
Impact of Disturbances on Species Diversity and Regeneration of Nepalese Sal (Shorea robusta) Forests Managed under Different Management Regimes


Abstract
:1. Introduction
2. Materials and Methods
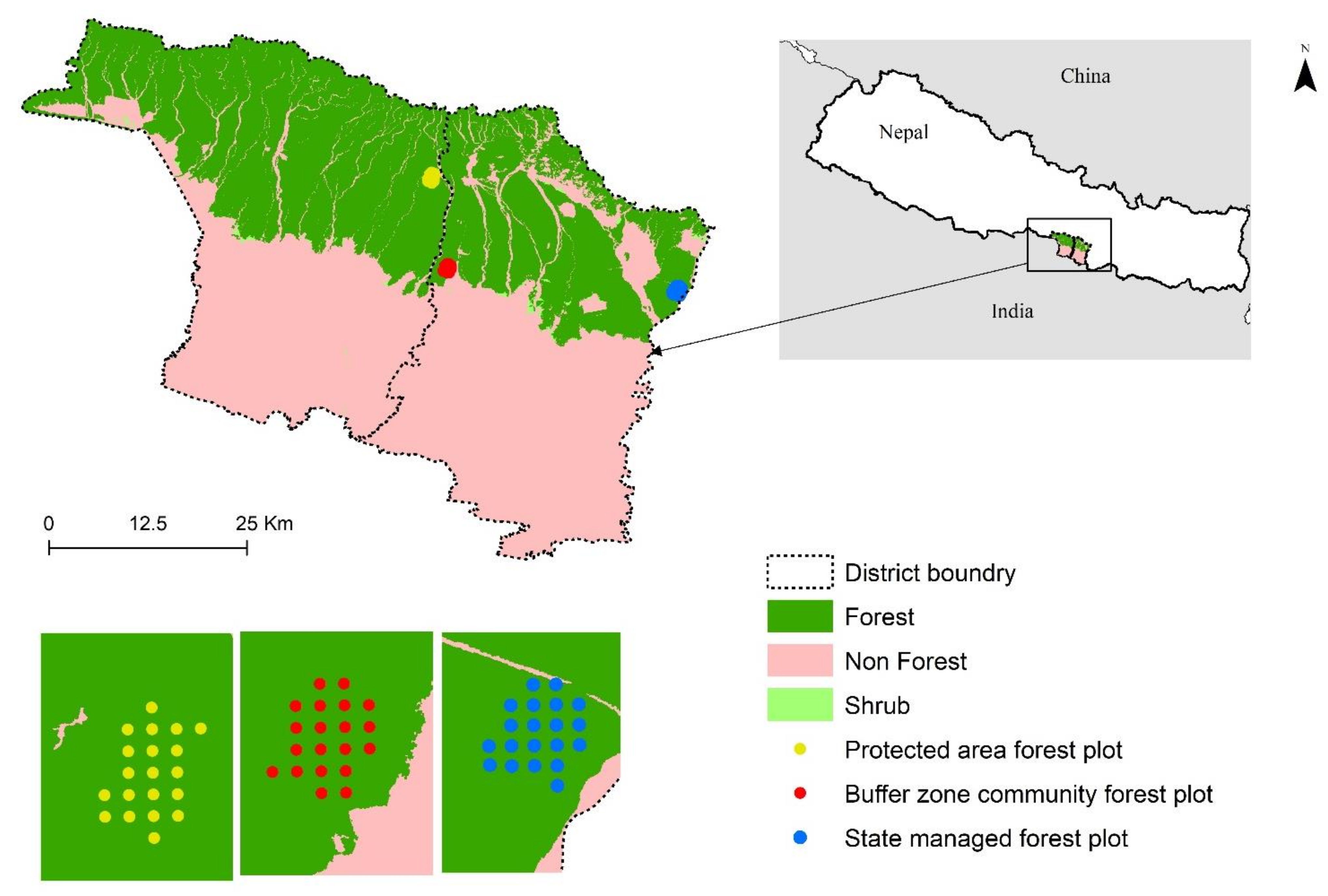
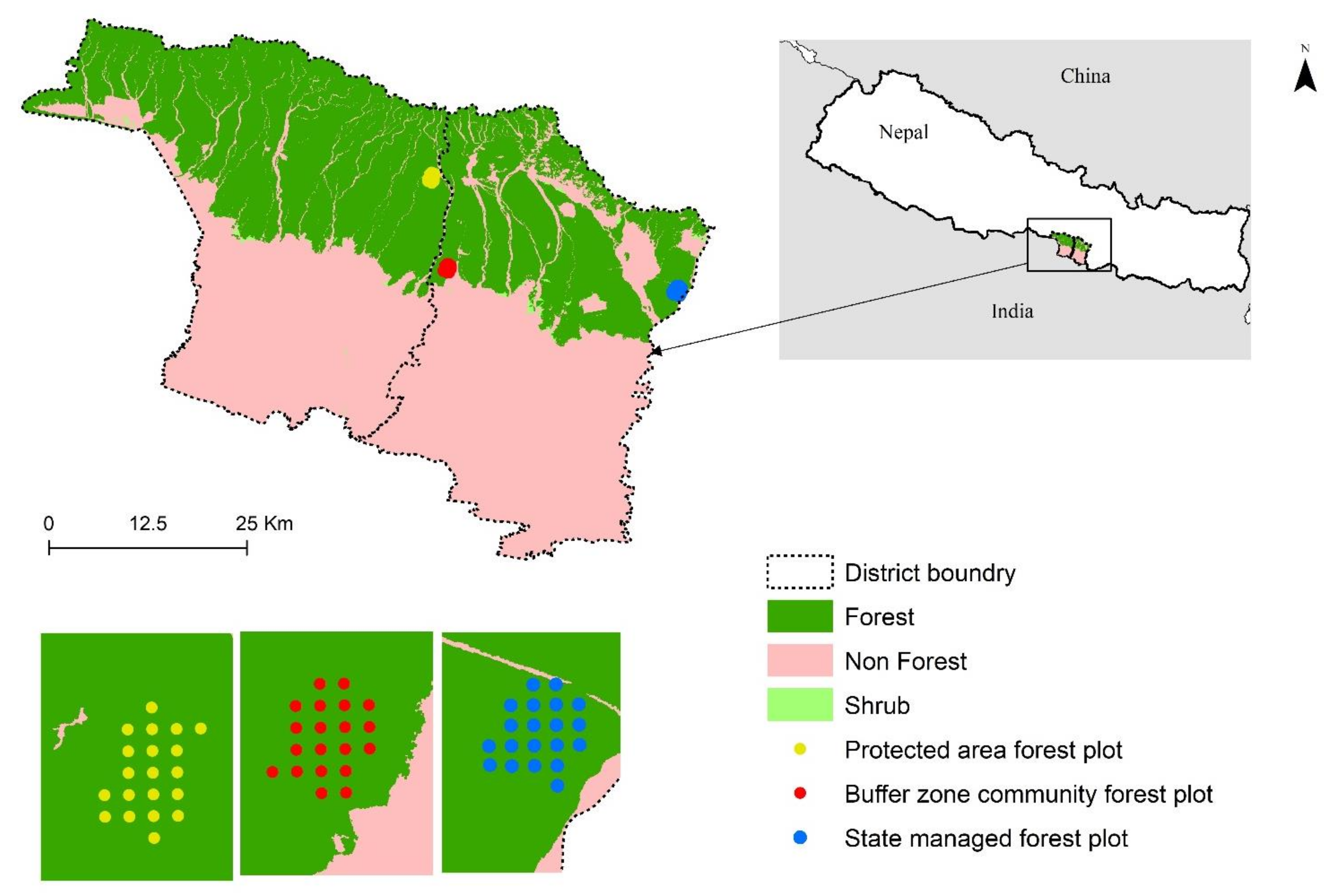
2.1. Study Area
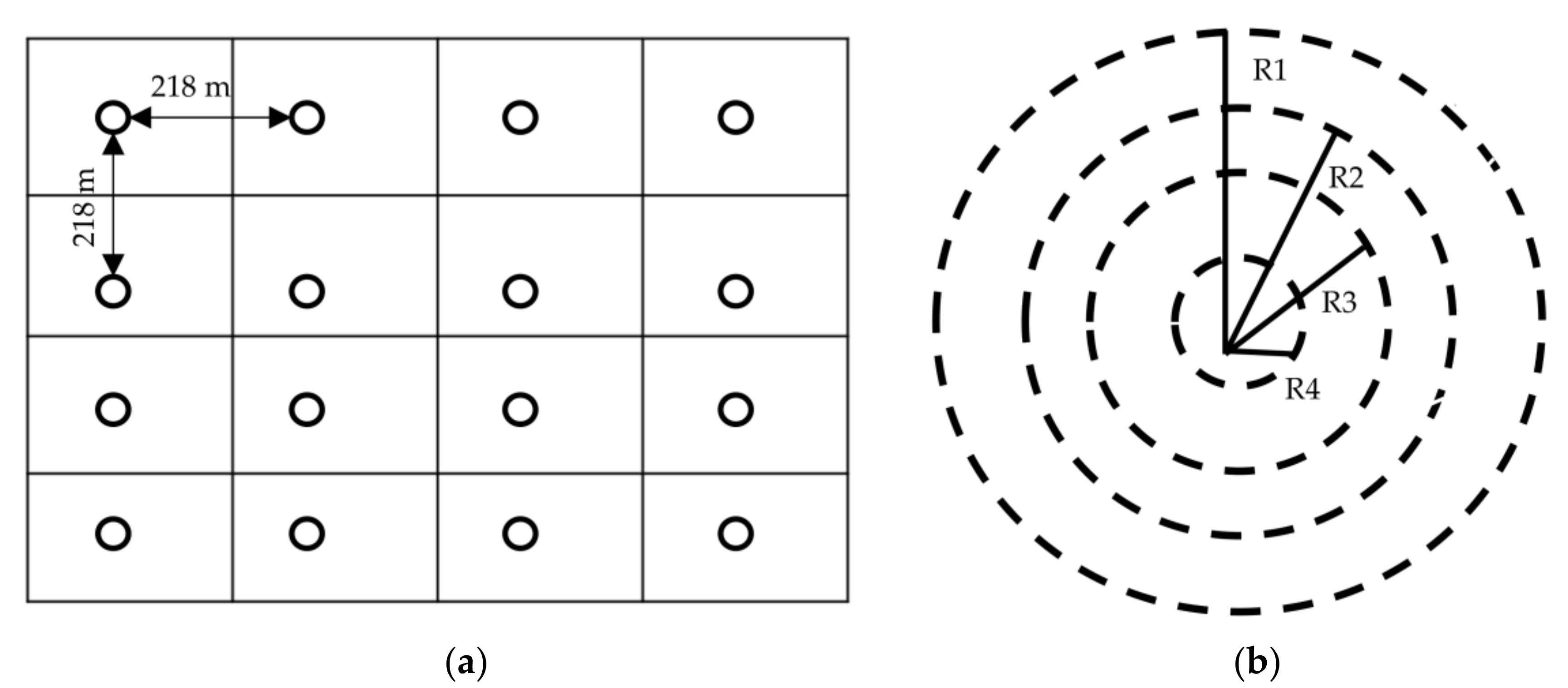
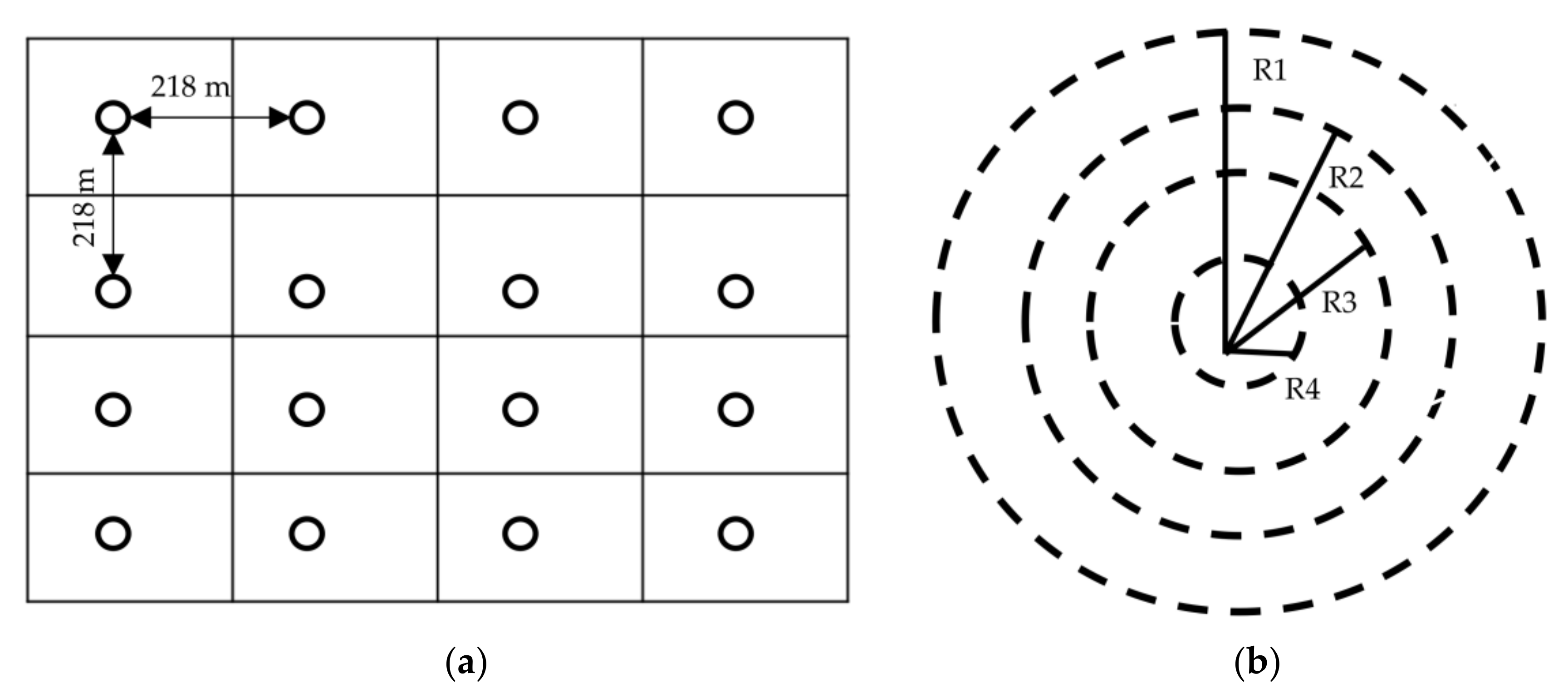
2.2. Sampling and Inventory
2.3. Assessment of Disturbances
2.4. Data Analysis
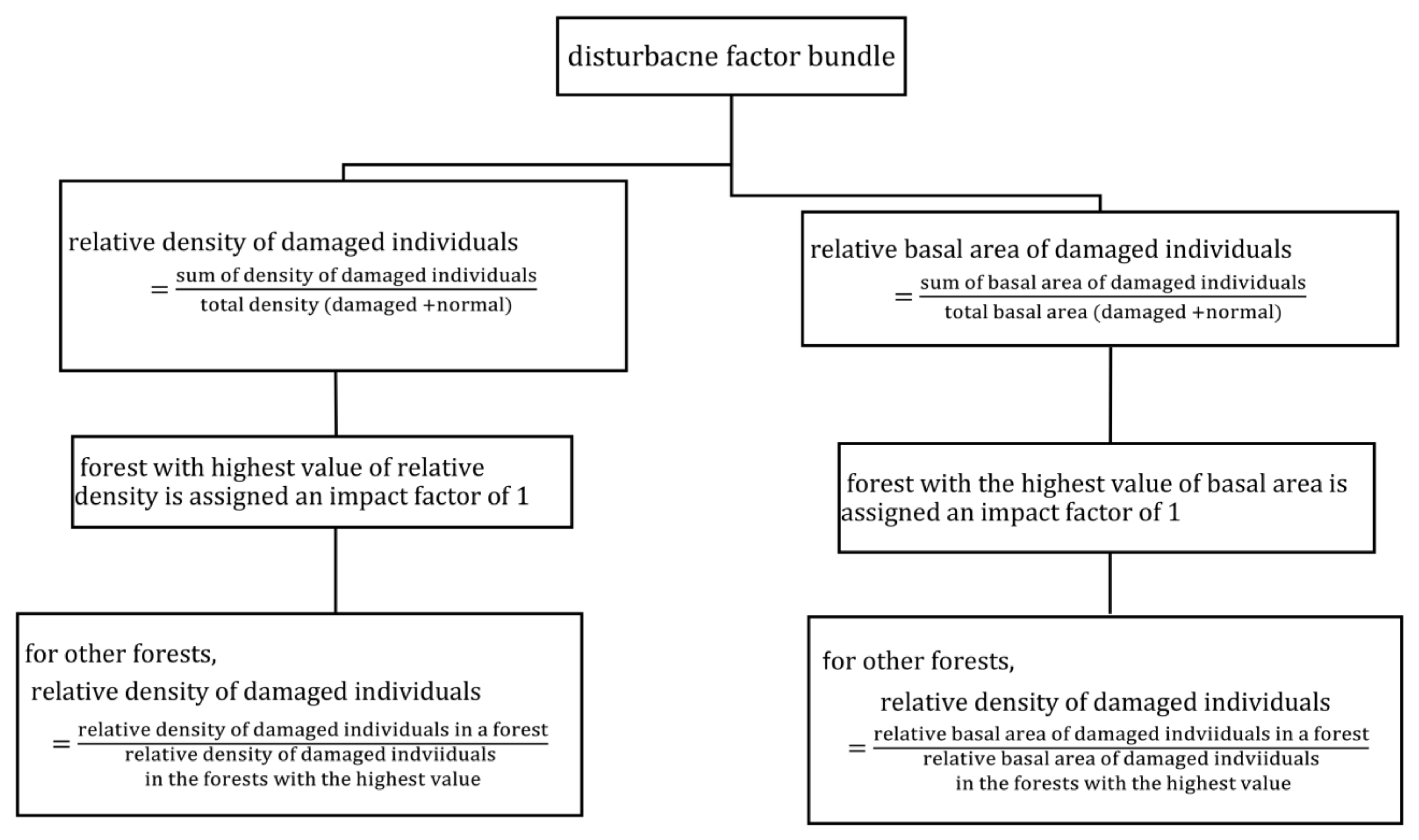
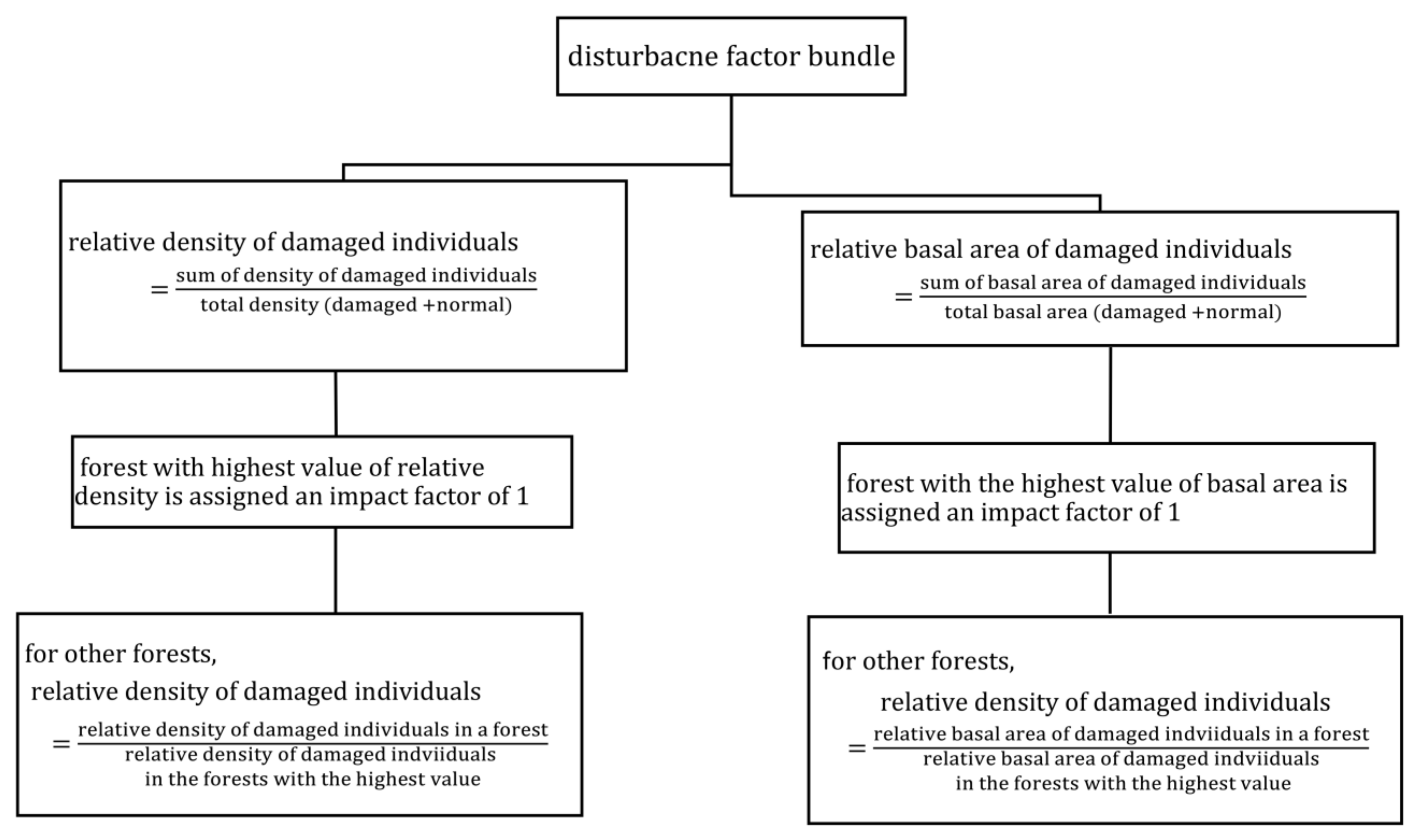
2.4.1. Disturbance Factor Estimation
2.4.2. Species Richness
2.4.3. Biodiversity Indices
2.4.4. Species Dominance
3. Results
3.1. Physical and Chemical Properties of Soils
3.2. Disturbance Factor (DF)
3.3. Species Richness and Diversity
3.4. Stand Structure
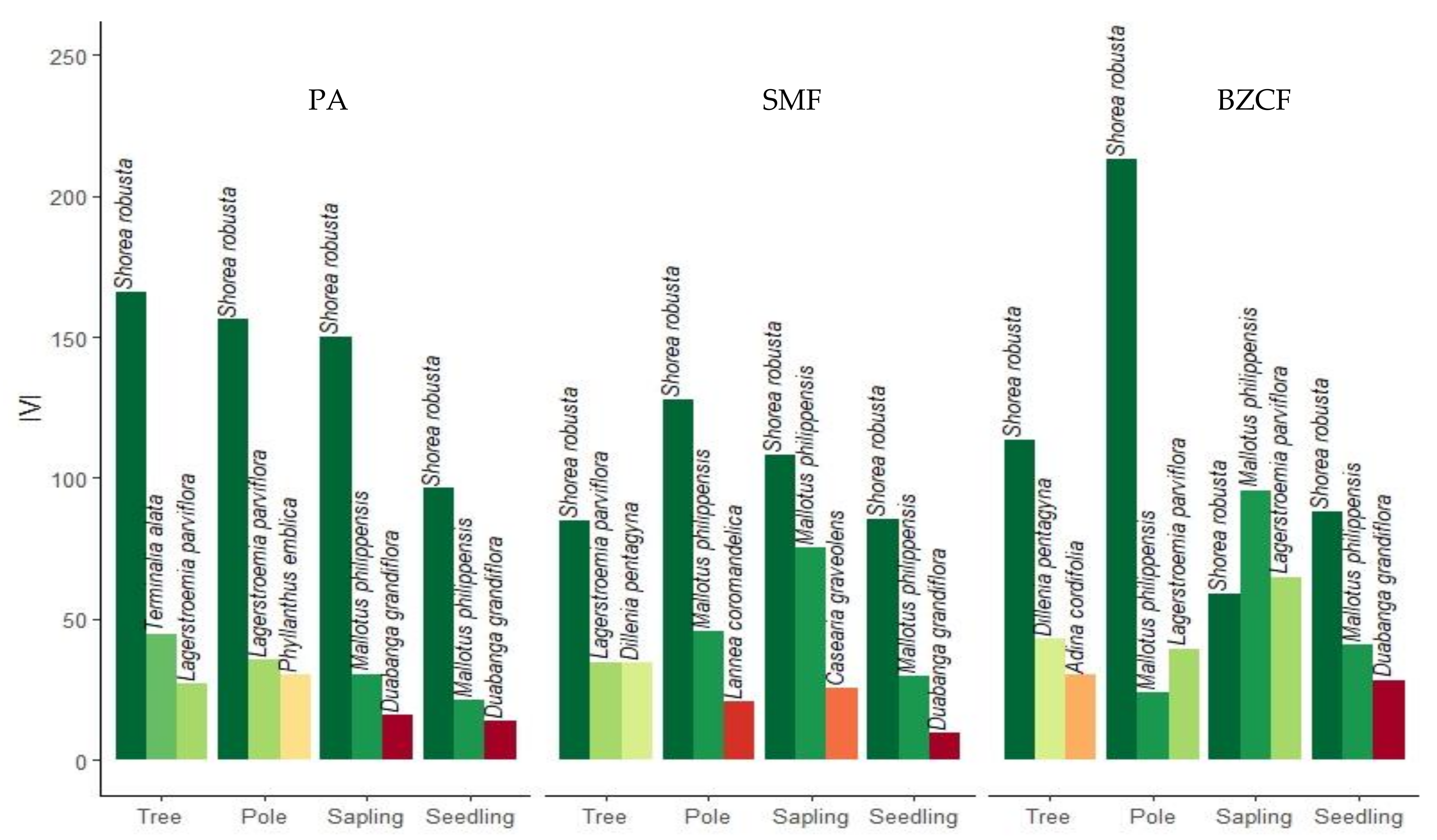
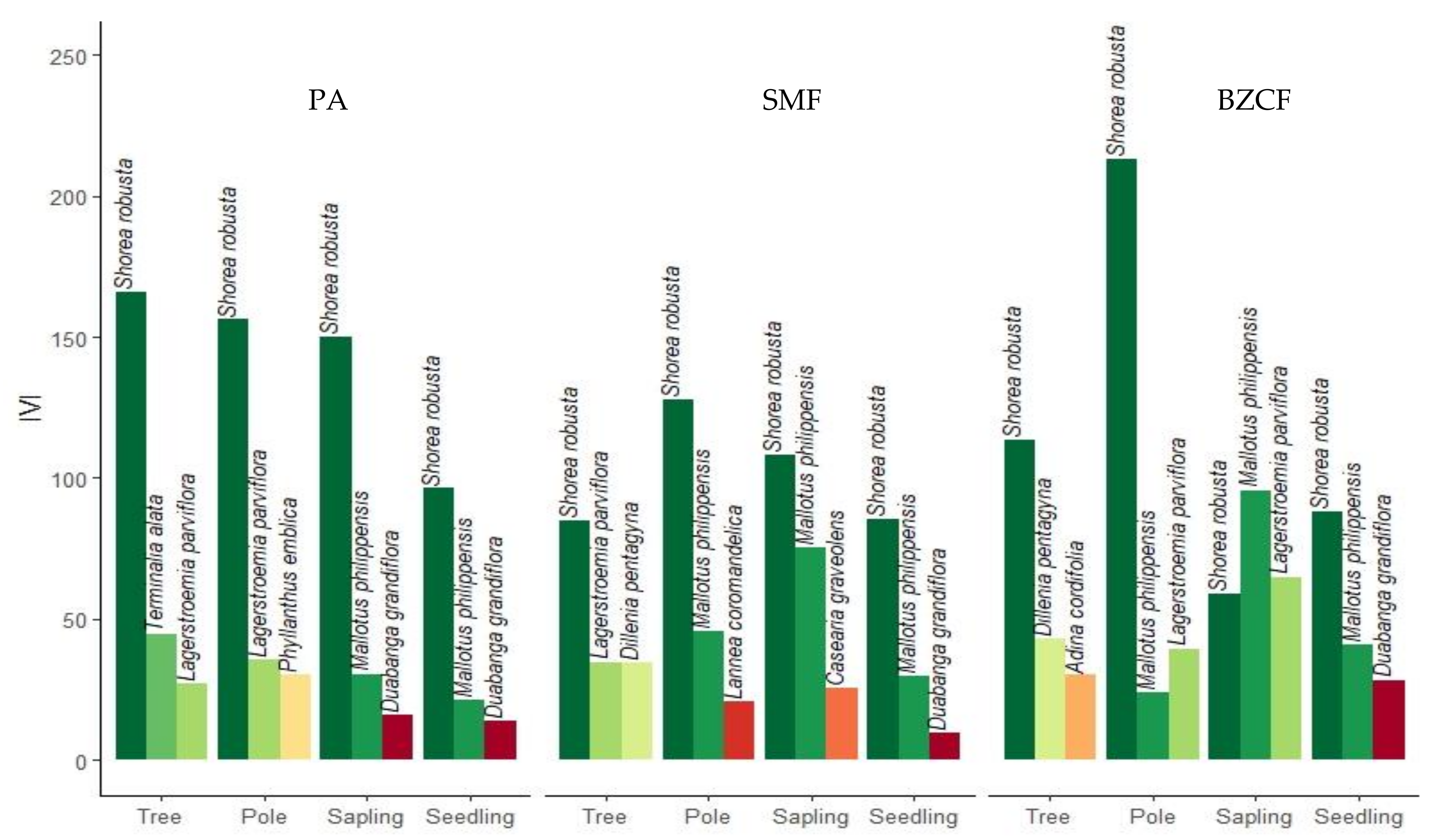
3.5. Dominance of the Species
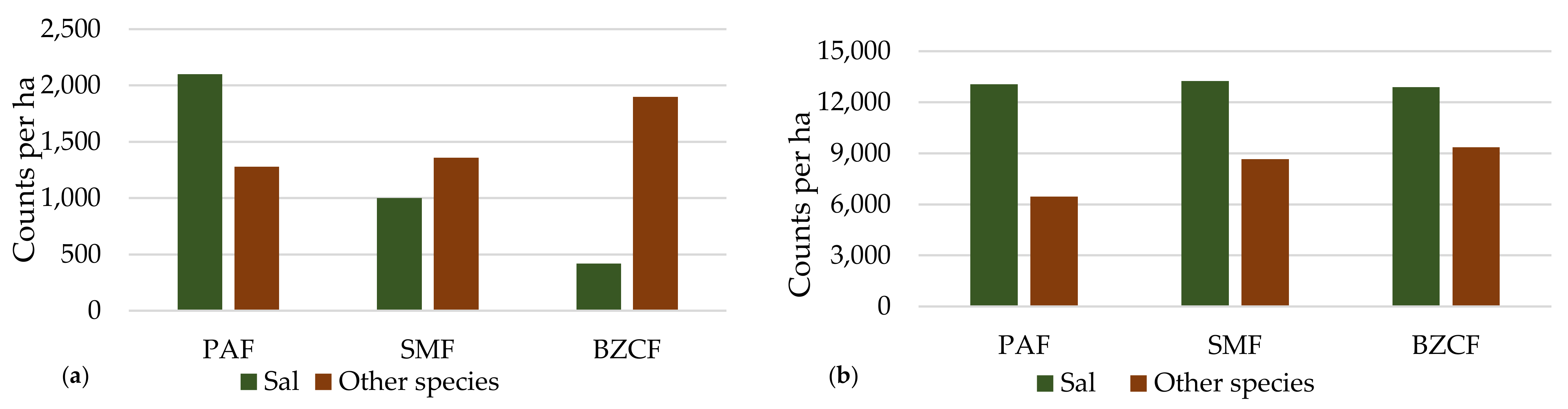
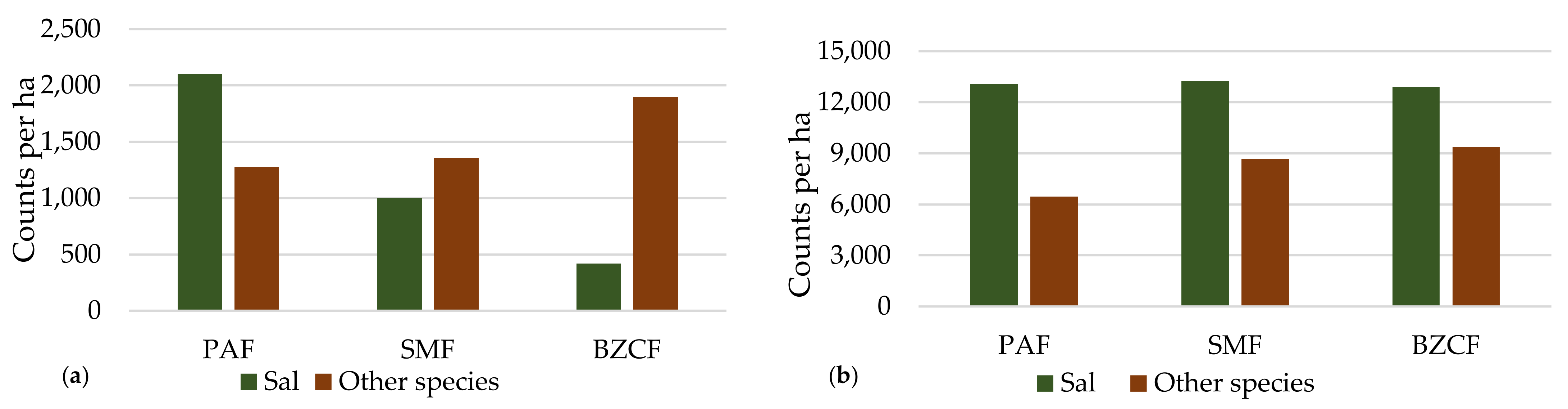
3.6. Status of Regeneration
3.7. Disturbance Bundles
4. Discussion
4.1. Soil Properties
4.2. Species Richness, Diversity, and Stand Structure
4.3. Dominance of the Species
4.4. Status of Regeneration
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sagar, R.; Raghubanshi, A.S.; Singh, J.S. Tree Species Composition, Dispersion and Diversity along a Disturbance Gradient in a Dry Tropical Forest Region of India. For. Ecol. Manag. 2003, 186, 61–71. [Google Scholar] [CrossRef]
- White, P.S.; Jentsch, A. The Search for Generality in Studies of Disturbance and Ecosystem Dynamics. In Progress in Botany; Springer: Berlin/Heidelberg, Germany, 2001; pp. 399–450. [Google Scholar]
