

The Impact of Land Cover on Nest Occupancy of the White Stork (Ciconia ciconia (L.)): A Case Study of Kampinos Forest, 2006–2018


 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
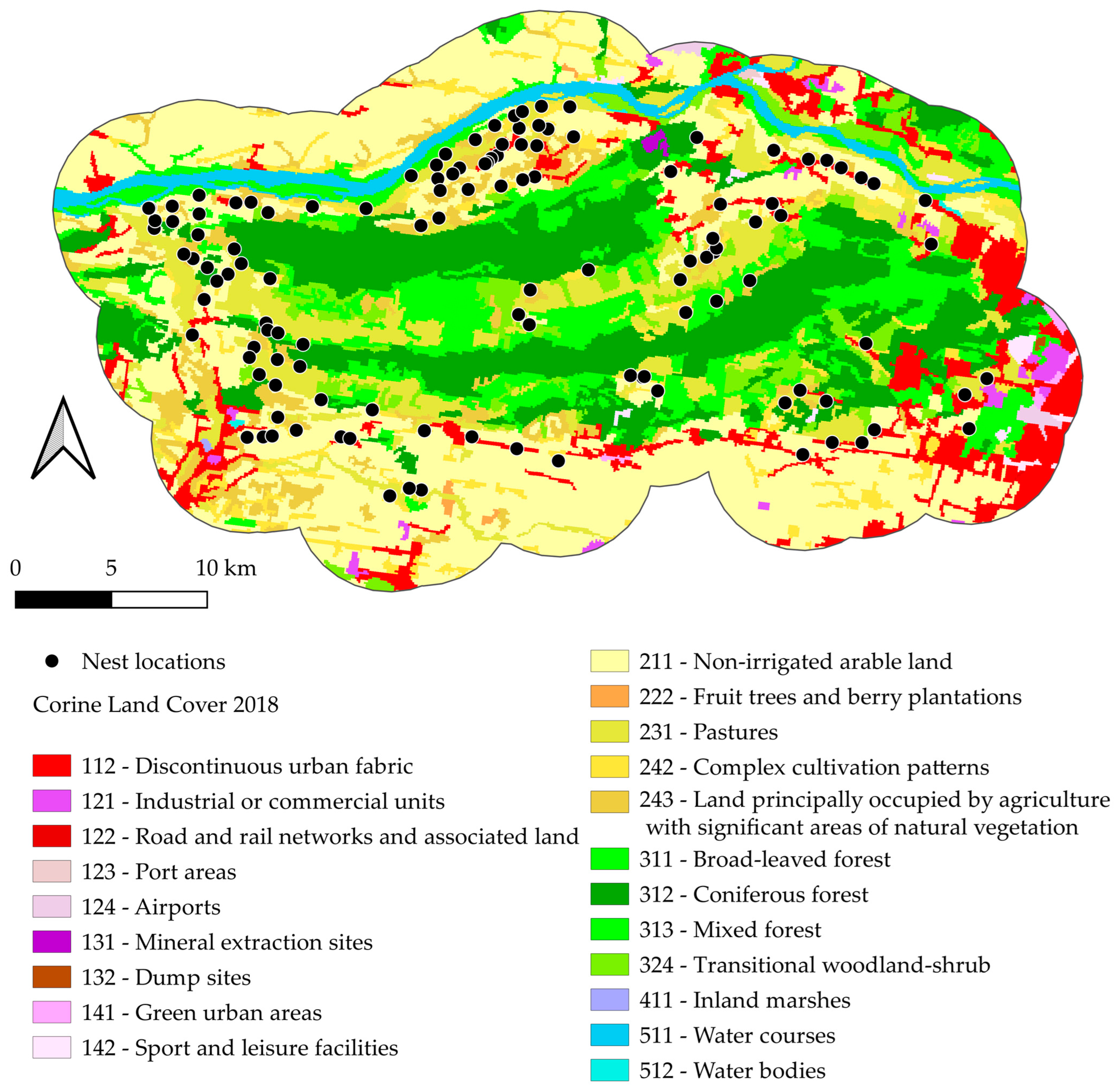
2.1. Research Area
2.2. Data on the White Stork
2.3. CORINE Land Cover
2.4. Statistical and Spatial Analyses
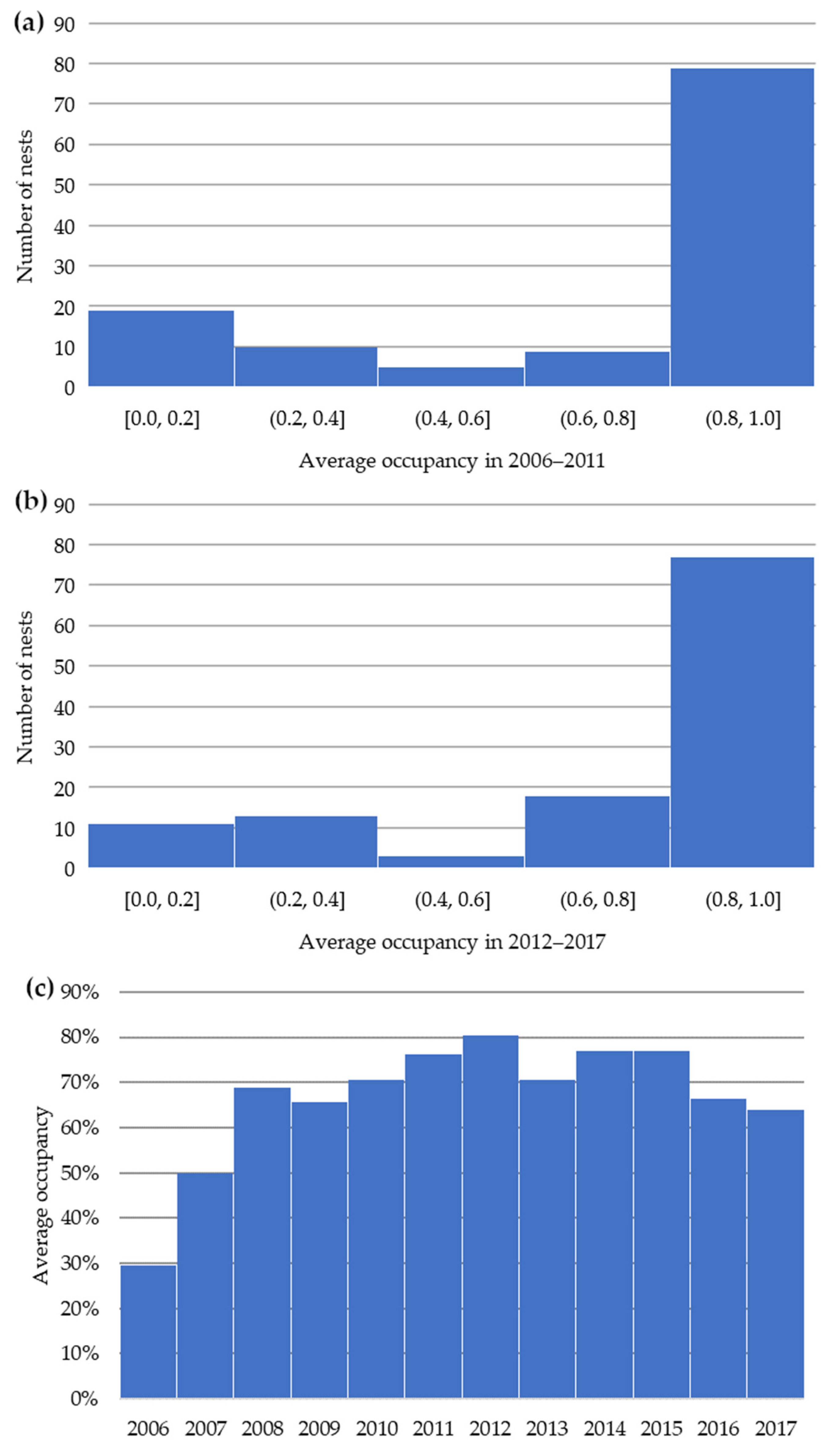
3. Results
3.1. Results for Level 1 CORINE Land Cover
3.2. Results for Level 2 CORINE Land Cover
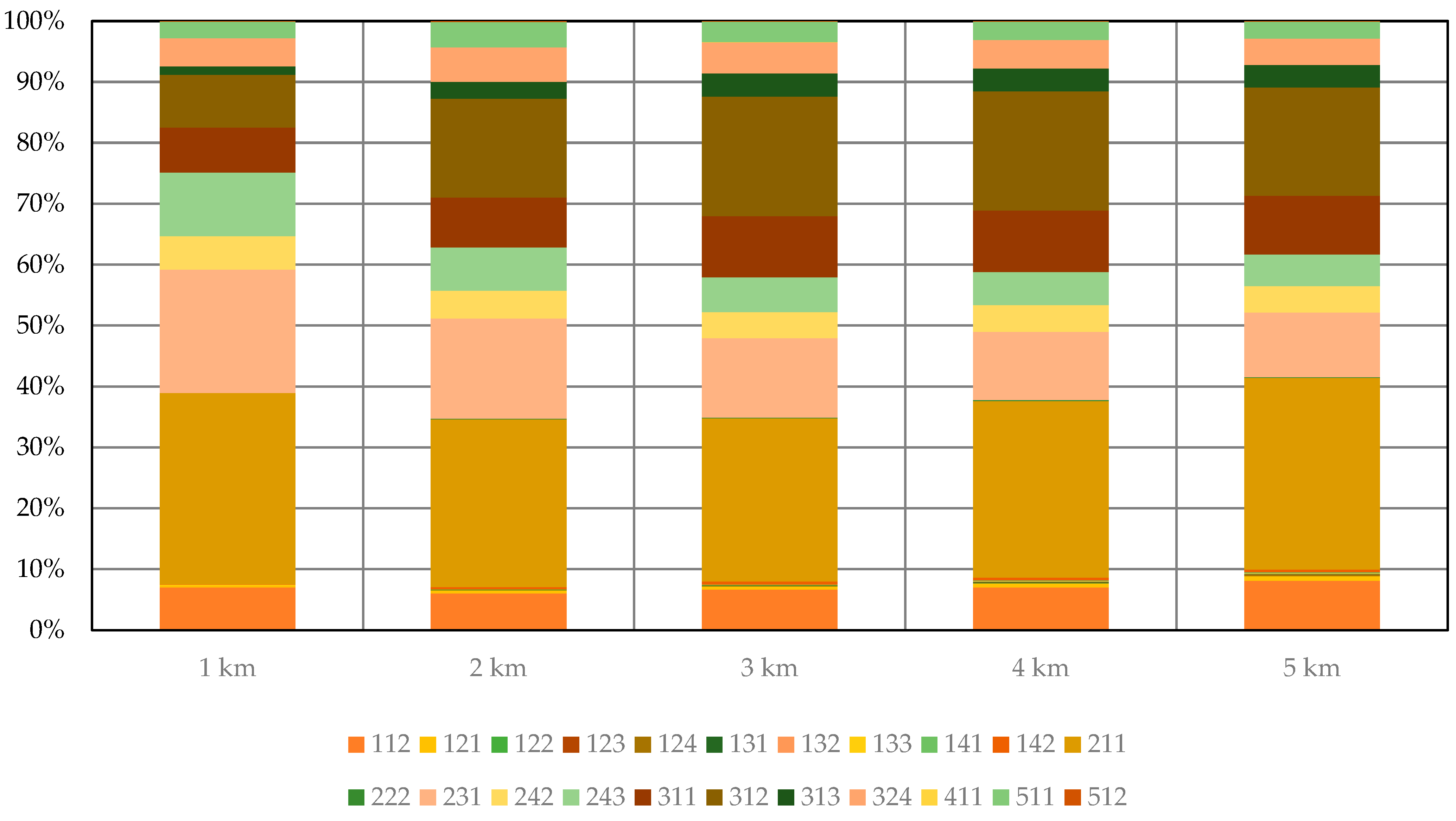
3.3. Results for Level 3 CORINE Land Cover
4. Discussion
5. Conclusions
- In the study area, white stork nest occupancy is most influenced by anthropogenic areas, agricultural areas, and forests and semi-natural ecosystems.
- Only land principally occupied by agriculture, with significant areas of natural vegetation (243) and non-irrigated arable land (211) show a positive influence on nest occupancy.
- The strength of the correlation of each land cover category depends on the distance from the nests, generally monotonically increasing or decreasing with distance.
- In the study area, land covered with wetlands and water (4 and 5) was not a significant factor influencing the location of white stork nests.
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zawadzka, D.; Zawadzki, J. Ptaki Jako Gatunki Wskaznikowe Roznorodnosci Biologicznej i Stopnia Naturalnosci Lasow. Stud. Mater. Cent. Edukac. Przyr. Śnej 2006, 8, 249–262. [Google Scholar]
- Lifshitz, N.; St. Clair, C.C. Iridescent Coloration of Tree Swallows Relates to Environmental Metal Pollution. Avian Conserv. Ecol. 2019, 14, art7. [Google Scholar] [CrossRef]
- Hagar, J.; Branch, E. Western Purple Martin (Progne Subis Arboricola) Occurrence on the Siuslaw National Forest, Summer 2019; U.S. Geological Survey: Reston, VA, USA, 2020; Open-File Report. [CrossRef]
- Flesch, A.D. Landcover Change and Habitat Quality Mediate Impacts of Temperature and Precipitation on Population Dynamics of a Threatened Aridland Predator. Anim. Conserv. 2023, 26, 464–476. [Google Scholar] [CrossRef]
