
In Silico Identification of Protein Targets Associated to the Insecticide Activity of Eugenol Derivatives †

 ,
, 
 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Methods
3. Results and Discussion
4. Conclusions
Acknowledgments
References
- Liu, X.; Cao, A.; Yan, D.; Ouyang, C.; Wang, Q.; Li, Y. Overview of mechanisms and uses of biopesticides. Int. J. Pest Manag. 2019, 67, 1–8. [Google Scholar] [CrossRef]
- Venthur, H.; Zhou, J. Odorant Receptors and Odorant-Binding Proteins as Insect Pest Control Targets: A Comparative Analysis. Front. Physiol. 2018, 9, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Osman, K.A. Pesticides and Human Health. In Pestic. Mod. World—Eff. Pestic. Expo; InTech: Hong Kong, 2011. [Google Scholar]
- Amoabeng, B.W.; Johnson, A.C.; Gurr, G.M. Natural enemy enhancement and botanical insecticide source: a review of dual use companion plants. Appl. Entomol. Zool. 2019, 54, 1–19. [Google Scholar] [CrossRef]
- Enan, E. Insecticidal activity of essential oils: octopaminergic sites of action. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2001, 130, 325–337. [Google Scholar] [CrossRef]
- Liska, A.; Rozman, V.; Kalinovic, I.; Ivezic, M.; Balicevir, R. Contact and fumigant activity of 1,8-cineole, eugenol and camphor against Tribolium castaneum (Herbst). Julius-Kühn-Archiv 2010, 425, 716. [Google Scholar]
- Wang, C.; Zhang, J.; Chen, H.; Fan, Y.; Shi, Z. Antifungal activity of eugenol against Botrytis cinerea. Trop. Plant Pathol. 2010, 35, 137–143. [Google Scholar] [CrossRef]
- Hu, X.; Yin, B.; Cappelle, K.; Swevers, L.; Smagghe, G.; Yang, X.; Zhang, L. Identification of novel agonists and antagonists of the ecdysone receptor by virtual screening. J. Mol. Graph. Model. 2018, 81, 77–85. [Google Scholar] [CrossRef]
- Harada, T.; Nakagawa, Y.; Ogura, T.; Yamada, Y.; Ohe, T.; Miyagawa, H. Virtual Screening for Ligands of the Insect Molting Hormone Receptor. J. Chem. Inf. Model. 2011, 51, 296–305. [Google Scholar] [CrossRef]
- Dong, Y.; Jiang, X.; Liu, T.; Ling, Y.; Yang, Q.; Zhang, L.; He, X. Structure-Based Virtual Screening, Compound Synthesis, and Bioassay for the Design of Chitinase Inhibitors. J. Agric. Food Chem. 2018, 66, 3351–3357. [Google Scholar] [CrossRef]