- Siebert, S.F.; Belsky, J.M. Historic Livelihoods and Land Uses as Ecological Disturbances and Their Role in Enhancing Biodiversity: An Example from Bhutan. Biol. Conserv. 2014, 177, 82–89. [Google Scholar] [CrossRef]
- Gautam, M.K.; Manhas, R.K.; Tripathi, A.K. Patterns of Diversity and Regeneration in Unmanaged Moist Deciduous Forests in Response to Disturbance in Shiwalik Himalayas, India. J. Asia Pac. Biodivers. 2016, 9, 144–151. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Mao, Z.; Hu, L.; Zhang, J. Plant Diversity of Secondary Forests in Response to Anthropogenic Disturbance Levels in Montane Regions of Northeastern China. J. For. Res. 2007, 12, 403–416. [Google Scholar] [CrossRef]
- Sapkota, R.P.; Stahl, P.D.; Norton, U. Anthropogenic Disturbances Shift Diameter Distribution of Woody Plant Species in Shorea Robusta Gaertn. (Sal) Mixed Forests of Nepal. J. Asia Pac. Biodivers. 2019, 12, 115–128. [Google Scholar] [CrossRef]
- Ramírez-Marcial, N.; González-Espinosa, M.; Williams-Linera, G. Anthropogenic Disturbance and Tree Diversity in Montane Rain Forests in Chiapas, Mexico. For. Ecol. Manag. 2001, 154, 311–326. [Google Scholar] [CrossRef]
- Kennard, D.K.; Gould, K.; Putz, F.E.; Fredericksen, T.S.; Morales, F. Effect of Disturbance Intensity on Regeneration Mechanisms in a Tropical Dry Forest. For. Ecol. Manag. 2002, 162, 197–208. [Google Scholar] [CrossRef] [Green Version]
- Franklin, J.F. Ecological Characteristics of Old-Growth Douglas-Fir Forests; United States Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Statation: Corvallis, Oregon, USA, 1981; Volume 118. [Google Scholar]