- Kuczyński, L.; Antczak, M.; Czechowski, P.; Grzybek, J.; Jerzak, L.; Zabłocki, P.; Tryjanowski, P. A Large Scale Survey of the Great Grey Shrike Lanius Excubitor in Poland: Breeding Densities, Habitat Use and Population Trends. Ann. Zool. Fenn. 2010, 47, 67–78. [Google Scholar] [CrossRef]
- Morelli, F. Plasticity of Habitat Selection By Red-Backed Shrikes (Lanius Collurio) Breeding In Different Landscapes. Wilson J. Ornithol. 2012, 124, 51–56. [Google Scholar] [CrossRef]
- Frederick, P.; Gawlik, D.E.; Ogden, J.C.; Cook, M.I.; Lusk, M. The White Ibis and Wood Stork as Indicators for Restoration of the Everglades Ecosystem. Ecol. Indic. 2009, 9, S83–S95. [Google Scholar] [CrossRef]
- Faria, D.; Morante-Filho, J.C.; Baumgarten, J.; Bovendorp, R.S.; Cazetta, E.; Gaiotto, F.A.; Mariano-Neto, E.; Mielke, M.S.; Pessoa, M.S.; Rocha-Santos, L.; et al. The Breakdown of Ecosystem Functionality Driven by Deforestation in a Global Biodiversity Hotspot. Biol. Conserv. 2023, 283, 110126. [Google Scholar] [CrossRef]
- Dolata, P.T. The White Stork Ciconia ciconia Protection in Poland by Tradition, Customs, Law, and Active Efforts. In The White Stork in Poland: Studies in Biology, Ecology and Conservation; Tryjanowski, P., Sparks, T.H., Jerzak, L., Eds.; Bogucki Wydawnictwo Naukowe: Poznań, Poland, 2006; pp. 477–492. ISBN 978-83-60247-35-8. [Google Scholar]
- Hancock, J.; Kushlan, J.A.; Kahl, M.P. Storks, Ibises and Spoonbills of the World; Christopher Helm Publishers: London, UK, 2010; ISBN 978-1-4081-3499-3. [Google Scholar]
- Flack, A.; Nagy, M.; Fiedler, W.; Couzin, I.D.; Wikelski, M. From Local Collective Behavior to Global Migratory Patterns in White Storks. Science 2018, 360, 911–914. [Google Scholar] [CrossRef] [PubMed]
- Martín, B.; Onrubia, A.; Ferrer, M. Climate Change and the Spatiotemporal Variation in Survival of a Long-Distance Migrant (White Stork, Ciconia ciconia) across Western Europe. Birds 2021, 2, 362–380. [Google Scholar] [CrossRef]