- Liu, J.; Liu, M.; Yao, Y.; Wang, J.; Li, Y.; Li, G.; Wang, Y. Identification of Novel Potential β-N-Acetyl-D-Hexosaminidase Inhibitors by Virtual Screening, Molecular Dynamics Simulation and MM-PBSA Calculations. Int. J. Mol. Sci. 2012, 13, 4545–4563. [Google Scholar] [CrossRef]
- Dong, L.; Shen, S.; Xu, Y.; Wang, L.; Yang, Q.; Zhang, J.; Lu, H. Identification of novel insect β-N-acetylhexosaminidase OfHex1 inhibitors based on virtual screening, biological evaluation, and molecular dynamics simulation. J. Biomol. Struct. Dyn. 2021, 39, 1735–1743. [Google Scholar] [CrossRef] [PubMed]
- Min, J.; Lin, D.; Zhang, Q.; Zhang, J.; Yu, Z. Structure-based virtual screening of novel inhibitors of the uridyltransferase activity of Xanthomonas oryzae pv. oryzae GlmU. Eur. J. Med. Chem. 2012, 53, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Ramos, R.; Costa, J.; Silva, R.; da Costa, G.; Rodrigues, A.; Rabelo, É.; Souto, R.; Taft, C.; Silva, C.; Rosa, J.; et al. Identification of Potential Inhibitors from Pyriproxyfen with Insecticidal Activity by Virtual Screening. Pharmaceuticals 2019, 12, 20. [Google Scholar] [CrossRef] [PubMed]
- Riva, C.; Suzanne, P.; Charpentier, G.; Dulin, F.; Halm-Lemeille, M.-P.; Sopkova-de Oliveira Santos, J. In silico chemical library screening and experimental validation of novel compounds with potential varroacide activities. Pestic. Biochem. Physiol. 2019, 160, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Fattouch, S.; Raboudi-Fattouch, F.; Ponce, J.V.G.; Forment, J.V.; Lukovic, D.; Marzouki, N.; Vidal, D.R. Concentration dependent effects of commonly used pesticides on activation versus inhibition of the quince (Cydonia Oblonga) polyphenol oxidase. Food Chem. Toxicol. 2010, 48, 957–963. [Google Scholar] [CrossRef]
- Fu, Y.; Liu, Y.-X.; Kang, T.; Sun, Y.-N.; Li, J.-Z.; Ye, F. Identification of novel inhibitors of p-hydroxyphenylpyruvate dioxygenase using receptor-based virtual screening. J. Taiwan Inst. Chem. Eng. 2019, 103, 33–43. [Google Scholar] [CrossRef]