- Bennett, L.T.; Adams, M.A. Assessment of Ecological Effects Due to Forest Harvesting: Approaches and Statistical Issues. J. Appl. Ecol. 2004, 41, 585–598. [Google Scholar] [CrossRef]
- Elderd, B.D.; Doak, D.F. Comparing the Direct and Community-Mediated Effects of Disturbance on Plant Population Dynamics: Flooding, Herbivory and Mimulus Guttatus. J. Ecol. 2006, 94, 656–669. [Google Scholar] [CrossRef]
- Khumbongmayum, A.D.; Khan, M.L.; Tripathi, R.S. Biodiversity Conservation in Sacred Groves of Manipur, Northeast India: Population Structure and Regeneration Status of Woody Species. In Human Exploitation and Biodiversity Conservation; Springer: Berlin/Heidelberg, Germany, 2006; pp. 99–116. [Google Scholar]
- Sapkota, I.P.; Tigabu, M.; Odén, P.C. Changes in Tree Species Diversity and Dominance across a Disturbance Gradient in Nepalese Sal (Shorea Robusta Gaertn. f.) Forests. J. For. Res. 2010, 21, 25–32. [Google Scholar] [CrossRef]
- Nagaike, T.; Kamitani, T.; Nakashizuka, T. The Effect of Shelterwood Logging on the Diversity of Plant Species in a Beech (Fagus Crenata) Forest in Japan. For. Ecol. Manag. 1999, 118, 161–171. [Google Scholar] [CrossRef]
- Sapkota, I.P.; Tigabu, M.; Odén, P.C. Spatial Distribution, Advanced Regeneration and Stand Structure of Nepalese Sal (Shorea Robusta) Forests Subject to Disturbances of Different Intensities. For. Ecol. Manag. 2009, 257, 1966–1975. [Google Scholar] [CrossRef]
- Sheil, D.; Burslem, D.F.R.P. Disturbing Hypotheses in Tropical Forests. Trends Ecol. Evol. 2003, 18, 18–26. [Google Scholar] [CrossRef]
