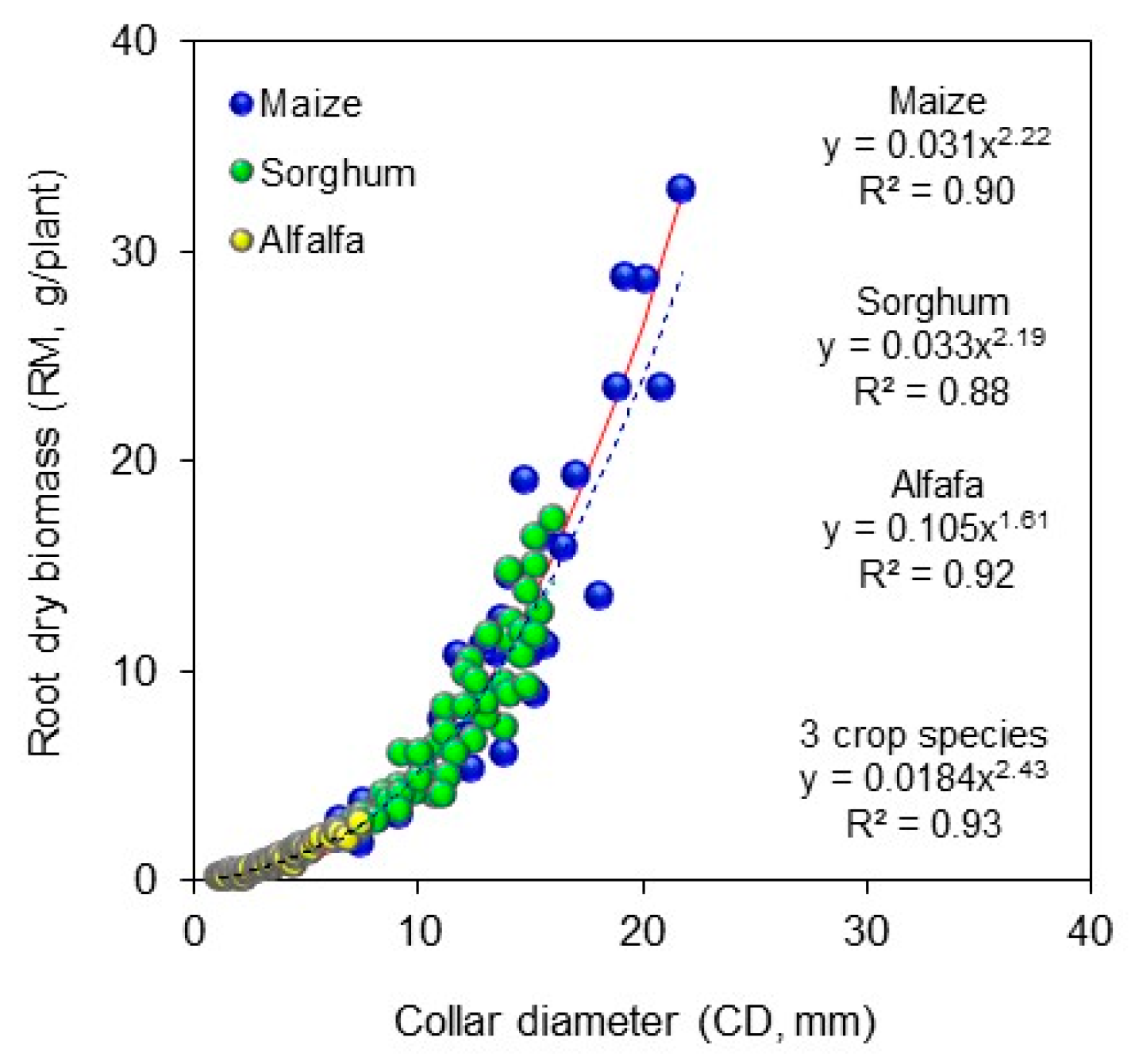
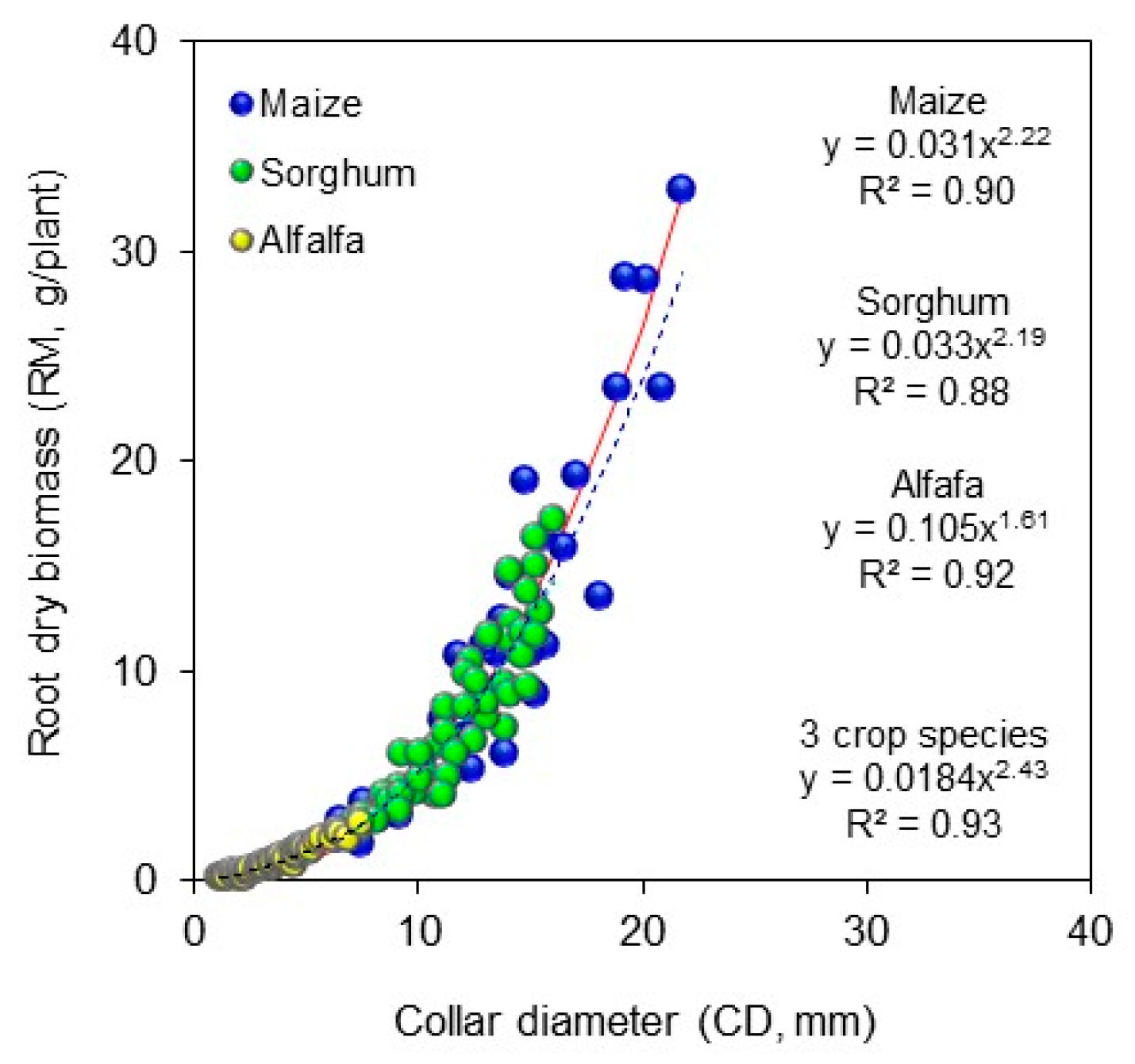
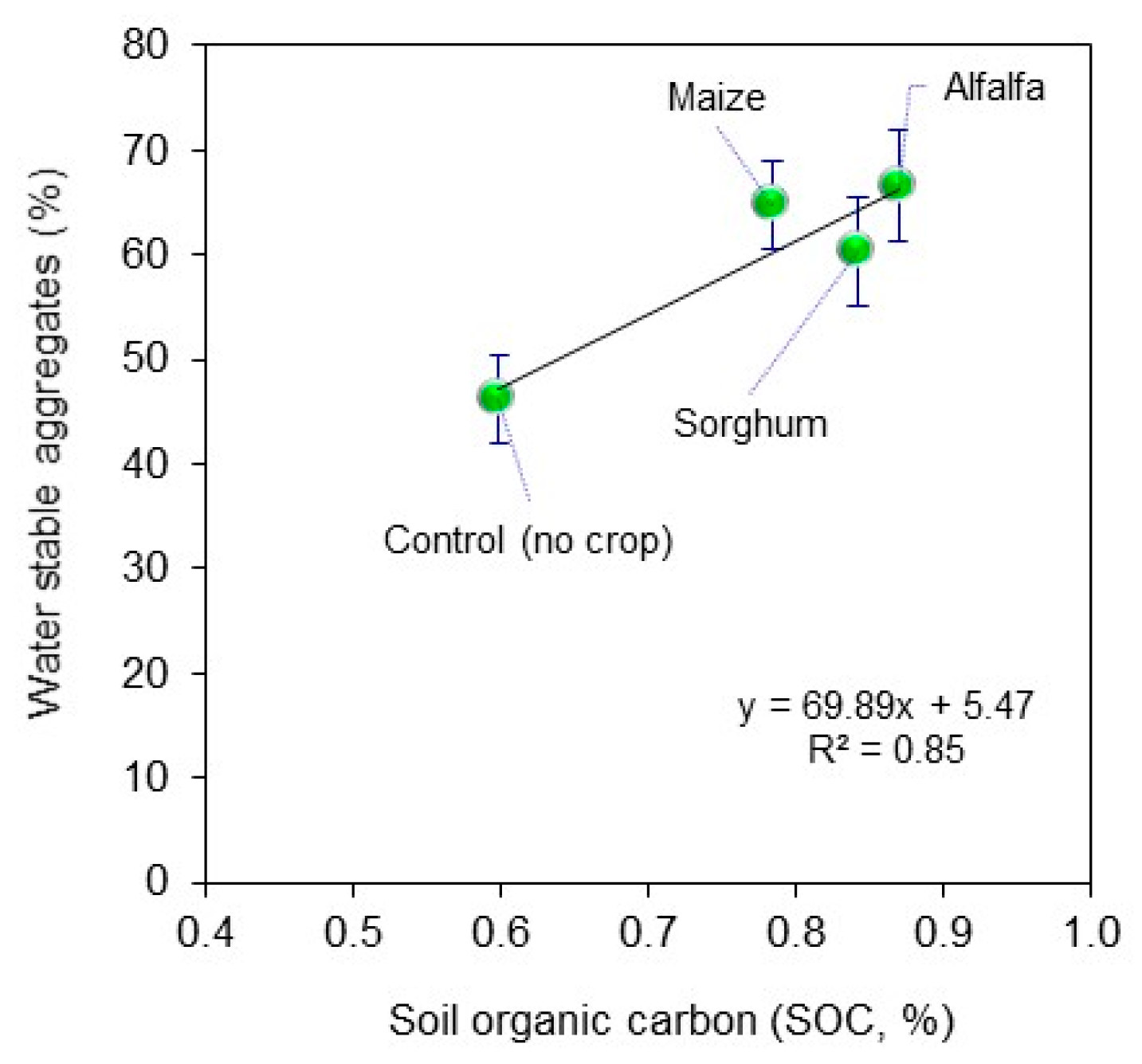
2. Material and Methods
3. Results and Discussions
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Powlson, D.S.; Gregory, P.J.; Whalley, W.R.; Quinton, J.N.; Hopkins, D.W.; Whitmore, A.P.; Hirsch, P.R.; Goulding, K.W.T. Soil management in relation to sustainable agriculture and ecosystem services. Food Policy 2011, 36, S72–S87. [Google Scholar] [CrossRef]
- Daynes, C.N.; Field, D.J.; Saleeba, J.A.; Cole, M.A.; McGee, P.A. Development and stabilisation of soil structure via interactions between organic matter, arbuscular mycorrhizal fungi and plant roots. Soil Biol. Biochem. 2013, 57, 683–694. [Google Scholar] [CrossRef]
- Roumet, C.; Birouste, M.; Picon-Cochard, C.; Ghestem, M.; Osman, N.; Vrignon-Brenas, S.; Cao, K.; Stokes, A. Root structure–function relationships in 74 species: Evidence of a root economics spectrum related to carbon economy. New Phytol. 2016, 210, 815–826. [Google Scholar] [CrossRef] [PubMed]
