
Noncoding RNAs Interactions in Hepatic Stellate Cells during Hepatic Fibrosis



Abstract
:1. Introduction
2. MicroRNAs and HSCs
2.1. Profibrotic miRNAs
2.2. Anti-Fibrotic miRNAs
3. Long Noncoding RNAs and HSCs

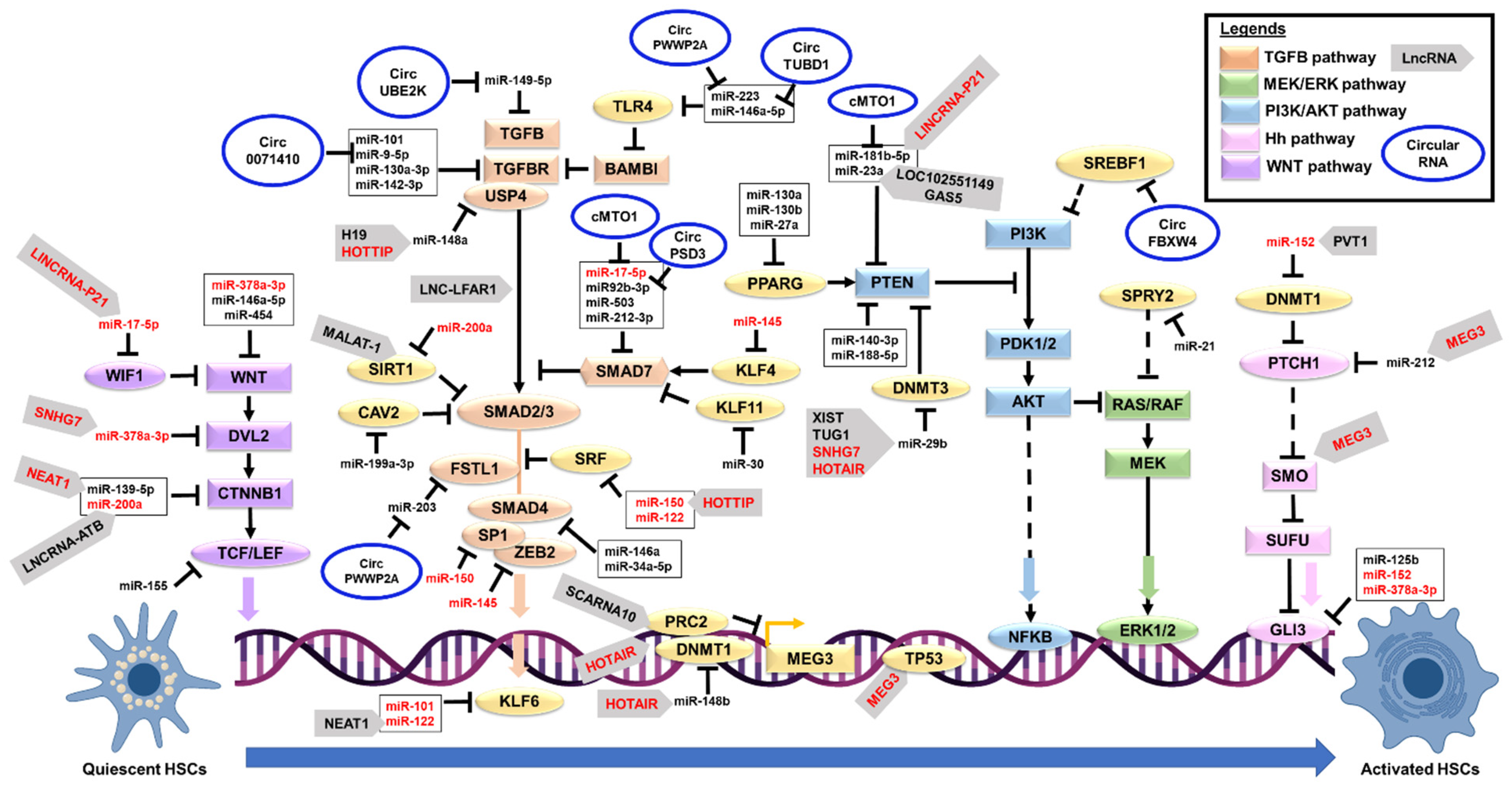{kind=link}
| LncRNA | Expression | Target Molecule | Disease Model | Function | Role in Fibrosis | References |
|---|---|---|---|---|---|---|
| GPR137B-PS | Up | miR-200a-3p/CXCL14 | Mouse CCI(4)-treated | HSC activation and collagen production | Profibrotic | [119] |
| H19 | Up | miR-148a/Usp4 | Mouse CCI(4)-treated, Human LX-2, L02 and HEK293T/17 cell lines | HSC activation via TGFB1 pathway | Profibrotic | [114] |
| HOTAIR | Up | miR-29b/Dnmt3b/Pten | Patient samples, Mouse CCI(4)-treated | HSC activation and collagen production | Profibrotic | [120] |
| Up | miR-148b/DNMT1 | Patient samples, Mouse CCI(4)-treated, Human LX-2, AML-12 and RAW264.7 cell lines | HSC activation and collagen production | Profibrotic | [121] | |
| Up | PRC2 | Patient samples, Mouse CCI(4)-treated, Human LX-2, AML-12 and RAW264.7 cell lines | HSC activation and collagen production | Profibrotic | [121] | |
| HOTTIP | Up | miR-148a/TGFBR | Patient samples, Mouse CCI(4)-treated | HSC activation | Profibrotic | [116] |
| Up | miR-150/Srf | Mouse CCI(4)-treated | Activates HSC proliferation | Profibrotic | [117] | |
| LINC-SCRG1 | Up | ZFP36 | Patient samples, Human LX-2 cell line | Activates HSC proliferation and ECM protein accumulation | Profibrotic | [122] |
| Lnc-Lfar1 | Up | Smad2/3 | Mouse CCI(4)-treated, AML-12 and HEK293T cell lines | HSC activation via TGFB1 and Notch pathways | Profibrotic | [113] |
| LNCRNA-ATB | Up | miR-200a/CTNNB1 | Patient samples, Human LX-2 cell line | HSC activation via Wnt/β-catenin pathway | Profibrotic | [123] |
| MALAT1 | Up | miR-101b/RAC1 | Mouse CCI(4)-treated | HSC activation and collagen production | Profibrotic | [124] |
| Up | SIRT1 | Mouse CCI(4)-treated, Human LX-2 cell line | HSC activation and collagen production | Profibrotic | [118] | |
| MEG8 | Up | NOTCH2/3 | Human LX-2 and AML-12 cell line | Suppresses HSC activation and collagen production | Anti-fibrotic | [125] |
| NEAT1 | Up | miR-122/Klf6 | Mouse CCI(4)-treated | HSC activation and collagen production | Profibrotic | [126] |
| Up | miR-148a-3p/miR-22-3p/Cyth3 | Mouse CCI(4)-treated | HSC activation and collagen production | Profibrotic | [127] | |
| Up | miR-139-5p/CTNNB1 | Mouse CCI(4)-treated, Human LX-2 cell line | Activates HSC proliferation | Profibrotic | [128] | |
| Pvt1 | Up | miR-152/Dnmt1/Ptch1 | Mouse CCI(4)-treated | HSC activation and collagen production | Profibrotic | [129,130] |
| Scarna10 | Up | Prc2 | Patient samples, AML-12 cell line; Mouse CCI(4)-treated | HSC activation | Profibrotic | [131] |
| Snhg7 | Up | miR-29b/Dnmt3a | Mouse CCI(4)-treated | Activates HSC proliferation | Profibrotic | [132] |
| Up | miR-378a-3p/Dvl2 | Patient samples; Mouse CCI(4)-treated, Human LX-2 cell line | Activates HSC proliferation | Profibrotic | [133] | |
| Tug1 | Up | miR-29b | Patient samples, Mouse CCI(4)-treated, Human LX-2 and HEK293T cell lines | HSC activation and profibrogenic factors | Profibrotic | [134] |
| XIST | Up | miR-29b/HMGB1 | Mouse model, Human LX-2 cell line | HSC activation and profibrogenic factors | Profibrotic | [135] |