- Carrascal, L.M.; Bautista, L.M.; Lázaro, E. Geographical Variation in the Density of the White Stork Ciconia ciconia in Spain: Influence of Habitat Structure and Climate. Biol. Conserv. 1993, 65, 83–87. [Google Scholar] [CrossRef]
- Latus, C.; Kujawa, K. The Effect of Land Cover and Fragmentation of Agricultural Landscape on the Density of White Stork [Ciconia ciconia L.] in Brandenburg, Germany. Pol. J. Ecol. 2005, 4, 535–543. [Google Scholar]
- Olsson, O.; Rogers, D.J. Predicting the Distribution of a Suitable Habitat for the White Stork in Southern Sweden: Identifying Priority Areas for Reintroduction and Habitat Restoration. Anim. Conserv. 2009, 12, 62–70. [Google Scholar] [CrossRef]
- Chylarecki, P.; Sikora, A.; Cenian, Z.; Chodkiewicz, T. Monitoring Ptaków Lęgowych: Poradnik Metodyczny: Praca Zbiorowa, 2nd ed.; GIOŚ: Warszawa, Poland, 2015; ISBN 978-83-61227-45-8. [Google Scholar]
- Gyalus, A.; Végvári, Z.; Csörgő, T. Changes in the Nest Sites of White Stork in Hungary. Ornis Hung. 2018, 26, 65–88. [Google Scholar] [CrossRef]
- Janiszewski, T.; Minias, P.; Wojciechowski, Z.; Podlaszczuk, P. Habitat Selection by White Storks Breeding in a Mosaic Agricultural Landscape of Central Poland. Wilson J. Ornithol. 2014, 126, 591–599. [Google Scholar] [CrossRef]
- Kuźniak, S.; Tobółka, M. Spadek Liczebności Bociana Białego Ciconia ciconia Na Ziemi Leszczyńskiej i Program Jego Ochrony. Chrońmy Przyr Ojcz 2010, 66, 97–106. [Google Scholar]
- Wuczyński, A.; Krogulec, G.; Neubauer, G.; Profus, P. Pierwsza Ocena Liczebności Bociana Białego Ciconia ciconia w Polsce w 1958 Roku–Streszczenie Prezentacji. Chrońmy Przyr Ojcz 2019, 75, 336–344. [Google Scholar]
- Białas, J.T.; Dylewski, Ł.; Tobółka, M. Determination of Nest Occupation and Breeding Effect of the White Stork by Human-Mediated Landscape in Western Poland. Environ. Sci. Pollut. Res. 2020, 27, 4148–4158. [Google Scholar] [CrossRef] [PubMed]
- Tobolka, M.; Sparks, T.H.; Tryjanowski, P. Brief Report Does the White Stork Ciconia ciconia Reflect Farmland Bird Diversity? Ornis Fenn. 2012, 89, 222–228. [Google Scholar] [CrossRef]
