
An In Silico Approach to Enzymatic Synthesis of Fucooligosaccharides Using α-l-Fucosidase from Thermotoga maritima †


 , , , , and
, , , , and
Abstract
:1. Introduction
2. Methods
2.1. Geometry Optimization and HOMO-LUMO Parameters
2.2. Molecular Docking for Hydrolysis and Transfucosylation Process
3. Results and Discussion
3.1. HOMO-LUMO Parameters
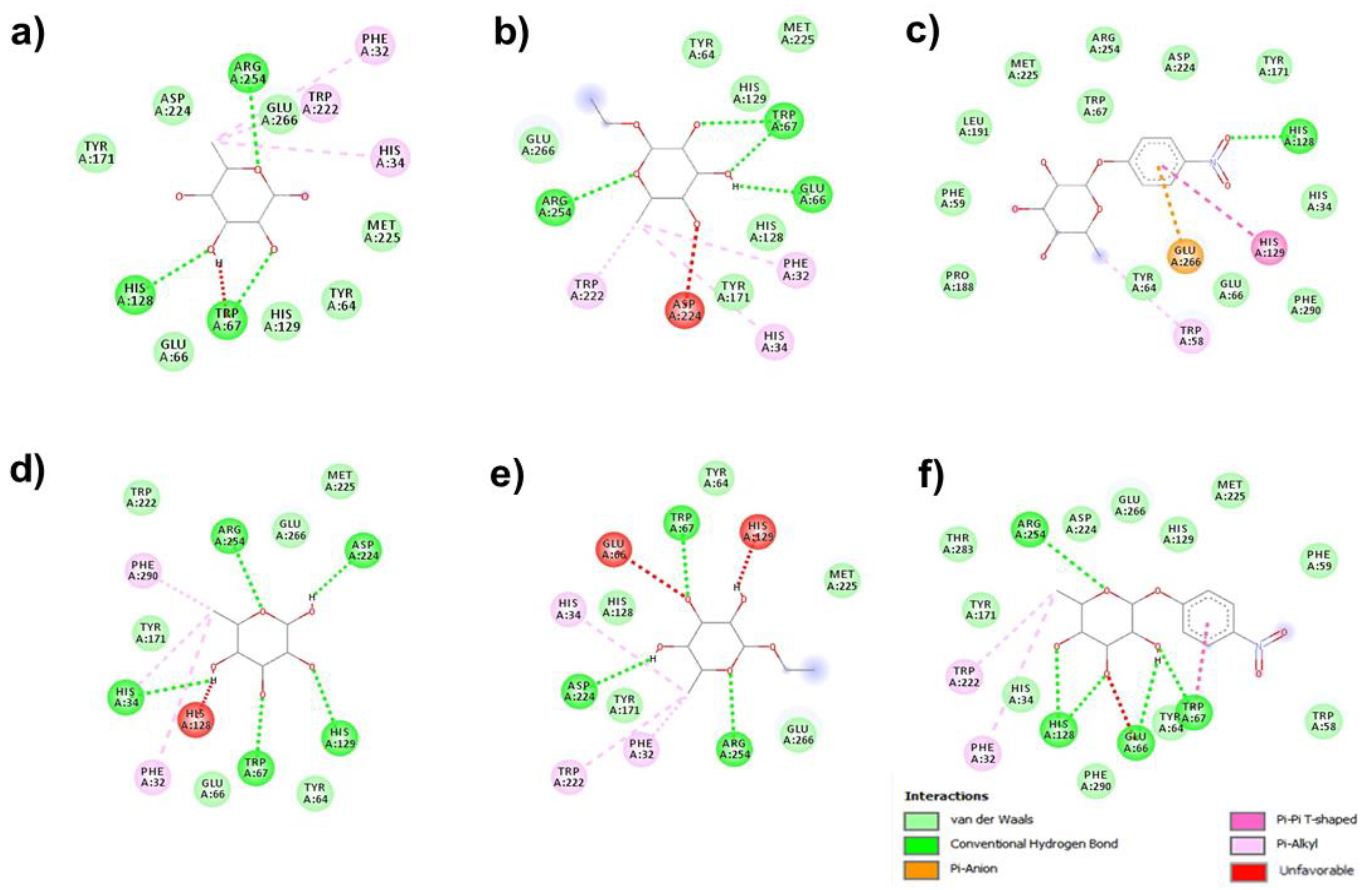
3.2. Molecular Docking for Hydrolysis Processes
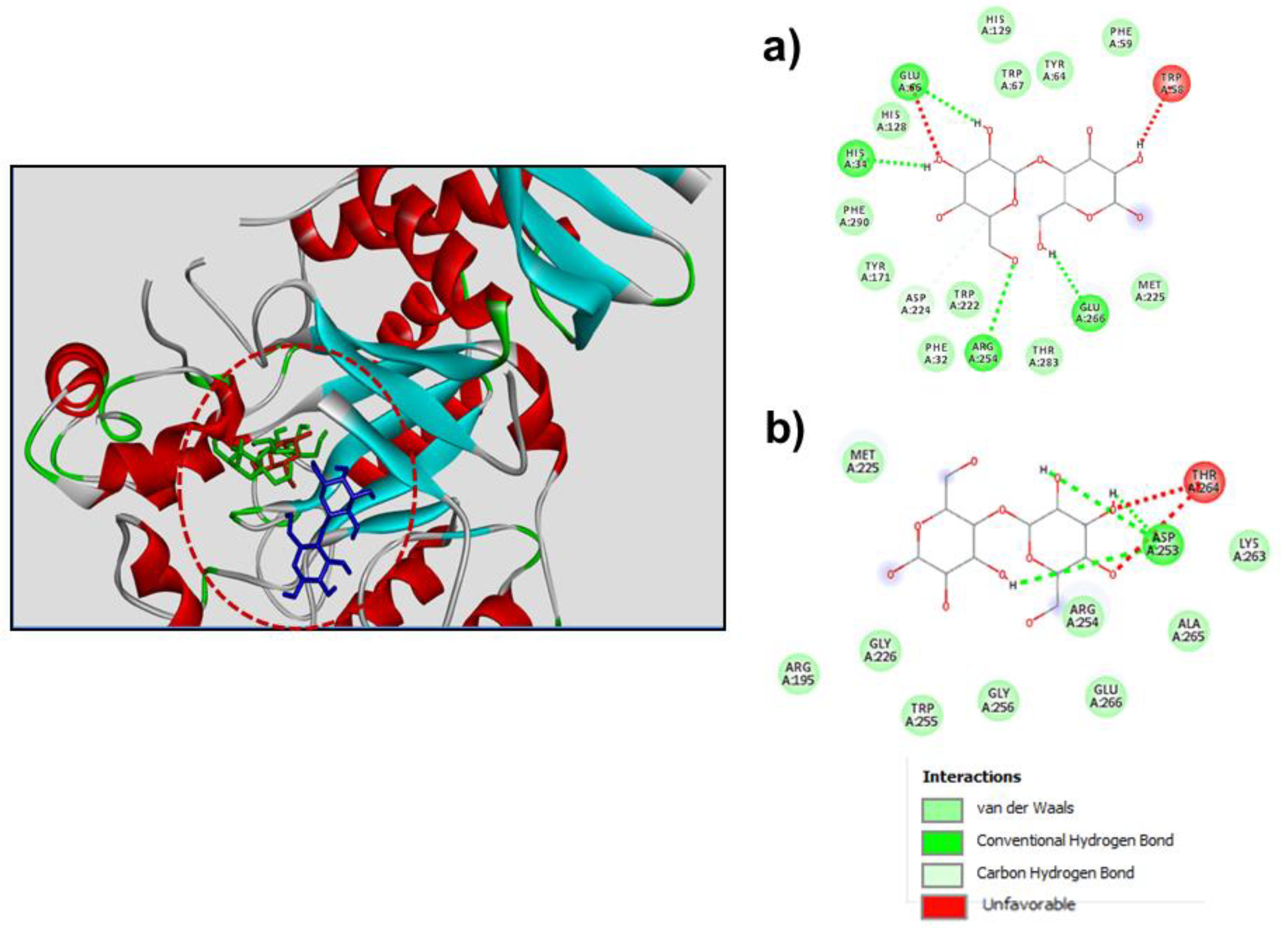
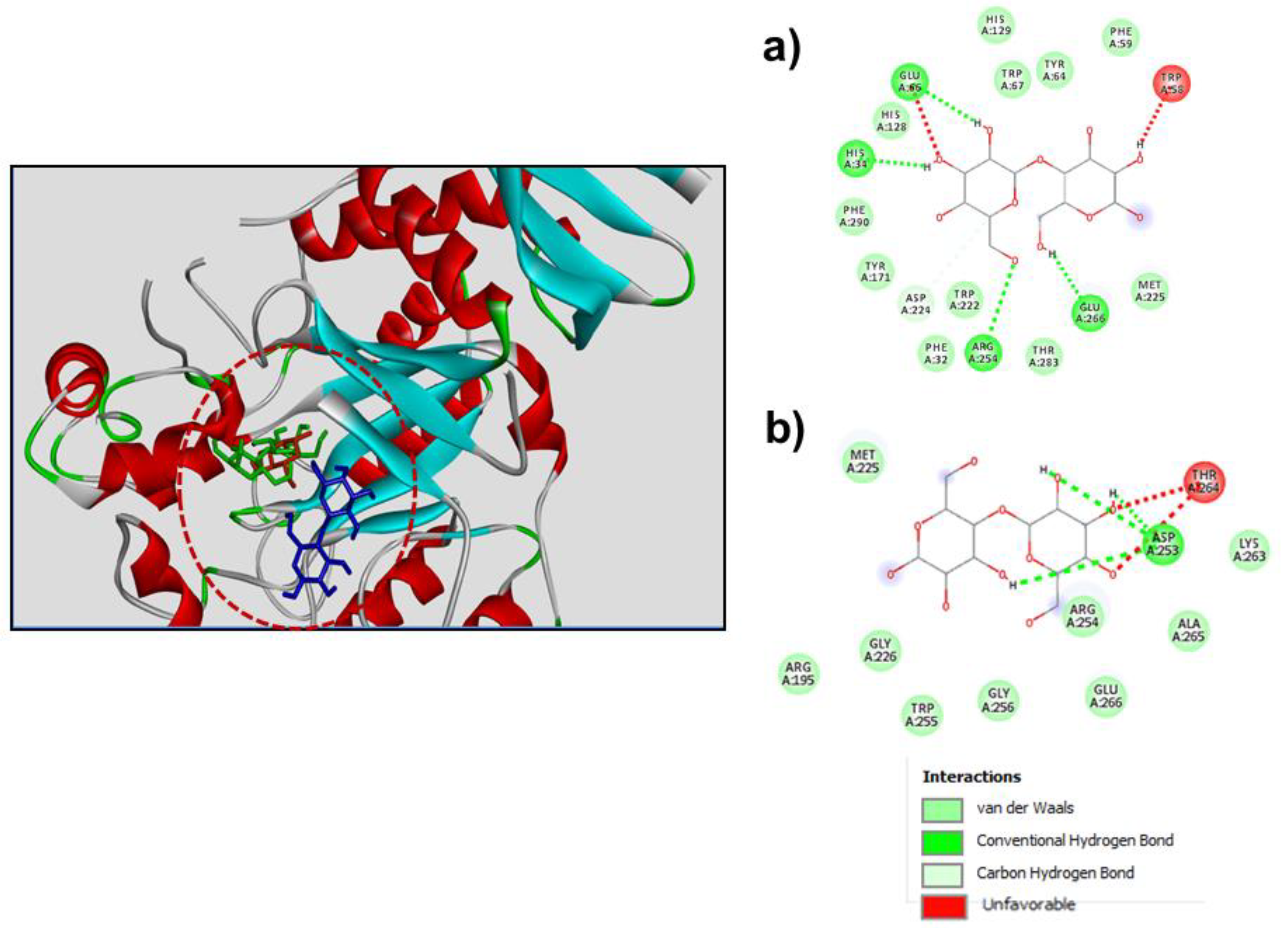
3.3. Molecular Docking for Studying Transfucosylation
4. Conclusions
Author Contributions
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pérez-Escalante, E.; Alatorre-Santamaría, S.; Castañeda-Ovando, A.; Salazar-Pereda, V.; Bautista-Ávila, M.; Cruz-Guerrero, A.E.; Flores-Aguilar, J.F.; González-Olivares, L.G. Human milk oligosaccharides as bioactive compounds in infant formula: Recent advances and trends in synthetic methods. Crit. Rev. Food Sci. Nutr. 