- Connell, J.H. Diversity in Tropical Rain Forests and Coral Reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molino, J.; Sabatier, D. Tree Diversity in Tropical Rain Forests: A Validation of the Intermediate Disturbance Hypothesis. Science 2001, 294, 1702–1704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandel, P.; Chapagain, P.S.; Sharma, L.N.; Vetaas, O.R. Consumption Patterns of Fuelwood in Rural Households of Dolakha District, Nepal: Reflections from Community Forest User Groups. Small Scale For. 2016, 15, 481–495. [Google Scholar] [CrossRef]
- Hardin, G. The Tragedy of the Commons. J. Nat. Resour. Policy Res. 2009, 1, 243–253. [Google Scholar] [CrossRef]
- Forest Resource Assessment (FRA) Nepal, Department of Forest Research and Survey (DFRS). State of Nepal’s Forests; Department of Forest Research and Survey: Kathmandu, Nepal, 2015. [Google Scholar]
- Joshi, A.R. Establishing Biological Monitoring System for Terai Arc Landscape; A report submitted to World Wildlife Fund (WWF) Nepal Program; WWF Nepal: Kathmandu, Nepal, 2002. [Google Scholar]
- Timilsina, N.; Ross, M.S.; Heinen, J.T. A Community Analysis of Sal (Shorea Robusta) Forests in the Western Terai of Nepal. For. Ecol. Manag. 2007, 241, 223–234. [Google Scholar] [CrossRef]
- DFRS/FRA. Terai Forests of Nepal; DFRS/FRA: Kathmandu, Nepal, 2014. [Google Scholar]
- Ghimire, B.R.; Nagai, M.; Tripathi, N.K.; Witayangkurn, A.; Mishara, B.; Sasaki, N. Mapping of Shorea Robusta Forest Using Time Series MODIS Data. Forests 2017, 8, 384. [Google Scholar] [CrossRef]
- Lalfakawma; Sahoo, U.K.; Roy, S.; Vanlalhriatpuia, K.; Vanalalhluna, P.C. Community Composition and Tree Population Semi-Evergreen Forest Stands of North-East India. Appl. Ecol. Environ. Res. 2009, 7, 303–318. [Google Scholar] [CrossRef]