- Szyszko, J.; Schwerk, A.; Malczyk, J. Animals as an Indicator of Carbon Sequestration and Valuable Landscapes. ZooKeys 2011, 100, 565–573. [Google Scholar] [CrossRef]
- Kronenberg, J.; Bocheński, M.; Dolata, P.; Jerzak, L.; Profus, P.; Tobółka, M.; Tryjanowski, P.; Wuczyński, A.; Żołnierowicz, K. Znaczenie Bociana Białego Ciconia ciconia Dla Społeczeństwa: Analiza z Perspektywy Koncepcji Usług Ekosystemów. Chrońmy Przyr. Ojczystą 2013, 69, 179–203. [Google Scholar]
- Romero Lopez, J.M. Cooperación y Conflicto Entre Hermanos de Nido En Cigüeña Blanca Ciconia ciconia; Universidad Pablo de Olavide: Sevilla, Spain, 2017. [Google Scholar]
- Zurell, D.; von Wehrden, H.; Rotics, S.; Kaatz, M.; Groß, H.; Schlag, L.; Schäfer, M.; Sapir, N.; Turjeman, S.; Wikelski, M.; et al. Home Range Size and Resource Use of Breeding and Non-Breeding White Storks Along a Land Use Gradient. Front. Ecol. Evol. 2018, 6, 79. [Google Scholar] [CrossRef]
- Dziewiaty, K.; Schulz, H. Störche in der Elbtalaue; K & W, Dr.- und Verl.-Haus: Schleswig, Germany, 1998; ISBN 978-3-88242-128-6. [Google Scholar]
- Podlaszczuk, M.; Wojciechowski, Z.; Podlaszczuk, P.; Minias, P.; Janiszewski, T.; Wojciechowska, A. Shortening Day Length as a Previously Unrecognized Selective Pressure for Early Breeding in a Bird with Long Parental Care. J. Ornithol. 2015, 156, 389–396. [Google Scholar] [CrossRef]
- Sikora, A.; Chodkiewicz, T.; Rohde, Z.; Lewandowska, J. Monitoring Ptaków z Uwzględnieniem Obszarów Specjalnej Ochrony Ptaków Natura 2000, Lata 2018–2021; Część I. Monitoring ptaków pospolitych, terenów podmokłych i leśnych z uwzględnieniem obszarów specjalnej ochrony ptaków Natura 2000, lata 2018–2021; GIOŚ: Marki, Gdańsk, Warszawa, Poland, 2021. [Google Scholar]
- Copernicus Programme CORINE Land Cover—Copernicus Land Monitoring Service. Available online: https://land.copernicus.eu/pan-european/corine-land-cover (accessed on 26 February 2023).