- Faucon, M.P.; Houben, D.; Lambers, H. Plant functional traits: Soil and ecosystem services. Trends Plant Sci. 2017, 22, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Hirte, J.; Leifeld, J.; Abiven, S.; Oberholzer, H.-R.; Mayer, J. Below ground carbon inputs to soil via root biomass and rhizodeposition of field-grown maize and wheat at harvest are independent of net primary productivity. Agric. Ecosyst. Environ. 2018, 265, 556–566. [Google Scholar] [CrossRef]
- Dijkstra, F.A.; Zhu, B.; Cheng, W. Root effects on soil organic carbon: A double-edged sword. New Phytol. 2021, 230, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Freschet, G.T.; Pagès, L.; Iversen, C.M.; Comas, L.H.; Rewald, B.; Roumet, C.; Klimešová, J.; Zadworny, M.; Poorter, H.; Postma, J.A.; et al. A starting guide to root ecology: Strengthening ecological concepts and standardising root classification, sampling, processing and trait measurements. New Phytol. 2021, 232, 973–1122. [Google Scholar] [CrossRef] [PubMed]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. Tansley Review. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef] [PubMed]
- Addo-Danso, S.D.; Prescott, C.E.; Smith, A.R. Methods for estimating root biomass and production in forest and woodland ecosystem carbon studies: A review. For. Ecol. Manag. 2015, 359, 332–351. [Google Scholar] [CrossRef]
- Marden, M.; Lambie, S.; Phillips, C. Biomass and root attributes of eight of New Zealand’s most common indigenous evergreen conifer and broadleaved forest species during the first 5 years of establishment. N. Z. J. For. Sci. 2018, 48, 9. [Google Scholar] [CrossRef]
- Fageria, N.K. The Role of Plant Roots in Crop Production; CRC Press: Boca Raton, FL, USA, 2012; p. 467. [Google Scholar] [CrossRef]
- Reddy, V.R.; Pachepsky, Y.A.; Whisler, F.D. Allometric relationships in field grown soybean. Ann. Bot. 1998, 82, 125–131. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Rui, Y.; Birch, C.; Hanan, J. Allometric relationships of maize organ development under different water regimes and plant densities. Plant Prod. Sci. 2015, 18, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Mamedov, A.I.; Gasimova, K.; Husiyev, E.K.; Farzaliyev, V.; Alizade, V.M.; Toderich, K. Root and Shoot Relation of the Quinoa and Forage Plants in Salt-Affected Clay Soil. In Emerging Research in Alternative Crops. Environment & Policy; Hirich, A., Choukr-Allah, R., Ragab, R., Eds.; Springer: Cham, Switzerland, 2020; Volume 58, pp. 185–220. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mamedov, A.I.; Husiyev, E.K. Allometric Model for Predicting Root Biomass of Field Crops in the Salt-Affected Clay Soil: Novel Approach. Environ. Sci. Proc. 2022, 16, 11. https://doi.org/10.3390/environsciproc2022016011
Mamedov AI, Husiyev EK. Allometric Model for Predicting Root Biomass of Field Crops in the Salt-Affected Clay Soil: Novel Approach. Environmental Sciences Proceedings. 2022; 16(1):11. https://doi.org/10.3390/environsciproc2022016011
Chicago/Turabian StyleMamedov, Amrakh I., and Emin K. Husiyev. 2022. "Allometric Model for Predicting Root Biomass of Field Crops in the Salt-Affected Clay Soil: Novel Approach" Environmental Sciences Proceedings 16, no. 1: 11. https://doi.org/10.3390/environsciproc2022016011
APA StyleMamedov, A. I., & Husiyev, E. K. (2022). Allometric Model for Predicting Root Biomass of Field Crops in the Salt-Affected Clay Soil: Novel Approach. Environmental Sciences Proceedings, 16(1), 11. https://doi.org/10.3390/environsciproc2022016011