2020, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Bych, K.; Mikš, M.H.; Johanson, T.; Hederos, M.J.; Vigsnæs, L.K.; Becker, P. Production of HMOs using microbial hosts—From cell engineering to large scale production. Curr. Opin. Biotechnol. 2019, 56, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Zeuner, B.; Meyer, A.S. Enzymatic transfucosylation for synthesis of human milk oligosaccharides. Carbohydr. Res. 2020, 493, 108029. [Google Scholar] [CrossRef] [PubMed]
- Wan, L.; Zhu, Y.; Zhang, W.; Mu, W. α-L-Fucosidases and their applications for the production of fucosylated human milk oligosaccharides. Appl. Microbiol. Biotechnol. 2020, 104, 5619–5631. [Google Scholar] [CrossRef] [PubMed]
- Guzmán-Rodríguez, F.; Alatorre-Santamaría, S.; Gómez-Ruiz, L.; Rodríguez-Serrano, G.; García-Garibay, M.; Cruz-Guerrero, A. Synthesis of a fucosylated trisaccharide via transglycosylation by α-L-fucosidase from Thermotoga maritima. Appl. Biochem. Biotechnol. 2018, 186, 681–691. [Google Scholar] [CrossRef] [PubMed]
- Shi, R.; Ma, J.; Yan, Q.; Yang, S.; Fan, Z.; Jiang, Z. Biochemical characterization of a novel α-L-fucosidase from Pedobacter sp. and its application in synthesis of 3′-fucosyllactose and 2′-fucosyllactose. Appl. Microbiol. Biotechnol. 2020, 104, 5813–5826. [Google Scholar] [CrossRef] [PubMed]
- Usvalampi, A.; Medrano, M.R.; Maaheimo, H.; Salminen, H.; Tossavainen, O.; Frey, A.D. Production and characterization of Aspergillus niger GH29 family α-fucosidase and production of a novel non-reducing 1-fucosyllactose. Glycoconj. J. 2020, 37, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Zeuner, B.; Muschiol, J.; Holck, J.; Lezyk, M.; Gedde, M.R.; Jers, C.; Mikkelsen, J.D.; Meyer, A.S. Substrate specificity and transfucosylation activity of GH29 α-L-fucosidases for enzymatic production of human milk oligosaccharides. New Biotechnol. 2018, 41, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Allouche, A.R. Gabedit—A graphical user interface for computational chemistry softwares. J. Comput. Chem. 2011, 32, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 13, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Dallakyan, S.; Olson, A.J. Small-molecule library screening by docking with PyRx. Methods Mol. Biol. 2015, 1263, 243–250. [Google Scholar] [PubMed]
- Sulzenbacher, G.; Bignon, C.; Nishimura, T.; Tarling, C.A.; Withers, S.G.; Henrissat, B.; Bourne, Y. Crystal Structure of Thermotoga maritima α-L-Fucosidase insights into the catalytic mechanism and the molecular basis for fucosidosis. J. Biol. Chem. 2004, 279, 13119–13128. [Google Scholar] [CrossRef] [PubMed]
- Dassault Systèmes BIOVIA. Discovery Studio Visualizer; v19.1.0.18287; Dassault Systèmes: San Diego, CA, USA, 2019. [Google Scholar]
- Talmaciu, M.M.; Bodoki, E.; Oprean, R. Global chemical reactivity parameters for several chiral beta-blockers from the Density Functional Theory viewpoint. Clujul Med. 2016, 89, 513–518. [Google Scholar] [CrossRef] [PubMed]
- Boukli-Hacene, F.; Merad, M.; Ghalem, S.; Soufi, W. DFT Study of the Interaction of Cu (II), Zn (II), Sn (II) with Carbohydrates in Aqueous Solution. J. Chem. 2014, 8, 1009–1017. [Google Scholar]