| Gas5 | Down | miR-222/CDKN1B | Patient samples, Mouse CCI(4)-treated | Suppresses HSC activation and collagen production | Anti-fibrotic | [136] |
| Down | miR-23a/Pten | Rat CCI(4)-treated | Suppresses HSC activation | Anti-fibrotic | [39] | |
| HIF1A-AS1 | Down | TET3 | Human LX-2 cell line | Suppresses HSC proliferation | Anti-fibrotic | [137] |
| LINCRNA-P21 | Down | miR-181b/PTEN | Patient samples, Human LX-2 cell line | Suppresses HSC activation and collagen production | Anti-fibrotic | [138] |
| Down | miR-17-5p | Mouse HSC cells | HSC proliferation upon Salvianolic acid B treatment | Profibrotic | [139] | |
| Loc102551149 | Down | miR-23a/Pten | Rat NDMA-treated, Rat HSC-T6 cell line | Suppresses HSC activation | Anti-fibrotic | [140] |
| MEG3 | Down | TP53 | Mouse CCI(4)-treated, Human LX-2 cell line | Suppresses HSC proliferation and ECM proteins | Anti-fibrotic | [141] |
| Down | miR-212/Ptch1/ Smo | Patient samples, Mouse CCI(4)-treated | Suppresses HSC proliferation and ECM proteins | Anti-fibrotic | [142] |
4. Circular RNAs and HSCs
5. Noncoding RNA Interactions in HSCs
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Weiskirchen, R.; Weiskirchen, S.; Tacke, F. Organ and tissue fibrosis: Molecular signals, cellular mechanisms and translational implications. Mol. Aspects Med. 2019, 65, 2–15. [Google Scholar] [CrossRef]
- Parola, M.; Pinzani, M. Liver fibrosis: Pathophysiology, pathogenetic targets and clinical issues. Mol. Aspects Med. 2019, 65, 37–55. [Google Scholar] [CrossRef]
- Cai, X.; Wang, J.; Wang, J.; Zhou, Q.; Yang, B.; He, Q.; Weng, Q. Intercellular crosstalk of hepatic stellate cells in liver fibrosis: New insights into therapy. Pharmacol. Res. 2020, 155, 104720. [Google Scholar] [CrossRef]
- Kisseleva, T.; Brenner, D.A. Hepatic stellate cells and the reversal of fibrosis. J. Gastroenterol. Hepatol. 2006, 21, S84–S87. [Google Scholar] [CrossRef]
- Yan, Y.; Zeng, J.; Xing, L.; Li, C. Extra- and intra-cellular mechanisms of hepatic stellate cell activation. Biomedicines 2021, 9, 1014. [Google Scholar] [CrossRef] [PubMed]
- Senoo, H.; Mezaki, Y.; Fujiwara, M. The stellate cell system (vitamin a-storing cell system). Anat. Sci. Int. 2017, 92, 387–455. [Google Scholar] [CrossRef]
- Fujita, T.; Narumiya, S. Roles of hepatic stellate cells in liver inflammation: A new perspective. Inflamm. Regen. 2016, 36, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faubion, W.A.; Gores, G.J. Death receptors in liver biology and pathobiology. Hepatology 1999, 29, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Taimr, P.; Higuchi, H.; Kocova, E.; Rippe, R.A.; Friedman, S.; Gores, G.J. Activated stellate cells express the trail receptor-2/death receptor-5 and undergo trail-mediated apoptosis. Hepatology 2003, 37, 87–95. [Google Scholar] [CrossRef]
- Saile, B.; Eisenbach, C.; Dudas, J.; El-Armouche, H.; Ramadori, G. Interferon-gamma acts proapoptotic on hepatic stellate cells (hsc) and abrogates the antiapoptotic effect of interferon-alpha by an hsp70-dependant pathway. Eur. J. Cell Biol. 2004, 83, 469–476. [Google Scholar] [CrossRef]