- Dinerstein, E. An Ecological Survey of the Royal Karnali-Bardia Wildlife Reserve, Nepal. Part I: Vegetation, Modifying Factors, and Successional Relationships. Biol. Conserv. 1979, 15, 127–150. [Google Scholar] [CrossRef]
- Webb, E.L.; Sah, R.N. Structure and Diversity of Natural and Managed Sal (Shorea Robusta Gaertn.f.) Forest in the Terai of Nepal. For. Ecol. Manag. 2003, 176, 337–353. [Google Scholar] [CrossRef]
- Acharya, H.; Ram, A.; Shtestha, S. Parsa National Park and Its Buffer Zone Management Plan (2075/76-2079/80); Parsa National Park: Aadhavar, Bara, Nepal, 2018. [Google Scholar]
- Radhakrishna Buffer Zone Management Plan. Management Plan of Radhakrishna Bufferzone Community Forest; Radhakrishna Bufffer Zone Management Committee: Simara, Bara, Nepal, 2018. [Google Scholar]
- Poudyal, B.H.; Maraseni, T.; Cockfield, G. Impacts of Forest Management on Tree Species Richness and Composition: Assessment of Forest Management Regimes in Tarai Landscape Nepal. Appl. Geogr. 2019, 111, 102078. [Google Scholar] [CrossRef]
- Bhuyan, P.; Khan, M.L.; Tripathi, R.S. Tree Diversity and Population Structure in Undisturbed and Human-Impacted Stands of Tropical Wet Evergreen Forest in Arunachal Pradesh, Eastern Himalayas, India. Biodivers. Conserv. 2003, 12, 1753–1773. [Google Scholar] [CrossRef]
- Thapa, S.; Chapman, D.S. Impacts of Resource Extraction on Forest Structure and Diversity in Bardia National Park, Nepal. For. Ecol. Manag. 2010, 259, 641–649. [Google Scholar] [CrossRef]