- Liu, X.-H.; Chen, P.-Q.; Wang, B.-L.; Dong, W.-L.; Li, Y.-H.; Xie, X.-Q.; Li, Z.-M. High Throughput Receptor-Based Virtual Screening Under ZINC Database, Synthesis, and Biological Evaluation of Ketol-Acid Reductoisomerase Inhibitors. Chem. Biol. Drug Des. 2010, 75, 228–232. [Google Scholar] [CrossRef]
- Shen, H.; Li, Z.; Jiang, Y.; Pan, X.; Wu, J.; Cristofori-Armstrong, B.; Smith, J.J.; Chin, Y.K.Y.; Lei, J.; Zhou, Q.; et al. Structural basis for the modulation of voltage-gated sodium channels by animal toxins. Science 2018, 362, 1–8. [Google Scholar] [CrossRef]
- Offermann, L.R.; Chan, S.L.; Osinski, T.; Tan, Y.W.; Chew, F.T.; Sivaraman, J.; Mok, Y.-K.; Minor, W.; Chruszcz, M. The major cockroach allergen Bla g 4 binds tyramine and octopamine. Mol. Immunol. 2014, 60, 86–94. [Google Scholar] [CrossRef]
- Cai, J.; Du, X.; Wang, C.; Lin, J.; Du, X. Identification of Potential Helicoverpa armigera (Lepidoptera: Noctuidae) Sterol Carrier Protein-2 Inhibitors Through High-Throughput Virtual Screening. J. Econ. Entomol. 2017, 110, 1779–1784. [Google Scholar] [CrossRef]
- Joshi, T.; Joshi, T.; Sharma, P.; Chandra, S.; Pande, V. Molecular docking and molecular dynamics simulation approach to screen natural compounds for inhibition of Xanthomonas oryzae pv. Oryzae by targeting peptide deformylase. J. Biomol. Struct. Dyn. 2021, 39, 823–840. [Google Scholar] [CrossRef] [PubMed]
- Correy, G.J.; Zaidman, D.; Harmelin, A.; Carvalho, S.; Mabbitt, P.D.; Calaora, V.; James, P.J.; Kotze, A.C.; Jackson, C.J.; London, N. Overcoming insecticide resistance through computational inhibitor design. Proc. Natl. Acad. Sci. USA 2019, 116, 21012–21021. [Google Scholar] [CrossRef] [PubMed]
- Laughlin, J.D.; Ha, T.S.; Jones, D.N.M.; Smith, D.P. Activation of Pheromone-Sensitive Neurons Is Mediated by Conformational Activation of Pheromone-Binding Protein. Cell 2008, 133, 1255–1265. [Google Scholar] [CrossRef]
- Oliferenko, P.V.; Oliferenko, A.A.; Poda, G.I.; Osolodkin, D.I.; Pillai, G.G.; Bernier, U.R.; Tsikolia, M.; Agramonte, N.M.; Clark, G.G.; Linthicum, K.J.; et al. Promising Aedes aegypti Repellent Chemotypes Identified through Integrated QSAR, Virtual Screening, Synthesis, and Bioassay. PLoS ONE 2013, 8, e64547. [Google Scholar] [CrossRef]