- Buda, A.; Błażejewska, S.; Jarynowski, A. Life Time of Correlations and Its Applications. Vol. 1; Andrzej Buda Wydawnictwo Niezależne: Głogów, Poland, 2010; ISBN 978-83-915272-9-0. [Google Scholar]
- Fisher, R.A. Statistical Methods for Research Workers; Biological monographs and manuals; Oliver and Boyd: Edinburg, UK, 1925. [Google Scholar]
- QGIS Development Team. QGIS Geographic Information System Ver 3.22 Białowieża; QGIS Association, 2021. [Google Scholar]
- Virtanen, P.; Gommers, R.; Oliphant, T.E.; Haberland, M.; Reddy, T.; Cournapeau, D.; Burovski, E.; Peterson, P.; Weckesser, W.; Bright, J.; et al. SciPy 1.0: Fundamental Algorithms for Scientific Computing in Python. Nat. Methods 2020, 17, 261–272. [Google Scholar] [CrossRef]
- Reback, J.; Jbrockmendel; McKinney, W.; Van Den Bossche, J.; Roeschke, M.; Augspurger, T.; Hawkins, S.; Cloud, P.; Gfyoung; Sinhrks; et al. Pandas-Dev/Pandas: Pandas 1.4.3. Zenodo 2022. [Google Scholar] [CrossRef]
- Harris, C.R.; Millman, K.J.; Van Der Walt, S.J.; Gommers, R.; Virtanen, P.; Cournapeau, D.; Wieser, E.; Taylor, J.; Berg, S.; Smith, N.J.; et al. Array Programming with NumPy. Nature 2020, 585, 357–362. [Google Scholar] [CrossRef]
- Van Rossum, G.; Drake, F.L. The Python Language Reference; Python documentation manual/Guido van Rossum; Drake, F.L., Ed.; Release 3.0.1 [Repr.]; Python Software Foundation: Hampton, NH, USA, 2010; ISBN 978-1-4414-1269-0. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Otręba, A.; Ferchmin, M.; Kębłowska, A.; Kloss, M.; Michalska-Hejduk, D. Ochrona Flory i Roślinności w Kampinoskim Parku Narodowym. In Z Mazowsza Na Polesie Wileńszczyznę; Obidziński, A., Ed.; Polskie Towarzystwo Botaniczne—Zarząd Główny: Warsaw, Poland, 2010; pp. 67–94. ISBN 978-83-86292-78-3. [Google Scholar]
- Solon, J.; Otręba, A.; Andrzejewska, A.; Zaniewski, P.T. Zmiany składu gatunkowego i wskaźników struktury zbiorowisk roślinnych w latach 2001–2018 na wybranych powierzchniach stałych w Kampinoskim Parku Narodowym = Changes of species composition and structural indices among plant communities in 2001–2018 on selected permanent plots in Kampinos National Park. Przegląd Geogr. 2021, 93, 413–443. [Google Scholar] [CrossRef]
- Rozporządzenie Rady Ministrów z Dnia 16 Stycznia 1959 r. w Sprawie Utworzenia Kampinoskiego Parku Narodowego (Ordinance of the Council of Ministers of 16 January 1959 on the establishment of the Kampinos National Park.). Dziennik Ustaw Polskiej Rzeczypospolitej Ludowej, Warsaw, Poland, 1959; Dz.U. 1959 nr 17 poz. 91. Available online: https://isap.sejm.gov.pl/isap.Nsf/DocDetails.xsp?id=WDU19590170091 (accessed on 29 May 2024).
- Rozporządzenie Rady Ministrów z Dnia 25 Września 1997 r. w Sprawie Kampinoskiego Parku Narodowego (Ordinance of the Council of Ministers of September 25, 1997 on the Kampinos National Park.). Dziennik Ustaw, Warsaw, Poland, 1997; Dz.U. 1997 nr 132 poz. 876. Available online: https://isap.sejm.gov.pl/isap.nsf/DocDetails.xsp?id=WDU19971320876 (accessed on 29 May 2024).