- Sahu, P.K.; Sagar, R.; Singh, J.S. Tropical Forest Structure and Diversity in Relation to Altitude and Disturbance in a Biosphere Reserve in Central India. Appl. Veg. Sci. 2008, 11, 461–470. [Google Scholar] [CrossRef]
- Mishra, B.P.; Tripathi, O.P.; Tripathi, R.S.; Pandey, H.N. Effects of Anthropogenic Disturbance on Plant Diversity and Community Structure of a Sacred Grove in Meghalaya, Northeast India. Biodivers. Conserv. 2004, 13, 421–436. [Google Scholar] [CrossRef]
- Htun, N.Z.; Mizoue, N.; Yoshida, S. Tree Species Composition and Diversity at Different Levels of Disturbance in Popa Mountain Park, Myanmar. Biotropica 2011, 43, 597–603. [Google Scholar] [CrossRef]
- Jaccard, P. The Distribution of the Flora in the Alpine Zone. New Phytol. 1912, 11, 37–50. [Google Scholar] [CrossRef]
- Magurran, A.E. Ecological Diversity and Its Measurement; Princeton University Press: Princeton, NJ, USA, 1988. [Google Scholar]
- Simpson, E.H. Measurment of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Pielou, E.C. Species-Diversity and Pattern-Diversity in the Study of Ecological Succession. J. Theor. Biol. 1966, 10, 370–383. [Google Scholar] [CrossRef]
- Misra, R. Ecology Workbook; Oxford & IBH Publishing Company: New Delhi, India, 1968. [Google Scholar]
- Curtis, J.T.; McIntosh, R.P. An Upland Forest Continuum in the Prairie-Forest Border Region of Wisconsin. Ecology 1951, 32, 476–496. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. R Package Vegan version 2.2-1. 2018, pp. 2–5. Available online: https://www.worldagroforestry.org/publication/vegan-community-ecology-package-r-package-vegan-vers-22-1 (accessed on 10 October 2020).