- Seeliger, D.; de Groot, B.L. Ligand docking and binding site analysis with PyMOL and Autodock/Vina. J. Comput. Aided. Mol. Des. 2010, 24, 417–422. [Google Scholar] [CrossRef]
- Jones, G.; Willett, P.; Glen, R.C.; Leach, A.R.; Taylor, R. Development and validation of a genetic algorithm for flexible docking. J. Mol. Biol. 1997, 267, 727–748. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2009, 31, 455–461. [Google Scholar] [CrossRef]
- Liu, N.; Xu, Z. Using LeDock as a docking tool for computational drug design. IOP Conf. Ser. Earth Environ. Sci. 2019, 218, 012143. [Google Scholar] [CrossRef]
- Sander, T.; Freyss, J.; von Korff, M.; Rufener, C. DataWarrior: An Open-Source Program For Chemistry Aware Data Visualization And Analysis. J. Chem. Inf. Model. 2015, 55, 460–473. [Google Scholar] [CrossRef]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An open chemical toolbox. J. Cheminform. 2011, 3, 33. [Google Scholar] [CrossRef]
- Brito, N.F.; Moreira, M.F.; Melo, A.C.A. A look inside odorant-binding proteins in insect chemoreception. J. Insect Physiol. 2016, 95, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.S.; Xiao, S.; Carlson, J.R. The diverse small proteins called odorant-binding proteins. Open Biol. 2018, 8, 180208. [Google Scholar] [CrossRef] [PubMed]
- Guo, D.; Luo, J.; Zhou, Y.; Xiao, H.; He, K.; Yin, C.; Xu, J.; Li, F. ACE: An efficient and sensitive tool to detect insecticide resistance-associated mutations in insect acetylcholinesterase from RNA-Seq data. BMC Bioinform. 2017, 18, 330. [Google Scholar] [CrossRef] [PubMed]
- Pang, Y.-P.; Brimijoin, S.; Ragsdale, D.W.; Zhu, K.Y.; Suranyi, R. Novel and Viable Acetylcholinesterase Target Site for Developing Effective and Environmentally Safe Insecticides. Curr. Drug Targets 2012, 13, 471–482. [Google Scholar] [CrossRef]
- Pang, Y. Insect Acetylcholinesterase as a Target for Effective and Environmentally Safe Insecticides. In Target Recept. Control Insect Pests Part II; Elsevier Inc.: Amsterdam, The Netherlands, 2014; pp. 435–494. [Google Scholar]