- Ustawa z Dnia 16 Kwietnia 2004 r. o Ochronie Przyrody (Act of 16 April 2004 on Nature Protection.). Dziennik Ustaw, Warsaw, Poland, 2023; Dz.U. 2023 poz. 1336. Available online: https://isap.sejm.gov.pl/isap.nsf/DocDetails.xsp?id=WDU20230001336 (accessed on 29 May 2024).
- KPN Ochrona Przyrody. Nature Conservation. Available online: https://kampn.gov.pl/ochrona-przyrody (accessed on 23 June 2024).
- KPN Plan Ochrony KPN. Conservation Plan Kampinos National Park. Available online: https://bip.kampinoski-pn.gov.pl/index.php/do-cignicia/Plan-Ochrony-KPN/ (accessed on 30 July 2024).
- Johst, K.; Brandl, R.; Pfeifer, R. Foraging in a Patchy and Dynamic Landscape: Human Land Use and the White Stork. Ecol. Appl. 2001, 11, 60–69. [Google Scholar] [CrossRef]
- Janiszewski, T.; Minias, P.; Wojciechowska, A.; Wojciechowski, Z. Wyniki Dwudziestoletniego Monitoringu Populacji Bociana Białego Ciconia ciconia Na Ziemi Łowickiej (1994–2013)–Zmiany Liczebności, Efektów Lęgów i Sposobu Gniazdowania. Chrońmy Przyr. Ojczystą 2014, 70, 312–321. [Google Scholar]
- Bihałowicz, J.; Bihałowicz, J. Uncertainties in Land Cover. The Mutual Validation of Remote Sensing Land Cover and Topographic Map. In Proceedings of the 3rd International Seminar and Workshop on Geoinformatics, Remote Sensing and Engineering in Spatial Planning and Environmental Science, Warsaw, Poland, 3–4 October 2022. [Google Scholar]
- Radović, A.; Tepić, N. Using CORINE Land Cover Habitat Database for the Analysis of Breeding Bird Habitat: Case Study of White Storks (Ciconia ciconia) from Northern Croatia. Biologia 2009, 64, 1212–1218. [Google Scholar] [CrossRef]
- Orłowski, G.; Karg, J.; Jerzak, L.; Bocheński, M.; Profus, P.; Książkiewicz-Parulska, Z.; Zub, K.; Ekner-Grzyb, A.; Czarnecka, J. Linking Land Cover Satellite Data with Dietary Variation and Reproductive Output in an Opportunistic Forager: Arable Land Use Can Boost an Ontogenetic Trophic Bottleneck in the White Stork Ciconia ciconia. Sci. Total Environ. 2019, 646, 491–502. [Google Scholar] [CrossRef]
- Sikora, A. Rozmieszczenie, Zmiany Liczebności i Produktywność Bociana Białego Ciconia ciconia Na Żuławach Wiślanych. Chrońmy Przyr. Ojczystą 2017, 73, 363–378. [Google Scholar]
- Pestka, Z.; Zbyryt, A.; Menderski, S.; Jakubas, D. Habitat Suitability Mapping of White Stork Ciconia ciconia in One of Its Key European Breeding Areas. Ecol. Indic. 2023, 151, 110278. [Google Scholar] [CrossRef]
- Tryjanowski, P.; Grzywaczewski, G.; Zbyryt, A. Foraging of White Stork Ciconia ciconia in Forests—The Heritage of an Ancient Behaviour? Pol. J. Ecol. 2018, 66, 250–256. [Google Scholar] [CrossRef]
- Nowakowski, J.J. Habitat Structure and Breeding Parameters of the White Stork Ciconia ciconia in the Kolno Upland (NE Poland). Acta Ornithol. 2003, 38, 39–46. [Google Scholar] [CrossRef]
- Janiszewski, T.; Minias, P.; Wojciechowski, Z. Occupancy Reliably Reflects Territory Quality in a Long-lived Migratory Bird, the White Stork. J. Zool. 2013, 291, 178–184. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter Value | Code from the Inventory | Explanation |
|---|---|---|
| Nest unoccupied (0) | HO | The nest remained unoccupied, and no storks were observed to have arrived. |
| HB1 | The nest was visited by a single stork for a period of one week to one month, or alternatively, was occupied intermittently during the breeding season. | |
| HB2 | The nest was visited by two storks for a period of one to two weeks, or intermittently during the breeding season. | |
| HBx | The nest was visited for a period of between one and two weeks, although the number of birds present at the time is unknown. | |
| Nest occupied (1) | HP | The nest was occupied by a pair of birds for a period of more than one month between the dates of 14 April and 15 June. |