- Kindt, R.; Coe, R. Tree Diversity Analysis: A Manual and Software for Common Statistical Methods for Ecological and Biodiversity Studies; World Agroforestry Centre: Nairobi, Kenya, 2005. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2013. [Google Scholar]
- Parthasarathy, N.; Karthikeyan, R. Plant Biodiversity Inventory and Conservation of Two Tropical Dry Evergreen Forests on the Coromandel Coast, South India. Biodivers. Conserv. 1997, 6, 1063–1083. [Google Scholar] [CrossRef]
- Nurse, M.; Malla, Y. Advances in Community Forestry in Asia. Capital. Shar. Exp. Interact. For. Policies L. Use Patterns Asia 2006, 2, 25–31. [Google Scholar]
- Gilmour, D.; Malla, Y.; Nurse, M. Linkages between Community Forestry and Poverty. Bangkok: Regional Community Forestry Center for Asia and the Pacific. 2004. Available online: http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.514.3609&rep=rep1&type=pdf (accessed on 10 October 2020).
- Shrestha, U.B.; Shrestha, S.; Chaudhary, P.; Chaudhary, R.P. How Representative Is the Protected Areas System of Nepal? Mt. Res. Dev. 2010, 30, 282–294. [Google Scholar] [CrossRef]
- Paudel, S.; Sah, J.P. Physiochemical Characteristics of Soil in Tropical Sal (Shorea Robusta Gaertn.) Forests in Eastern Nepal. Himal. J. Sci. 2003, 1, 107–110. [Google Scholar] [CrossRef] [Green Version]
- Bhatnagar, H.P. Soils from Different Quality Sal (Shorea Robusta) Forests of Uttar Pradesh. Trop. Ecol. 1965, 6, 56–62. [Google Scholar]
- Jackson, J.K. Manual of Afforestation in Nepal: Forest Research and Survey Center; Forest Survey and Research Office, Department of Forest: Kathmandu, Nepal, 1994. [Google Scholar]
- Reinhardt, J.R. Community-Level Impacts of Management and Disturbance in Western Michigan Oak Savannas. Am. Midl. Nat. 2017, 177, 112–125. [Google Scholar] [CrossRef]
- McKinney, M.L. Urbanization, Biodiversity, and ConservationThe Impacts of Urbanization on Native Species Are Poorly Studied, but Educating a Highly Urbanized Human Population about These Impacts Can Greatly Improve Species Conservation in All Ecosystems. Bioscience 2002, 52, 883–890. [Google Scholar] [CrossRef]
- Rahman, M.D.M.; Nishat, A.; Vacik, H. Anthropogenic Disturbances and Plant Diversity of the Madhupur Sal Forests (Shorea Robusta, C.F. Gaertn) of Bangladesh. Int. J. Biodivers. Sci. Manag. 2009, 5, 162–173. [Google Scholar] [CrossRef]
- Pandey, S.K.; Shukla, R.P. Plant Diversity in Managed Sal (Shorea Robusta Gaertn.) Forests of Gorakhpur, India: Species Composition, Regeneration and Conservation. Biodivers. Conserv. 2003, 12, 2295–2319. [Google Scholar] [CrossRef]
- Rabha, D. Species Composition and Structure of Sal (Shorea Robusta Gaertn. f.) Forests along Disturbance Gradients of Western Assam, Northeast India. Trop. Plant Res. 2014, 1, 16–21. [Google Scholar]
- Onaindia, M.; Dominguez, I.; Albizu, I.; Garbisu, C.; Amezaga, I. Vegetation Diversity and Vertical Structure as Indicators of Forest Disturbance. For. Ecol. Manag. 2004, 195, 341–354. [Google Scholar] [CrossRef]
- Peltzer, D.A.; Bast, M.L.; Wilson, S.D.; Gerry, A.K. Plant Diversity and Tree Responses Following Contrasting Disturbances in Boreal Forest. For. Ecol. Manag. 2000, 127, 191–203. [Google Scholar] [CrossRef]
- Rao, P.; Barik, S.K.; Pandey, H.N.; Tripathi, R.S. Community Composition and Tree Population Structure in a Sub-Tropical Broad-Leaved Forest along a Disturbance Gradient. Vegetatio 1990, 88, 151–162. [Google Scholar] [CrossRef]
- Banda, T.; Schwartz, M.W.; Caro, T. Woody Vegetation Structure and Composition along a Protection Gradient in a Miombo Ecosystem of Western Tanzania. For. Ecol. Manag. 2006, 230, 179–185. [Google Scholar] [CrossRef]
- Connell, J.H.; Lowman, M.D. Low-Diversity Tropical Rain Forests: Some Possible Mechanisms for Their Existence. Am. Nat. 1989, 134, 88–119. [Google Scholar] [CrossRef]
- Gautam, K.H.; Devoe, N.N. Ecological and Anthropogenic Niches of Sal (Shorea Robusta Gaertn. f.) Forest and Prospects for Multiple-Product Forest Management—A Review. Forestry 2006, 79, 81–101. [Google Scholar] [CrossRef]
- Mandal, G.; Joshi, S.P. Analysis of Vegetation Dynamics and Phytodiversity from Three Dry Deciduous Forests of Doon Valley, Western Himalaya, India. J. Asia Pac. Biodivers. 2014, 7, 292–304. [Google Scholar] [CrossRef] [Green Version]
- Keya, G.A. Herbaceous Layer Production and Utilization by Herbivores under Different Ecological Conditions in an Arid Savanna of Kenya. Agric. Ecosyst. Environ. 1998, 69, 55–67. [Google Scholar] [CrossRef]
- Beguin, J.; Pothier, D.; Prévost, M. Can the Impact of Deer Browsing on Tree Regeneration Be Mitigated by Shelterwood Cutting and Strip Clearcutting? For. Ecol. Manag. 2009, 257, 38–45. [Google Scholar] [CrossRef]
- Gautam, T.P.; Mandal, T.N. Effect of Disturbance on Plant Species Diversity in Moist Tropical Forest of Eastern Nepal. Our Nat. 2018, 16, 1–7. [Google Scholar] [CrossRef]
- Belbase, K.; Poudel, A.S.; Chhetri, S.G. Comparison of forest stand condition between thinned and unthinned blocks under collaborative forest management of western lowlands of Nepal. 2020; Preprint available at Research Square. [Google Scholar] [CrossRef]
- Bhatta, B.P.; Chhetri, S.G.; Silwal, T.; Poudel, M. Economic contribution of forestry sector to national economy in Nepal. J. Res. Eco. 2021, 12, 620–627. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Management Regime | ||
|---|---|---|---|
| PAF | SMF | BZCF | |
| Objective | Protection of range and endangered wild animals and plants and their habitat, promotion of biodiversity and ecotourism | Supply of forest products including timber and fuelwood at the national level | Fulfill the forest products’ needs of local communities and maintain biodiversity |
| Rights to govern on forest use and conservation | Park administration | District and local level forest officials under the Department of Forest | BZCF users and protected area authority collectively make the decision |