- Varela-Rey, M.; Montiel-Duarte, C.; Osés-Prieto, J.A.; López-Zabalza, M.J.; Jaffrèzou, J.P.; Rojkind, M.; Iraburu, M.J. P38 mapk mediates the regulation of alpha1 (i) procollagen mrna levels by tnf-alpha and tgf-beta in a cell line of rat hepatic stellate cells (1). FEBS Lett. 2002, 528, 133–138. [Google Scholar] [CrossRef] [Green Version]
- Sulaiman, S.A.; Muhsin, N.I.A.; Jamal, R. Regulatory non-coding rnas network in non-alcoholic fatty liver disease. Front. Physiol. 2019, 10, 279. [Google Scholar] [CrossRef]
- He, Z.; Yang, D.; Fan, X.; Zhang, M.; Li, Y.; Gu, X.; Yang, M. The roles and mechanisms of lncrnas in liver fibrosis. Int. J. Mol. Sci. 2020, 21, 1482. [Google Scholar] [CrossRef] [Green Version]
- Farooq, R.; Dongmin, L. Non-coding rna associated competitive endogenous rna regulatory network: Novel therapeutic approach in liver fibrosis. Curr. Gene Ther. 2019, 19, 305–317. [Google Scholar]
- Fang, Z.; Dou, G.; Wang, L. Micrornas in the pathogenesis of nonalcoholic fatty liver disease. Int. J. Biol. Sci. 2021, 17, 1851–1863. [Google Scholar] [CrossRef] [PubMed]
- Tomasello, L.; Distefano, R.; Nigita, G.; Croce, C.M. The microrna family gets wider: The isomirs classification and role. Front. Cell Dev. Biol. 2021, 9, 668648. [Google Scholar] [CrossRef] [PubMed]
- Medley, J.C.; Panzade, G.; Zinovyeva, A.Y. Microrna strand selection: Unwinding the rules. Wiley Interdiscip Rev. RNA 2021, 12, e1627. [Google Scholar] [CrossRef]
- Salim, U.; Kumar, A.; Kulshreshtha, R.; Vivekanandan, P. Biogenesis, characterization, and functions of mirtrons. Wiley Interdiscip Rev. RNA 2021, e1680. [Google Scholar] [CrossRef] [PubMed]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2018, 47, D155–D162. [Google Scholar] [CrossRef]
- Chipman, L.B.; Pasquinelli, A.E. Mirna targeting: Growing beyond the seed. Trends Genet. 2019, 35, 215–222. [Google Scholar] [CrossRef]
- Ab Mutalib, N.-S.; Sulaiman, S.A.; Jamal, R. Chapter 6—computational tools for microrna target prediction. In Computational Epigenetics and Diseases; Wei, L.K., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 79–105. [Google Scholar]
- Li, G.; Li, J.; Li, C.; Qi, H.; Dong, P.; Zheng, J.; Yu, F. Microrna-125a-5p contributes to hepatic stellate cell activation through targeting fih1. Cell Physiol. Biochem. 2016, 38, 1544–1552. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Tan, W.; Cheng, S.; Wang, H.; Ye, S.; Yu, C.; He, Y.; Zeng, J.; Cen, J.; Hu, J.; et al. Upregulation of microrna-126 in hepatic stellate cells may affect pathogenesis of liver fibrosis through the nf-κb pathway. DNA Cell Biol. 2015, 34, 470–480. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Wang, J.; Lu, H.; Zhang, G.; Liu, Y.; Wang, J.; Zhang, Y.; Shang, H.; Ji, H.; Chen, X.; et al. Microrna-130a and -130b enhance activation of hepatic stellate cells by suppressing pparγ expression: A rat fibrosis model study. Biochem. Biophys. Res. Commun. 2015, 465, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, Z.; Wang, Y.; Li, X.; Yang, W.; Wei, S.; Shi, C.; Qiu, J.; Ni, M.; Rao, J.; et al. Mirna-130b-5p promotes hepatic stellate cell activation and the development of liver fibrosis by suppressing sirt4 expression. J. Cell. Mol. Med. 2021, 25, 7381–7394. [Google Scholar] [CrossRef]