- Spinelli, S.; Lagarde, A.; Iovinella, I.; Legrand, P.; Tegoni, M.; Pelosi, P.; Cambillau, C. Crystal structure of Apis mellifera OBP14, a C-minus odorant-binding protein, and its complexes with odorant molecules. Insect Biochem. Mol. Biol. 2012, 42, 41–50. [Google Scholar] [CrossRef] [PubMed]
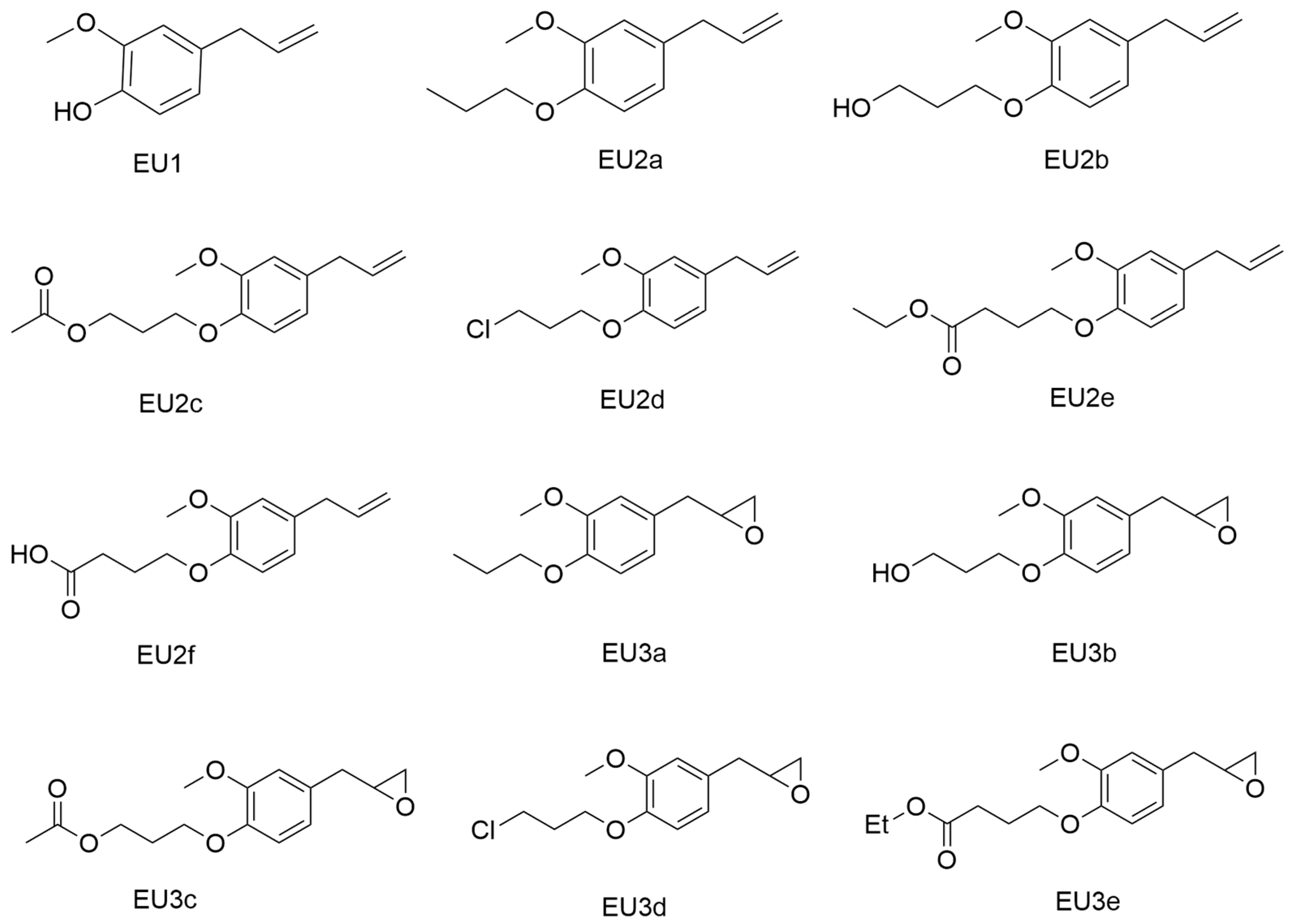

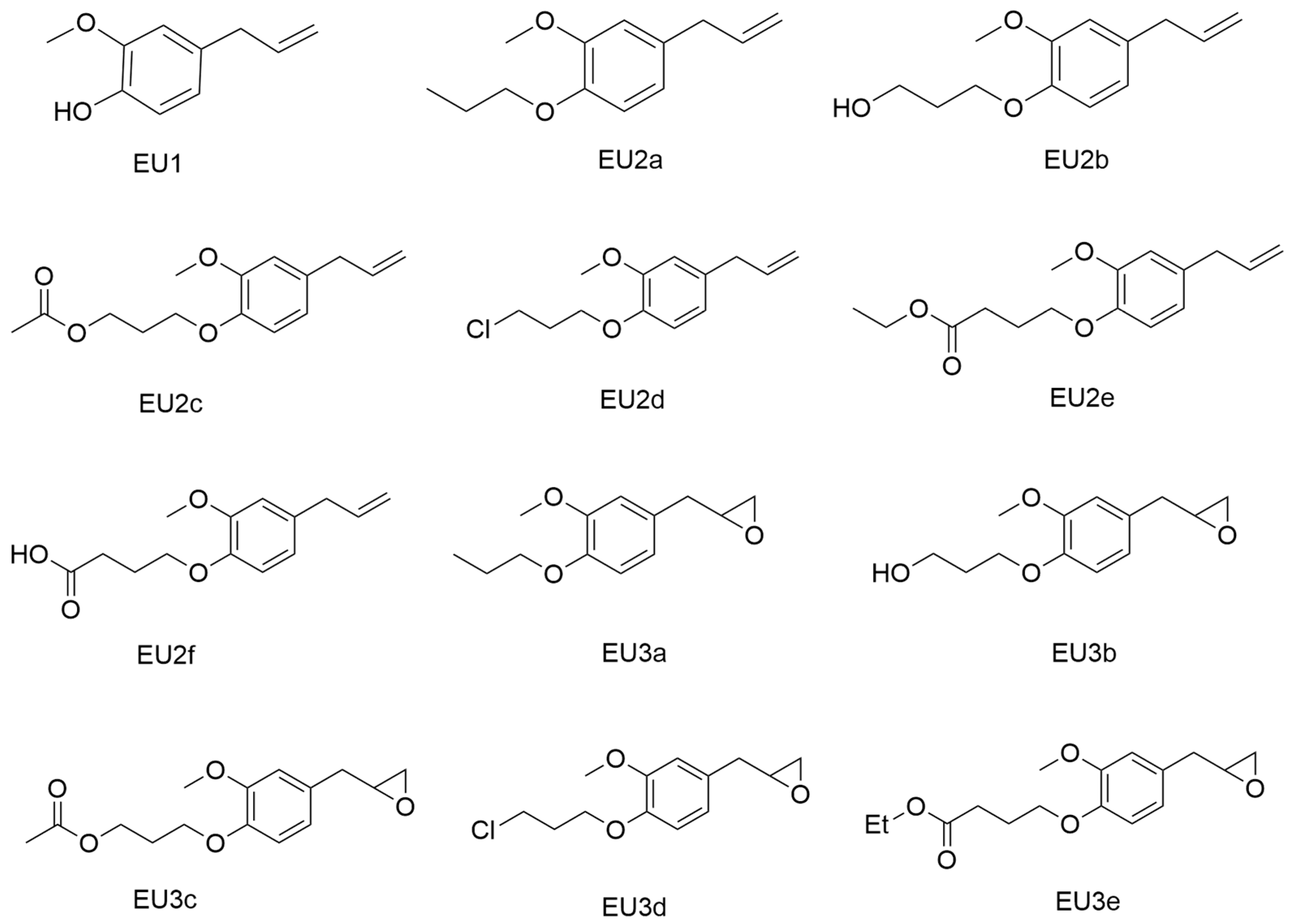{kind=link}
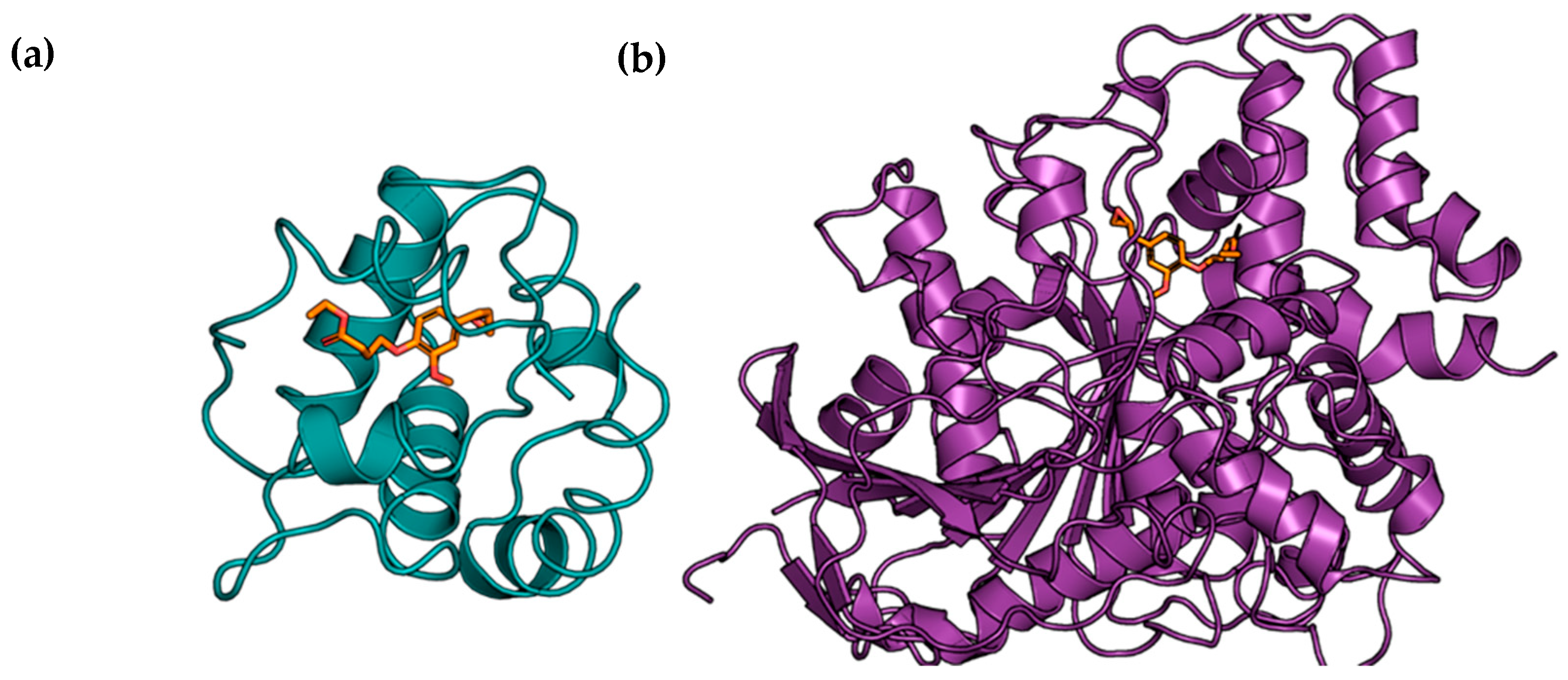
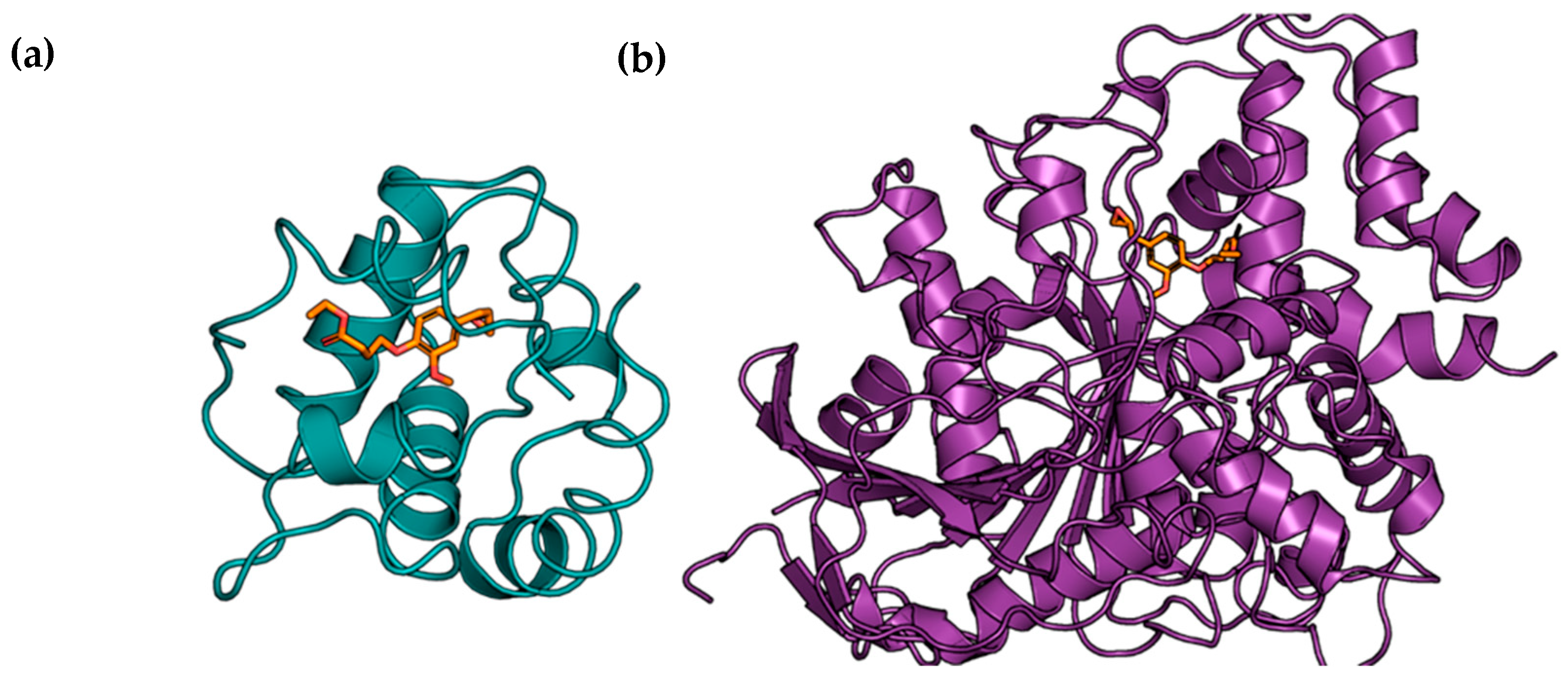{kind=link}
| Target | Organism | PDB Target | Resolution (Å) | Description | Reference |
|---|---|---|---|---|---|
| Ecdysone receptor | Heliothis virescens | 1R20 | 3.00 | VS based on 1R20 bound to an agonist as a model for the development of a receptor-based pharmacophore model. | [8] |
| 1R1K | 2.90 | VS of 2 million compounds against 1R1K, an ecdysone receptor structure bound to its known ligand Ponasterone A. | [9] | ||
| Chitinase | Ostrinia furnacalis | 3WL1 | 1.77 | Pharmacophore-based screening using two crystal structures of chitinases: 3WL1 bound to its reaction product and 3WQV bound to an inhibitor. | [10] |
| 3WQV | 2.04 | ||||
| beta-N-acetyl-D-hexosaminidase OfHex1 | 3NSN | 2.10 | VS of the ZINC database to identify OfHex1 inhibitors using 3NSN crystal structure bound to a known inhibitor. | [11] | |
| 3OZP | 2.00 | VS of the ZINC database targeting 3OZP, a crystal structure of OfHex1 bound to an inhibitor. | [12] | ||
| N-Acetylglucosamine-1-phosphate uridyltransferase (GlmU) | Xanthomonas oryzae | 2V0K | 2.30 | Homology model built for docking using 2V0K and 2VD4 as templates. 2V0K crystal structure is bound to its known ligand and 2VD4 is bound to a possible inhibitor. | [13] |
| 2VD4 | 1.90 | ||||
| Acetylcholinesterase | Aedes aegypti | 1QON | 2.72 | Search for new molecules with insecticidal activity against Ae. Aegypti using acetylcholinesterase crystal structures 1QON and 4EY6 as targets, both bound to possible inhibitors. | [14] |
| 4EY6 | 2.40 | ||||
| Drosophila melanogaster | 1DX4 | 2.70 | Homology 3D model built for vs. using 1DX4 as template. 1DX4 crystal structure is bound to a potent inhibitor. | [15] | |