| Rights to sell forest products | Prohibited use of most of the forest products | Government of Nepal | There is restriction on sale of forest products outside the user group except to sell non-timber forest products |
| Rights to manage forest | Officials under the Department of National Parks and Wildlife Conservation and Nepal Army | District and local level forest officials under the Department of Forests | BZCF users group manage and conserve the forest, but the park officials also have authority to involve in conservation |
| Local communities’ participation in forest management and conservation | Low | Low | High |
| Timber harvesting | Almost nil except for habitat management works and construction of roads and firelines | Planned forest harvesting is minimal despite having a forest management plan | Regular silvicultural operation and harvesting as provision in forest management plan |
| Frequency and extend of forest harvesting | Close to nil | Minimal | Depending on the resource condition and user group dynamics |
| Chemical Properties | Management Regime | ||
|---|---|---|---|
| BZC | SMF | PAF | |
| pH | 7.43 ± 0.12 a | 7.57 ± 0.15 a | 7.64 ± 0.13 a |
| N (%) | 0.131 ± 0.0052 a | 0.131 ± 0.0069 a | 0.148 ± 0.070 a |
| P (Kg/Ha) | 37.92 ± 2.2 a | 31.83 ± 2.69 a | 56.17 ± 3.74 b |
| K (Kg/Ha) | 176.94 ± 4 a | 181.71 ± 3.41 a | 240.49 ± 7.53 b |
| Disturbance Index | Management Regime | ||
|---|---|---|---|
| BZCF | SMF | PAF | |
| Disturbed seedling frequency | 1.00 | 0.43 | 0.07 |
| Disturbed saplings frequency | 1.00 | 0.36 | 0.06 |
| Disturbed sapling BA | 0.29 | 1.00 | 0.16 |
| Lopping/Branch cutting frequency + | 1.00 | 0.14 | 0.00 |
| Lopping/Branch cutting BA + | 1.00 | 0.08 | 0.00 |
| Cutting/chopping/girdling/bark removal frequency + | 1.00 | 0.40 | 0.81 |
| Cutting/chopping/girdling/bark removal BA + | 1.00 | 0.53 | 0.83 |
| Pest/Climber frequency + | 1.00 | 0.81 | 0.40 |
| Pest/Climber BA + | 1.00 | 0.89 | 0.30 |
| Stagheaded frequency + | 0.00 | 1.00 | 0.44 |
| Stagheaded BA + | 0.00 | 1.00 | 0.37 |
| Crown open (%) | 1.00 | 0.92 | 0.95 |
| Distance from settlements (m) | 1.00 | 0.21 | 0.19 |
| Footpath area (m2) | 1.00 | 0.00 | 0.00 |
| Wild boar/bison wallowing/mouse mound (m2) | 1.00 | 0.66 | 0.82 |
| Grazing/Grass cutting (m2) | 1.00 | 0.00 | 0.00 |
| Stump felled (BA) | 0.86 | 1.00 | 0.00 |
| Stump Small frequency | 1.00 | 0.78 | 0.00 |
| W/L spots/termites (no.) | 0.17 | 0.17 | 1.00 |
| Wind-thrown frequency | 0.00 | 0.37 | 1.00 |
| Total | 15.32 | 10.75 | 7.40 |
| Species Composition | Management Regime | Jaccard Index | ||
|---|---|---|---|---|
| PAF | SMF | BZCF | ||
| Total Species | 30 | 33 | 16 | - |
| No. of unique species | 4 | 5 | 1 | - |
| No. of species with single individual | 6 | 5 | 1 | - |
| Common species in all forest types | 13 | - | ||
| Overlapping species between PAF and SMF | 26 | 0.70 | ||
| No. of overlapping species between PAF and BZCF | 13 | 0.39 | ||
| No. of overlapping species between SMF and BZCF | 15 | 0.44 | ||
| Management Regime | Size Class | Total Species | Mean Species Richness | Shannon-Weiner Index | Simpson’s Index | Evenness Index | Abundance |
|---|---|---|---|---|---|---|---|
| BZCF | Tree | 11 | 2.3 ± 0.26 a | 0.74 ± 0.11 a | 0.44 ± 0.06 a | 0.44 ± 0.05 a | 3 ± 0.36 a |
| SMF | 18 | 4 ± 0.40 b | 1.24 ± 0.10 b | 0.29 ± 0.02 a | 0.50 ± 0.032 a | 5.5 ± 0.58 b | |
| PAF | 13 | 2.6 ± 0.35 a | 0.65 ± 0.12 a | 0.63 ± 0.06 b | 0.35 ± 0.052 a | 5.9 ± 0.60 b | |
| BZCF | Pole | 6 | 1.6 ± 0.18 a | 0.33 ± 0.072 a | 0.74 ± 0.05 a | 0.28 ± 0.06 a | 4.9 ± 0.68 a |
| SMF | 14 | 2.05 ± 0.28 ab | 0.52 ± 0.11 ab | 0.63 ± 0.06 a | 0.33 ± 0.06 a | 4.45 ± 0.79 a | |
| PAF | 13 | 2.8 ± 0.39 b | 0.77 ± 0.15 b | 0.58 ± 0.07 a | 0.32 ± 0.05 a | 5.15 ± 0.54 a | |
| BZCF | Sapling | 10 | 2.45 ± 0.23 a | 0.71 ± 0.10 a | 0.58 ± 0.06 a | 0.41 ± 0.05 a | 5.8 ± 0.71 a |
| SMF | 12 | 2.45 ± 0.28 a | 0.73 ± 0.10 a | 0.46 ± 0.05 a | 0.44 ± 0.05 a | 5.9 ± 1.09 a | |
| PAF | 15 | 3.1 ± 0.33 a | 0.82 ± 0.11 a | 0.51 ± 0.06 a | 0.34 ± 0.03 a | 8.45 ± 0.73 a | |
| BZCF | Seedling | 8 | 2.8 ± 0.24 a | 0.76 ± 0.08 a | 0.55 ± 0.04 a | 0.41 ± 0.04 a | 22.25 ± 2.13 a |
| SMF | 20 | 3.85 ± 0.26 b | 0.95 ± 0.07 a | 0.48 ± 0.03 a | 0.41 ± 0.02 a | 21.9 ± 1.48 a | |
| PAF | 17 | 3.6 ± 0.24 ab | 0.88 ± 0.061 a | 0.51 ± 0.03 a | 0.41 ± 0.03 a | 19.5 ± 1.69 a |
| Stand Structure | Size Class at DBH (cm) | Disturbance Levels | ||
|---|---|---|---|---|
| BZCF | SMF | PAF | ||
| Density | >30 | 60 | 110 | 118 |
| 10–29.9 | 490 | 445 | 515 | |
| 1.3–4.9 | 2320 | 2360 | 3380 | |
| <1.3 | 22,250 | 21,900 | 19,500 | |
| Aggregate | 4013 ± 283.06 b | 2915 ± 440.39 ab | 2870 ± 270.84 a | |
| Tree basal area | >30 | 13.61 | 24.69 | 23.14 |
| 10–29.9 | 10.25 | 12.1 | 13.35 | |
| 1.3–4.9 | 2.13 | 2.26 | 2.98 | |
| Aggregate | 39.47 ± 1.78 b | 39.05 ± 3.20 b | 25.49 ± 2.03 a | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chapagain, U.; Chapagain, B.P.; Nepal, S.; Manthey, M. Impact of Disturbances on Species Diversity and Regeneration of Nepalese Sal (Shorea robusta) Forests Managed under Different Management Regimes. Earth 2021, 2, 826-844. https://doi.org/10.3390/earth2040049
Chapagain U, Chapagain BP, Nepal S, Manthey M. Impact of Disturbances on Species Diversity and Regeneration of Nepalese Sal (Shorea robusta) Forests Managed under Different Management Regimes. Earth. 2021; 2(4):826-844. https://doi.org/10.3390/earth2040049
Chicago/Turabian StyleChapagain, Uttam, Binod P. Chapagain, Sunil Nepal, and Michael Manthey. 2021. "Impact of Disturbances on Species Diversity and Regeneration of Nepalese Sal (Shorea robusta) Forests Managed under Different Management Regimes" Earth 2, no. 4: 826-844. https://doi.org/10.3390/earth2040049
APA StyleChapagain, U., Chapagain, B. P., Nepal, S., & Manthey, M. (2021). Impact of Disturbances on Species Diversity and Regeneration of Nepalese Sal (Shorea robusta) Forests Managed under Different Management Regimes. Earth, 2(4), 826-844. https://doi.org/10.3390/earth2040049