- Wu, S.M.; Li, T.H.; Yun, H.; Ai, H.W.; Zhang, K.H. Mir-140-3p knockdown suppresses cell proliferation and fibrogenesis in hepatic stellate cells via pten-mediated akt/mtor signaling. Yonsei Med. J. 2019, 60, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Wang, X.; Si, C.; Duan, Y.; Chen, B.; Liang, H.; Yang, D. Downregulation of mir-141 deactivates hepatic stellate cells by targeting the pten/akt/mtor pathway. Int. J. Mol. Med. 2020, 46, 406–414. [Google Scholar] [CrossRef]
- Men, R.; Wen, M.; Zhao, M.; Dan, X.; Yang, Z.; Wu, W.; Wang, M.H.; Liu, X.; Yang, L. Mircorna-145 promotes activation of hepatic stellate cells via targeting krüppel-like factor 4. Sci. Rep. 2017, 7, 40468. [Google Scholar] [CrossRef]
- Yu, F.; Guo, Y.; Chen, B.; Dong, P.; Zheng, J. Microrna-17-5p activates hepatic stellate cells through targeting of smad7. Lab. Investig. 2015, 95, 781–789. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Wu, C.; Xu, Z.; Xia, P.; Dong, P.; Chen, B.; Yu, F. Hepatic stellate cell is activated by microrna-181b via pten/akt pathway. Mol. Cell. Biochem. 2015, 398, 1–9. [Google Scholar] [CrossRef]
- Wang, B.; Li, W.; Guo, K.; Xiao, Y.; Wang, Y.; Fan, J. Mir-181b promotes hepatic stellate cells proliferation by targeting p27 and is elevated in the serum of cirrhosis patients. Biochem. Biophys. Res. Commun. 2012, 421, 4–8. [Google Scholar] [CrossRef]
- Riaz, F.; Chen, Q.; Lu, K.; Osoro, E.K.; Wu, L.; Feng, L.; Zhao, R.; Yang, L.; Zhou, Y.; He, Y.; et al. Inhibition of mir-188-5p alleviates hepatic fibrosis by significantly reducing the activation and proliferation of hscs through pten/pi3k/akt pathway. J. Cell. Mol. Med. 2021, 25, 4073–4087. [Google Scholar] [CrossRef]
- Yang, X.; Ma, L.; Wei, R.; Ye, T.; Zhou, J.K.; Wen, M.; Men, R.; Aqeilan, R.I.; Peng, Y.; Yang, L. Twist1-induced mir-199a-3p promotes liver fibrosis by suppressing caveolin-2 and activating tgf-β pathway. Signal Transduct. Target. Ther. 2020, 5, 75. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Tang, N.; Wu, K.; Dai, W.; Ye, C.; Shi, J.; Zhang, J.; Ning, B.; Zeng, X.; Lin, Y. Mir-21 simultaneously regulates erk1 signaling in hsc activation and hepatocyte emt in hepatic fibrosis. PLoS ONE 2014, 9, e108005. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Zhang, Z.; Zhang, Y.; Li, W.; Zheng, W.; Yu, J.; Wang, B.; Chen, L.; Zhuo, Q.; Chen, L.; et al. Microrna-212 activates hepatic stellate cells and promotes liver fibrosis via targeting smad7. Biochem. Biophys. Res. Commun. 2018, 496, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Yang, X.; Wei, R.; Ye, T.; Zhou, J.K.; Wen, M.; Men, R.; Li, P.; Dong, B.; Liu, L.; et al. Microrna-214 promotes hepatic stellate cell activation and liver fibrosis by suppressing sufu expression. Cell Death Dis. 2018, 9, 718. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Enomoto, M.; Fujii, H.; Sekiya, Y.; Yoshizato, K.; Ikeda, K.; Kawada, N. Microrna-221/222 upregulation indicates the activation of stellate cells and the progression of liver fibrosis. Gut 2012, 61, 1600–1609. [Google Scholar] [CrossRef]
- Cheng, R.; Xu, H.; Hong, Y. Mir221 regulates tgf-β1-induced hsc activation through inhibiting autophagy by directly targeting lamp2. Mol. Med. Rep. 2021, 24, 777. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Li, S.; Wang, X.; Si, L.; Ma, R.; Bao, L.; Bo, A. Lncrna gas5 restrains ccl(4)-induced hepatic fibrosis by targeting mir-23a through the pten/pi3k/akt signaling pathway. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 316, G539–G550. [Google Scholar] [CrossRef]