| Polyphenol oxidase | Ipomoea batatas | 1BUG | 2.70 | Docking simulations using the homologous polyphenol oxidase crystal structure of sweet potato in complex with phenylthiourea, a commonly used pesticide. | [16] |
| p-hydroxyphenylpyruvate dioxygenase | Arabidopsis thaliana | 6ISD | 2.40 | Development of a receptor-ligand pharmacophore model based on the crystal structure 6ISD bound to a commonly used pesticide. The best model created was then used for VS. studies. | [17] |
| Oxidoreductase | Spinacia oleracea | 1YVE | 1.65 | Crystal structure of a plant oxireductase, 1YVE bound to its cofactor, NADPH used in VS. assays to find new inhibitors. | [18] |
| Voltage-gated sodium channel | Periplaneta americana | 6A95 | 2.60 | Crystallographic structure of a voltage-gated sodium channel NavPaS bound to a pore blocker, tetrodotoxin (TTX). | [19] |
| Octopamine receptor | Blattella germanica | 4N7C | 1.75 | Crystal structure of Bla g 4, an octopamine receptor, bound to tyramine. | [20] |
| Sterol carrier protein-2 (HaSCP-2) | Helicoverpa armigera | 4UEI | Solution NMR | Structure-based VS. of a database of commercially available compounds to find potential inhibitors of HaSCP-2. The residues Phe53, Thr128, and Gln131 were selected for the binding cavity. | [21] |
| Peptide deformylase | Xanthomonas oryzae | 5CY8 | 2.38 | Docking and VS of a library of 318 phytochemicals; 5CY8 crystal structure is bound to a possible inhibitor. | [22] |
| Alpha-esterase-7 (αE7) | Lucilia cuprina | 5TYJ | 1.75 | Computational design of potent and selective covalent inhibitors of αE7; 5TYJ and 5TYP crystal structures are bound to inhibitors: (3-bromo-5-phenoxylphenyl)boronic acid and (3-bromo-4-methylphenyl)boronic acid respectively. | [23] |
| 5TYP | 1.88 | ||||
| Odorant Binding Protein | Aedes aegypti | 5V13 | 1.84 | Search for new molecules with insecticidal activity against Ae. Aegypti using a crystal structure of a mosquito juvenile hormone-binding protein, 5V13 bound to its natural hormone. | [14] |
| Drosophila melanogaster | 2GTE | 1.40 | 2GTE crystal structure is bound to its natural ligand | [24] | |