- Bettelheim, F.A.; Brown, W.H.; Campbell, M.K.; Farrell, S.O. Introduction to Organic and Biochemistry, 7th ed.; Cengage Learning: Belmont, CA, USA, 2010; pp. 353–355. [Google Scholar]

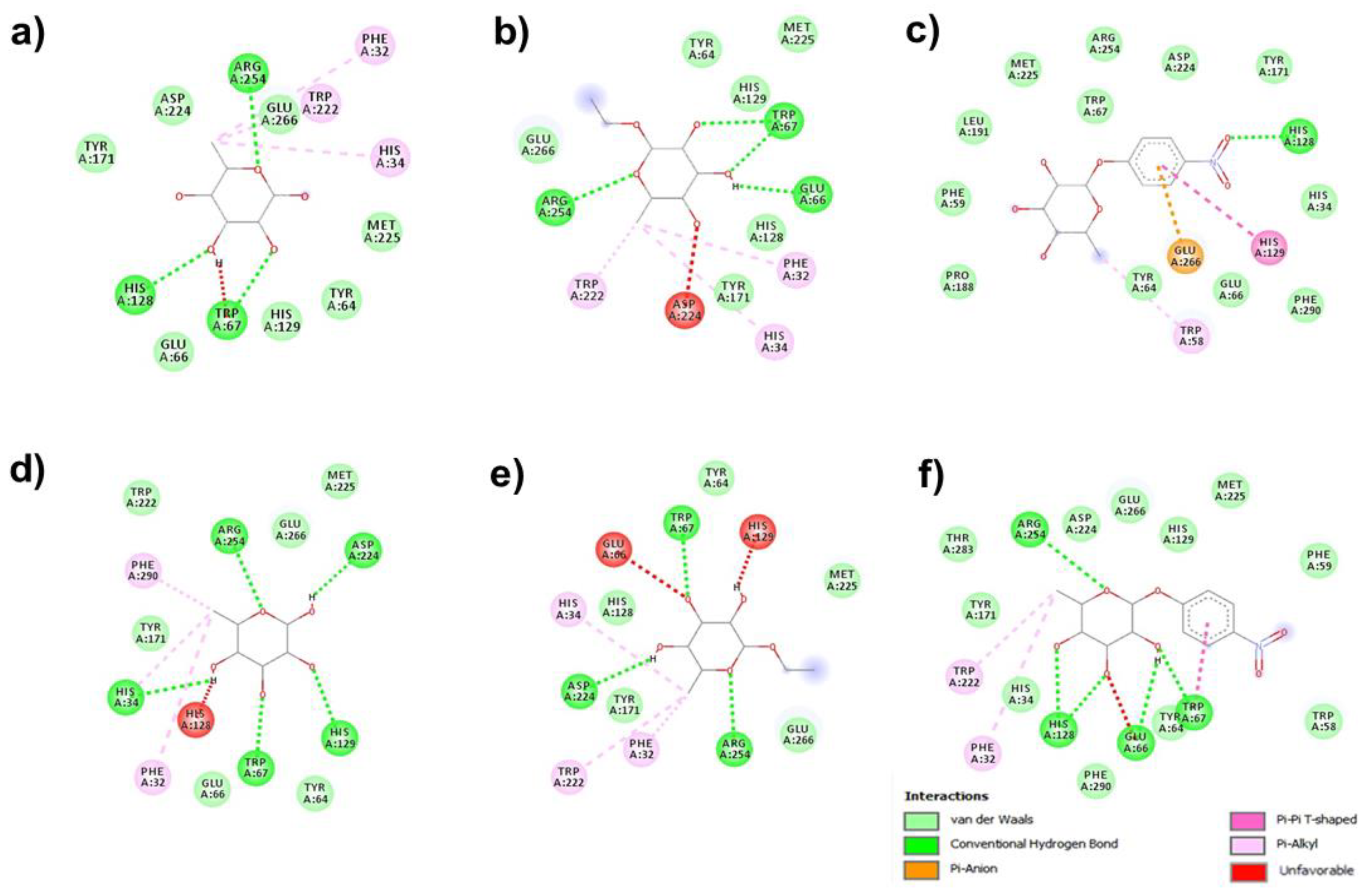

{kind=link}
{kind=link}
{kind=link}
| Fucosyl-Donor | Coupling Energy (kcal·mol−1) |
|---|---|
| α-fucose | −6.0 |
| β-fucose | −6.3 |
| α-ethyl-fucose | −5.5 |
| β-ethyl-fucose | −5.8 |
| α-pNP-fucose | −5.9 |
| β-pNP-fucose | −6.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Escalante, E.; González-Olivares, L.G.; Castañeda-Ovando, A.; Cruz-Guerrero, A.E.; Trant, J.F.; López-Orozco, W.; Mendoza-Huizar, L.H.; Alatorre-Santamaría, S. An In Silico Approach to Enzymatic Synthesis of Fucooligosaccharides Using α-l-Fucosidase from Thermotoga maritima. Chem. Proc. 2021, 3, 10. https://doi.org/10.3390/ecsoc-24-08303