- Zhang, H.; Yan, X.L.; Guo, X.X.; Shi, M.J.; Lu, Y.Y.; Zhou, Q.M.; Chen, Q.L.; Hu, Y.Y.; Xu, L.M.; Huang, S.; et al. Mir-27a as a predictor for the activation of hepatic stellate cells and hepatitis b virus-induced liver cirrhosis. Oncotarget 2017, 9, 1075. [Google Scholar] [CrossRef] [Green Version]
- Xie, X.; Dou, C.Y.; Zhou, Y.; Zhou, Q.; Tang, H.B. Microrna-503 targets mothers against decapentaplegic homolog 7 enhancing hepatic stellate cell activation and hepatic fibrosis. Dig. Dis. Sci. 2021, 66, 1928–1939. [Google Scholar] [CrossRef]
- Xu, T.; Pan, L.; Li, L.; Hu, S.; Zhou, H.; Yang, C.; Yang, J.; Li, H.; Liu, Y.; Meng, X.; et al. Microrna-708 modulates hepatic stellate cells activation and enhances extracellular matrix accumulation via direct targeting tmem88. J. Cell. Mol. Med. 2020, 24, 7127–7140. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Tao, Q.; Zhou, Y.; Chen, Q.; Li, L.; Hu, S.; Liu, Y.; Zhang, Y.; Shu, J.; Zhang, X.; et al. Microrna-708 represses hepatic stellate cells activation and proliferation by targeting zeb1 through wnt/β-catenin pathway. Eur. J. Pharmacol. 2020, 871, 172927. [Google Scholar] [CrossRef]
- Tao, L.; Xue, D.; Shen, D.; Ma, W.; Zhang, J.; Wang, X.; Zhang, W.; Wu, L.; Pan, K.; Yang, Y.; et al. Microrna-942 mediates hepatic stellate cell activation by regulating bambi expression in human liver fibrosis. Arch. Toxicol. 2018, 92, 2935–2946. [Google Scholar] [CrossRef] [PubMed]
- Tu, X.; Zhang, H.; Zhang, J.; Zhao, S.; Zheng, X.; Zhang, Z.; Zhu, J.; Chen, J.; Dong, L.; Zang, Y.; et al. Microrna-101 suppresses liver fibrosis by targeting the tgfβ signalling pathway. J. Pathol. 2014, 234, 46–59. [Google Scholar] [CrossRef]
- Li, J.; Ghazwani, M.; Zhang, Y.; Lu, J.; Li, J.; Fan, J.; Gandhi, C.R.; Li, S. Mir-122 regulates collagen production via targeting hepatic stellate cells and suppressing p4ha1 expression. J. Hepatol. 2013, 58, 522–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabater, L.; Locatelli, L.; Oakley, F.; Hardy, T.; French, J.; Robinson, S.M.; Sen, G.; Mann, D.A.; Mann, J. Rna sequencing reveals changes in the micrornaome of transdifferentiating hepatic stellate cells that are conserved between human and rat. Sci. Rep. 2020, 10, 21708. [Google Scholar] [CrossRef]
- Zeng, C.; Wang, Y.L.; Xie, C.; Sang, Y.; Li, T.J.; Zhang, M.; Wang, R.; Zhang, Q.; Zheng, L.; Zhuang, S.M. Identification of a novel tgf-β-mir-122-fibronectin 1/serum response factor signaling cascade and its implication in hepatic fibrogenesis. Oncotarget 2015, 6, 12224–12233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Z.; Li, L.; Ran, J.; Chu, G.; Gao, H.; Guo, L.; Chen, J. Mir-125b acts as anti-fibrotic therapeutic target through regulating gli3 in vivo and in vitro. Ann. Hepatol. 2019, 18, 825–832. [Google Scholar] [CrossRef]
- Guo, C.J.; Pan, Q.; Xiong, H.; Qiao, Y.Q.; Bian, Z.L.; Zhong, W.; Sheng, L.; Li, H.; Shen, L.; Hua, J.; et al. Dynamic expression of mir-126 * and its effects on proliferation and contraction of hepatic stellate cells. FEBS Lett. 2013, 587, 3792–3801. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Du, J.; Niu, X.; Fu, N.; Wang, R.; Zhang, Y.; Zhao, S.; Sun, D.; Nan, Y. Mir-130a-3p attenuates activation and induces apoptosis of hepatic stellate cells in nonalcoholic fibrosing steatohepatitis by directly targeting tgfbr1 and tgfbr2. Cell Death Dis. 2017, 8, e2792. [Google Scholar] [CrossRef] [Green Version]