| Anopheles gambiae | 3N7H | 1.60 | QSAR and docking studies for the rational design of mosquito repellents using the crystal structure 3K1E bound to a polyethylene glycol molecule; 3N7H crystal structure is bound to a commonly used repellent. | [25] | |
| Aedes aegypti | 3K1E | 1.85 | [25] |
| Target | PDB | PLP | ASP | ChemScore | GoldScore | Vina | LeDock |
|---|---|---|---|---|---|---|---|
| Ecdysone receptor | 1R20 | 57.3 | 27.5 | 28.1 | 52.5 | −6.4 | −4.7 |
| 1R1K | 59.3 | 26.4 | 28.3 | 54.5 | −7.1 | −5.2 | |
| Chitinase | 3WL1 | 63.0 | 40.8 | 30.1 | 60.0 | −6.9 | −4.8 |
| 3WQV | 63.4 | 40.7 | 30.6 | 55.7 | −6.5 | −4.3 | |
| beta-N-acetyl-D-hexosaminidase OfHex1 | 3NSN | 66.7 | 46.7 | 29.1 | 62.8 | −6.1 | −4.4 |
| 3OZP | 63.3 | 43.7 | 28.3 | 58.7 | −7.1 | −4.3 | |
| N-Acetylglucosamine-1-phosphate uridyltransferase (GlmU) | 2V0K | 55.0 | 24.1 | 23.3 | 54.3 | −5.9 | −4.6 |
| 2VD4 | 46.9 | 22.2 | 21.6 | 43.8 | −5.2 | −3.7 | |
| Acetylcholinesterase | 1QON | 73.3 | 48.2 | 35.3 | 62.2 | −7.6 | −5.0 |
| 4EY6 | 72.6 | 41.2 | 32.4 | 55.2 | −7.1 | −5.0 | |
| 1DX4 | 70.0 | 43.2 | 32.2 | 55.3 | −7.2 | −4.9 | |
| Polyphenol oxidase (PPO) | 1BUG | 56.7 | 27.2 | 25.9 | 56.2 | −5.2 | −4.1 |
| p-hydroxyphenylpyruvate dioxygenase | 6ISD | 57.9 | 31.6 | 24.8 | 47.8 | −6.3 | −4.3 |
| Oxidoreductase | 1YVE | 66.0 | 25.6 | 32.1 | 59.5 | −6.3 | −5.2 |
| Voltage-gated sodium channel | 6A95 | 53.1 | 23.6 | 22.3 | 56.5 | −5.8 | −4.5 |
| Octopamine receptor | 4N7C | 68.1 | 37.9 | 35.1 | 65.2 | −7.1 | −4.5 |
| Sterol carrier protein-2 (HaSCP-2) | 4UEI | 54.1 | 28.2 | 29.4 | 45.8 | −6.4 | −4.9 |
| Peptide deformylase | 5CY8 | 64.0 | 26.4 | 24.3 | 62.5 | −6.8 | −5.6 |
| α-esterase-7 | 5TYJ | 62.9 | 34.6 | 29.3 | 52.1 | −6.4 | −4.3 |
| 5TYP | 59.9 | 35.2 | 29.4 | 53.1 | −6.4 | −4.8 | |
| Odorant Binding Protein | 5V13 | 72.1 | 43.2 | 35.9 | 59.4 | −7.6 | −5.1 |
| 2GTE | 63.1 | 33.8 | 34.3 | 56.9 | −6.5 | −3.1 | |
| 3N7H | 64.8 | 34.5 | 28.9 | 56.6 | −6.3 | −4.6 | |
| 3K1E | 73.4 | 39.6 | 35.8 | 62.4 | −6.0 | −5.5 |
| Ranking | PLP | ASP | ChemScore | GoldScore | Vina | LeDock | Overall Ranking |
|---|---|---|---|---|---|---|---|
| Odorant Binding Protein | 1 | 4 | 1 | 3 | 2 | 2 | 1 |