Pérez-Escalante E, González-Olivares LG, Castañeda-Ovando A, Cruz-Guerrero AE, Trant JF, López-Orozco W, Mendoza-Huizar LH, Alatorre-Santamaría S. An In Silico Approach to Enzymatic Synthesis of Fucooligosaccharides Using α-l-Fucosidase from Thermotoga maritima. Chemistry Proceedings. 2021; 3(1):10. https://doi.org/10.3390/ecsoc-24-08303
Chicago/Turabian StylePérez-Escalante, Emmanuel, Luis Guillermo González-Olivares, Araceli Castañeda-Ovando, Alma Elizabeth Cruz-Guerrero, John F. Trant, Wendolyne López-Orozco, Luis Humberto Mendoza-Huizar, and Sergio Alatorre-Santamaría. 2021. "An In Silico Approach to Enzymatic Synthesis of Fucooligosaccharides Using α-l-Fucosidase from Thermotoga maritima" Chemistry Proceedings 3, no. 1: 10. https://doi.org/10.3390/ecsoc-24-08303
APA StylePérez-Escalante, E., González-Olivares, L. G., Castañeda-Ovando, A., Cruz-Guerrero, A. E., Trant, J. F., López-Orozco, W., Mendoza-Huizar, L. H., & Alatorre-Santamaría, S. (2021). An In Silico Approach to Enzymatic Synthesis of Fucooligosaccharides Using α-l-Fucosidase from Thermotoga maritima. Chemistry Proceedings, 3(1), 10. https://doi.org/10.3390/ecsoc-24-08303