- He, C.; Shu, B.; Zhou, Y.; Zhang, R.; Yang, X. The mir-139-5p/peripheral myelin protein 22 axis modulates tgf-β-induced hepatic stellate cell activation and ccl(4)-induced hepatic fibrosis in mice. Life Sci. 2021, 276, 119294. [Google Scholar] [CrossRef]
- Yang, X.; Dan, X.; Men, R.; Ma, L.; Wen, M.; Peng, Y.; Yang, L. Mir-142-3p blocks tgf-β-induced activation of hepatic stellate cells through targeting tgfβri. Life Sci. 2017, 187, 22–30. [Google Scholar] [CrossRef]
- Zhou, D.D.; Wang, X.; Wang, Y.; Xiang, X.J.; Liang, Z.C.; Zhou, Y.; Xu, A.; Bi, C.H.; Zhang, L. Microrna-145 inhibits hepatic stellate cell activation and proliferation by targeting zeb2 through wnt/β-catenin pathway. Mol. Immunol. 2016, 75, 151–160. [Google Scholar] [CrossRef]
- He, Y.; Huang, C.; Sun, X.; Long, X.R.; Lv, X.W.; Li, J. Microrna-146a modulates tgf-beta1-induced hepatic stellate cell proliferation by targeting smad4. Cell. Signal. 2012, 24, 1923–1930. [Google Scholar] [CrossRef]
- Du, J.; Niu, X.; Wang, Y.; Kong, L.; Wang, R.; Zhang, Y.; Zhao, S.; Nan, Y. Mir-146a-5p suppresses activation and proliferation of hepatic stellate cells in nonalcoholic fibrosing steatohepatitis through directly targeting wnt1 and wnt5a. Sci. Rep. 2015, 5, 16163. [Google Scholar] [CrossRef]
- Yuan, B.Y.; Chen, Y.H.; Wu, Z.F.; Zhuang, Y.; Chen, G.W.; Zhang, L.; Zhang, H.G.; Cheng, J.C.; Lin, Q.; Zeng, Z.C. Microrna-146a-5p attenuates fibrosis-related molecules in irradiated and tgf-beta1-treated human hepatic stellate cells by regulating ptpra-src signaling. Radiat. Res. 2019, 192, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Lin, Z.; Dong, P.; Lu, Z.; Gao, S.; Chen, X.; Wu, C.; Yu, F. Activation of hepatic stellate cells is suppressed by microrna-150. Int. J. Mol. Med. 2013, 32, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Zhang, L.; Zhao, X.; Cao, J.; Li, J.; Chu, G. Downregulation of mir-152 contributes to the progression of liver fibrosis via targeting gli3 in vivo and in vitro. Exp. Ther. Med. 2019, 18, 425–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, W.; Zhao, J.; Tang, N.; Zeng, X.; Wu, K.; Ye, C.; Shi, J.; Lu, C.; Ning, B.; Zhang, J.; et al. Microrna-155 attenuates activation of hepatic stellate cell by simultaneously preventing emt process and erk1 signalling pathway. Liver Int. 2015, 35, 1234–1243. [Google Scholar] [CrossRef]
- Ju, B.; Nie, Y.; Yang, X.; Wang, X.; Li, F.; Wang, M.; Wang, C.; Zhang, H. Mir-193a/b-3p relieves hepatic fibrosis and restrains proliferation and activation of hepatic stellate cells. J. Cell. Mol. Med. 2019, 23, 3824–3832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.C.; Chen, R.; Luo, X.; Li, Z.H.; Luo, S.Z.; Xu, M.Y. Microrna-194 inactivates hepatic stellate cells and alleviates liver fibrosis by inhibiting akt2. World J. Gastroenterol. 2019, 25, 4468–4480. [Google Scholar] [CrossRef]