| Acetylcholinesterase | 2 | 1 | 2 | 5 | 1 | 5 | 2 |
| Chitinase | 4 | 2 | 5 | 2 | 6 | 7 | 3 |
| Octopamine receptor | 3 | 5 | 3 | 1 | 5 | 10 | 4 |
| Peptide deformylase | 6 | 11 | 12 | 4 | 7 | 1 | 5 |
| Oxidoreductase | 5 | 12 | 4 | 6 | 11 | 4 | 6 |
| β-N-acetyl-D-hexosaminidase OfHex1 | 7 | 3 | 9 | 7 | 3 | 13 | 7 |
| Ecdysone receptor | 9 | 9 | 8 | 10 | 4 | 3 | 8 |
| α-esterase-7 | 8 | 6 | 7 | 12 | 9 | 8 | 9 |
| Sterol carrier protein-2 (HaSCP-2) | 13 | 8 | 6 | 14 | 8 | 6 | 10 |
| p-hydroxyphenylpyruvate dioxygenase | 10 | 7 | 11 | 13 | 10 | 12 | 11 |
| Polyphenol oxidase (PPO) | 11 | 10 | 10 | 9 | 14 | 14 | 12 |
| N-Acetylglucosamine-1-phosphate uridyltransferase (GlmU) | 12 | 13 | 13 | 11 | 12 | 9 | 13 |
| Voltage-gated sodium channel | 14 | 14 | 14 | 8 | 13 | 11 | 14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vieira, T.F.; Araújo, M.F.; Fernandes, M.J.G.; Pereira, D.M.; Fortes, A.G.; Castanheira, E.M.S.; Gonçalves, M.S.T.; Sousa, S.F. In Silico Identification of Protein Targets Associated to the Insecticide Activity of Eugenol Derivatives. Chem. Proc. 2021, 3, 138. https://doi.org/10.3390/ecsoc-24-08333
Vieira TF, Araújo MF, Fernandes MJG, Pereira DM, Fortes AG, Castanheira EMS, Gonçalves MST, Sousa SF. In Silico Identification of Protein Targets Associated to the Insecticide Activity of Eugenol Derivatives. Chemistry Proceedings. 2021; 3(1):138. https://doi.org/10.3390/ecsoc-24-08333
Chicago/Turabian StyleVieira, Tatiana F., Maria F. Araújo, Maria José G. Fernandes, David M. Pereira, A. Gil Fortes, Elisabete M. S. Castanheira, M. Sameiro T. Gonçalves, and Sérgio F. Sousa. 2021. "In Silico Identification of Protein Targets Associated to the Insecticide Activity of Eugenol Derivatives" Chemistry Proceedings 3, no. 1: 138. https://doi.org/10.3390/ecsoc-24-08333
APA StyleVieira, T. F., Araújo, M. F., Fernandes, M. J. G., Pereira, D. M., Fortes, A. G., Castanheira, E. M. S., Gonçalves, M. S. T., & Sousa, S. F. (2021). In Silico Identification of Protein Targets Associated to the Insecticide Activity of Eugenol Derivatives. Chemistry Proceedings, 3(1), 138. https://doi.org/10.3390/ecsoc-24-08333