| LEVEL 1 | LEVEL 2 | LEVEL 3 | |||
|---|---|---|---|---|---|
| CODE | NAME | CODE | NAME | CODE | NAME |
| 1 | Artificial surfaces | 11 | Urban fabric | 112 | Discontinuous urban fabric |
| 12 | Industrial, commercial, and transport units | 121 | Industrial or commercial units | ||
| 122 | Road and rail networks and associated land | ||||
| 123 | Port areas | ||||
| 124 | Airports | ||||
| 13 | Mine, dump and construction sites | 131 | Mineral extraction sites | ||
| 132 | Dump sites | ||||
| 133 | Construction sites | ||||
| 14 | Artificial, non-agricultural vegetated areas | 141 | Green urban areas | ||
| 142 | Port and leisure facilities | ||||
| 2 | Agricultural areas | 21 | Arable land | 211 | Non-irrigated arable land |
| 22 | Permanent crops | 222 | Fruit trees and berry plantations | ||
| 23 | Pastures | 231 | Pastures | ||
| 24 | Heterogeneous agricultural areas | 242 | Complex cultivation patterns | ||
| 243 | Land principally occupied by agriculture, with significant areas of natural vegetation | ||||
| 3 | Forest and semi-natural areas | 31 | Forests | 311 | Broad-leaved forest |
| 312 | Coniferous forest | ||||
| 313 | Mixed forest | ||||
| 32 | Scrub and/or herbaceous vegetation associations | 324 | Transitional woodland-shrub | ||
| 4 | Wetlands | 41 | Inland wetlands | 411 | Inland marshes |
| 5 | Reservoirs | 51 | Inland waters | 511 | Water courses |
| 512 | Water bodies | ||||
| CLC I | 2006–2011 | 2012–2017 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 km | 2 km | 3 km | 4 km | 5 km | 1 km | 2 km | 3 km | 4 km | 5 km | |
| 1—Artificial surfaces | −0.031 | −0.050 | −0.022 | −0.021 | −0.033 | −0.028 | −0.197 * | −0.225 * | −0.297 *** | −0.313 *** |
| 2—Agricultural areas | 0.169 ᵗʳ | 0.098 | 0.084 | 0.114 | 0.157 ᵗʳ | 0.315 *** | 0.272 ** | 0.201 * | 0.157 ᵗʳ | 0.114 |
| 3—Forest and semi-natural areas | −0.177 ᵗʳ | −0.116 | −0.097 | −0.121 | −0.155 ᵗʳ | −0.309 *** | −0.212 * | −0.14 | −0.066 | −0.001 |
| 4—Wetlands | 0.060 | 0.081 | 0.101 | 0.127 | 0.074 | 0.045 | −0.052 | −0.037 | ||
| 5—Water bodies | 0.086 | 0.112 | 0.091 | 0.086 | 0.079 | 0.028 | 0.060 | 0.100 | 0.108 | 0.106 |
| CLC II | 2006–2011 | 2012–2017 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 km | 2 km | 3 km | 4 km | 5 km | 1 km | 2 km | 3 km | 4 km | 5 km | |
| 11—Urban fabric | −0.043 | −0.057 | −0.022 | −0.02 | −0.019 | 0.017 | −0.139 | −0.180 * | −0.273 ** | −0.304 *** |
| 12—Industrial, commercial, and transport units | 0.051 | 0.018 | 0.021 | 0.004 | −0.073 | −0.251 ** | −0.336 *** | −0.331 *** | −0.313 *** | −0.324 *** |
| 13—Mines, dump sites, and construction sites | 0.070 | −0.029 | −0.105 | −0.106 | −0.104 | −0.028 | −0.015 | −0.004 | −0.034 | −0.043 |
| 14—Artificial, non-agricultural vegetated areas | −0.184 * | −0.035 | 0.011 | −0.001 | −0.020 | −0.109 | −0.229 * | −0.191 * | −0.273 ** | −0.265 ** |
| 21—Arable land | 0.097 | 0.104 | 0.100 | 0.094 | 0.112 | 0.245 ** | 0.235 ** | 0.159 ᵗʳ | 0.105 | 0.074 |
| 22—Permanent crops | 0.052 | 0.019 | 0.016 | 0.039 | 0.047 | −0.022 | −0.007 | 0.001 | ||
| 23—Pastures | 0.038 | −0.065 | −0.078 | −0.029 | 0.022 | −0.075 | −0.055 | −0.044 | −0.034 | −0.021 |
| 24—Heterogeneous agricultural areas | 0.038 | 0.083 | 0.083 | 0.115 | 0.138 | 0.144 | 0.178 * | 0.198 * | 0.201 * | 0.153 ᵗʳ |
| 31—Forests | −0.148 | −0.070 | −0.060 | −0.095 | −0.127 | −0.338 *** | −0.240 ** | −0.144 | −0.061 | 0.010 |
| 32—Scrub and/or herbaceous vegetation associations | −0.120 | −0.180 * | −0.181 * | −0.179 * | −0.217 * | −0.043 | −0.024 | −0.048 | −0.055 | −0.052 |
| 41—Inland wetlands | 0.060 | 0.081 | 0.101 | 0.127 | 0.074 | 0.045 | −0.052 | −0.037 | ||
| 51—Inland waters | 0.086 | 0.112 | 0.091 | 0.086 | 0.079 | 0.028 | 0.060 | 0.100 | 0.108 | 0.106 |