- Sekiya, Y.; Ogawa, T.; Iizuka, M.; Yoshizato, K.; Ikeda, K.; Kawada, N. Down-regulation of cyclin e1 expression by microrna-195 accounts for interferon-β-induced inhibition of hepatic stellate cell proliferation. J. Cell. Physiol. 2011, 226, 2535–2542. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; He, Y.; Ma, T.T.; Huang, C.; Zhang, L.; Li, J. Participation of mir-200a in tgf-β1-mediated hepatic stellate cell activation. Mol. Cell. Biochem. 2014, 388, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.J.; Tao, H.; Liu, L.P.; Hu, W.; Deng, Z.Y.; Li, J. Mir-200a controls hepatic stellate cell activation and fibrosis via sirt1/notch1 signal pathway. Inflamm. Res. 2017, 66, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Sekiya, Y.; Ogawa, T.; Yoshizato, K.; Ikeda, K.; Kawada, N. Suppression of hepatic stellate cell activation by microrna-29b. Biochem. Biophys. Res. Commun. 2011, 412, 74–79. [Google Scholar] [CrossRef]
- Tu, X.; Zheng, X.; Li, H.; Cao, Z.; Chang, H.; Luan, S.; Zhu, J.; Chen, J.; Zang, Y.; Zhang, J. Microrna-30 protects against carbon tetrachloride-induced liver fibrosis by attenuating transforming growth factor beta signaling in hepatic stellate cells. Toxicol. Sci. 2015, 146, 157–169. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Yu, Y.; Li, S.; Liu, Y.; Zhou, S.; Cao, S.; Yin, J.; Li, G. Microrna-30a ameliorates hepatic fibrosis by inhibiting beclin1-mediated autophagy. J. Cell. Mol. Med. 2017, 21, 3679–3692. [Google Scholar] [CrossRef]
- Chen, C.; Wu, C.Q.; Zhang, Z.Q.; Yao, D.K.; Zhu, L. Loss of expression of mir-335 is implicated in hepatic stellate cell migration and activation. Exp. Cell Res. 2011, 317, 1714–1725. [Google Scholar] [CrossRef]
- Duan, B.; Hu, J.; Zhang, T.; Luo, X.; Zhou, Y.; Liu, S.; Zhu, L.; Wu, C.; Liu, W.; Chen, C.; et al. Mirna-338-3p/cdk4 signaling pathway suppressed hepatic stellate cell activation and proliferation. BMC Gastroenterol. 2017, 17, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feili, X.; Wu, S.; Ye, W.; Tu, J.; Lou, L. Microrna-34a-5p inhibits liver fibrosis by regulating tgf-β1/smad3 pathway in hepatic stellate cells. Cell Biol. Int. 2018, 42, 1370–1376. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Wang, W.; Yu, F.; Dong, P.; Chen, B.; Zhou, M.T. Microrna-30a suppresses the activation of hepatic stellate cells by inhibiting epithelial-to-mesenchymal transition. Cell. Physiol. Biochem. 2018, 46, 82–92. [Google Scholar] [CrossRef]
- Liang, Z.; Li, J.; Zhao, L.; Deng, Y. Mir-375 affects the hedgehog signaling pathway by downregulating rac1 to inhibit hepatic stellate cell viability and epithelial-mesenchymal transition. Mol. Med. Rep. 2021, 23, 182. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Yang, J.; Huang, K.; Pan, X.; Chen, B.; Dong, P.; Zheng, J. The epigenetically-regulated microrna-378a targets tgf-β2 in tgf-β1-treated hepatic stellate cells. Cell. Physiol. Biochem. 2016, 40, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Hyun, J.; Wang, S.; Kim, J.; Rao, K.M.; Park, S.Y.; Chung, I.; Ha, C.S.; Kim, S.W.; Yun, Y.H.; Jung, Y. Microrna-378 limits activation of hepatic stellate cells and liver fibrosis by suppressing gli3 expression. Nat. Commun. 2016, 7, 10993. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.; Fan, X.; Chen, B.; Dong, P.; Zheng, J. Activation of hepatic stellate cells is inhibited by microrna-378a-3p via wnt10a. Cell. Physiol. Biochem. 2016, 39, 2409–2420. [Google Scholar] [CrossRef]