| CLC III | 2006–2011 | 2012–2017 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 km | 2 km | 3 km | 4 km | 5 km | 1 km | 2 km | 3 km | 4 km | 5 km | |
| 112—Discontinuous urban fabric | −0.043 | −0.057 | −0.022 | −0.02 | −0.019 | 0.017 | −0.139 | −0.180 * | −0.273 ** | −0.304 *** |
| 121—Industrial or commercial units | 0.051 | 0.018 | 0.019 | 0.02 | −0.046 | −0.251 ** | −0.336 *** | −0.339 *** | −0.282 ** | −0.308 *** |
| 122—Road and rail networks and associated land | −0.038 | −0.039 | −0.231 * | −0.235 ** | ||||||
| 123—Port areas | −0.126 | −0.088 | 0.032 | 0.033 | ||||||
| 124—Airports | 0.027 | −0.026 | −0.104 | −0.240 ** | −0.328 *** | −0.314 *** | ||||
| 131—Mineral extraction sites | 0.060 | −0.028 | −0.088 | −0.099 | −0.080 | −0.028 | −0.015 | −0.003 | −0.013 | −0.002 |
| 132—Dump sites | 0.060 | 0.060 | −0.024 | −0.129 | −0.129 | −0.252 ** | ||||
| 133—Construction sites | 0.060 | −0.010 | −0.055 | −0.057 | −0.086 | |||||
| 141—Green urban areas | 0.057 | 0.013 | 0.045 | −0.124 | −0.264 ** | −0.262 ** | ||||
| 142—Sport and leisure facilities | −0.184 * | −0.035 | 0.003 | −0.006 | −0.047 | −0.109 | −0.229 * | −0.190 * | −0.257 ** | −0.250 ** |
| 211—Non-irrigated arable land | 0.097 | 0.104 | 0.100 | 0.094 | 0.112 | 0.245 ** | 0.235 ** | 0.159 ᵗʳ | 0.105 | 0.074 |
| 222—Fruit trees and berry plantations | 0.052 | 0.019 | 0.016 | 0.039 | 0.047 | −0.022 | −0.007 | 0.001 | ||
| 231—Pastures | 0.038 | −0.065 | −0.078 | −0.029 | 0.022 | −0.075 | −0.055 | −0.044 | −0.034 | −0.021 |
| 242—Complex cultivation patterns | 0.048 | 0.027 | 0.022 | 0.016 | 0.052 | −0.041 | 0.057 | 0.089 | 0.073 | 0.057 |
| 243—Land principally occupied by agriculture | 0.013 | 0.084 | 0.092 | 0.142 | 0.157 ᵗʳ | 0.179 * | 0.180 * | 0.202 * | 0.225 * | 0.176 ᵗʳ |
| with significant areas of natural vegetation | −0.103 | −0.070 | −0.063 | −0.059 | −0.047 | −0.149 | −0.120 | −0.070 | −0.019 | 0.020 |
| 311—Broad-leaved forest | −0.152 ᵗʳ | −0.044 | −0.031 | −0.072 | −0.146 | −0.326 *** | −0.189 * | −0.113 | −0.054 | 0.003 |
| 312—Coniferous forest | 0.116 | −0.027 | −0.068 | −0.133 | −0.080 | −0.093 | −0.195 * | −0.16 ᵗʳ | −0.096 | 0.002 |
| 313—Mixed forest | −0.120 | −0.180 * | −0.181 * | −0.179 * | −0.217 * | −0.043 | −0.024 | −0.048 | −0.055 | −0.052 |
| 324—Transitional woodland/shrub | 0.060 | 0.081 | 0.101 | 0.127 | 0.074 | 0.045 | −0.052 | −0.037 | ||
| 411—Inland marshes | 0.078 | 0.106 | 0.087 | 0.082 | 0.074 | 0.026 | 0.064 | 0.108 | 0.110 | 0.107 |
| 511—Water courses | 0.079 | 0.110 | 0.095 | 0.127 | 0.131 | 0.027 | −0.074 | −0.165 ᵗʳ | −0.050 | −0.038 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bihałowicz, J.; Schwerk, A.; Dymitryszyn, I.; Olszewski, A.; Bihałowicz, J.S. The Impact of Land Cover on Nest Occupancy of the White Stork (Ciconia ciconia (L.)): A Case Study of Kampinos Forest, 2006–2018. Earth 2024, 5, 336-353. https://doi.org/10.3390/earth5030019
Bihałowicz J, Schwerk A, Dymitryszyn I, Olszewski A, Bihałowicz JS. The Impact of Land Cover on Nest Occupancy of the White Stork (Ciconia ciconia (L.)): A Case Study of Kampinos Forest, 2006–2018. Earth. 2024; 5(3):336-353. https://doi.org/10.3390/earth5030019
Chicago/Turabian StyleBihałowicz, Joanna, Axel Schwerk, Izabela Dymitryszyn, Adam Olszewski, and Jan Stefan Bihałowicz. 2024. "The Impact of Land Cover on Nest Occupancy of the White Stork (Ciconia ciconia (L.)): A Case Study of Kampinos Forest, 2006–2018" Earth 5, no. 3: 336-353. https://doi.org/10.3390/earth5030019
APA StyleBihałowicz, J., Schwerk, A., Dymitryszyn, I., Olszewski, A., & Bihałowicz, J. S. (2024). The Impact of Land Cover on Nest Occupancy of the White Stork (Ciconia ciconia (L.)): A Case Study of Kampinos Forest, 2006–2018. Earth, 5(3), 336-353. https://doi.org/10.3390/earth5030019