- Wang, Y.Z.; Zhang, W.; Wang, Y.H.; Fu, X.L.; Xue, C.Q. Repression of liver cirrhosis achieved by inhibitory effect of mir-454 on hepatic stellate cells activation and proliferation via wnt10a. J. Biochem. 2019, 165, 361–367. [Google Scholar] [CrossRef]
- Wei, S.; Wang, Q.; Zhou, H.; Qiu, J.; Li, C.; Shi, C.; Zhou, S.; Liu, R.; Lu, L. Mir-455-3p alleviates hepatic stellate cell activation and liver fibrosis by suppressing hsf1 expression. Mol. Ther. Nucleic Acids 2019, 16, 758–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Zhang, H.; Li, L.; Yu, L.; Fu, L. Microrna-9 limits hepatic fibrosis by suppressing the activation and proliferation of hepatic stellate cells by directly targeting mrp1/abcc1. Oncol. Rep. 2017, 37, 1698–1706. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.; Chen, B.; Fan, X.; Li, G.; Dong, P.; Zheng, J. Epigenetically-regulated microrna-9-5p suppresses the activation of hepatic stellate cells via tgfbr1 and tgfbr2. Cell. Physiol. Biochem. 2017, 43, 2242–2252. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Wei, S.; Zhou, H.; Li, L.; Zhou, S.; Shi, C.; Shi, Y.; Qiu, J.; Lu, L. Microrna-98 inhibits hepatic stellate cell activation and attenuates liver fibrosis by regulating hlf expression. Front. Cell Dev. Biol. 2020, 8, 513. [Google Scholar] [CrossRef]
- Fabregat, I.; Caballero-Díaz, D. Transforming growth factor-β-induced cell plasticity in liver fibrosis and hepatocarcinogenesis. Front. Oncol. 2018, 8, 357. [Google Scholar] [CrossRef] [Green Version]
- Yan, X.; Liu, Z.; Chen, Y. Regulation of tgf-beta signaling by smad7. Acta Biochim. Biophys. Sin. 2009, 41, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Yan, X.; Liao, H.; Cheng, M.; Shi, X.; Lin, X.; Feng, X.-H.; Chen, Y.-G. Smad7 protein interacts with receptor-regulated smads (r-smads) to inhibit transforming growth factor-β (tgf-β)/smad signaling. J. Biol. Chem. 2016, 291, 382–392. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, H.; Abdollah, S.; Qiu, Y.; Cai, J.; Xu, Y.Y.; Grinnell, B.W.; Richardson, M.A.; Topper, J.N.; Gimbrone, M.A.; Wrana, J.L.; et al. The mad-related protein smad7 associates with the tgfbeta receptor and functions as an antagonist of tgfbeta signaling. Cell 1997, 89, 1165–1173. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhang, X.; Xie, F.; Zhang, Z.; van Dam, H.; Zhang, L.; Zhou, F. The regulation of tgf-β/smad signaling by protein deubiquitination. Protein Cell 2014, 5, 503–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.; Peng, Z.; Tang, H.; Xie, D.; Jia, Z.; Zhong, L.; Zhao, S.; Ma, Z.; Gao, Y.; Zeng, L.; et al. Loss of klf4 and consequential downregulation of smad7 exacerbate oncogenic tgf-β signaling in and promote progression of hepatocellular carcinoma. Oncogene 2017, 36, 2957–2968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, L.; Vo-Ransdell, C.; Abel, B.; Willoughby, C.; Jang, S.; Sowa, G. Caveolin-2 is a negative regulator of anti-proliferative function and signaling of transforming growth factor-β in endothelial cells. Am. J. Physiol. Cell Physiol. 2011, 301, C1161–C1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corpechot, C.; Barbu, V.; Wendum, D.; Kinnman, N.; Rey, C.; Poupon, R.; Housset, C.; Rosmorduc, O. Hypoxia-induced vegf and collagen i expressions are associated with angiogenesis and fibrogenesis in experimental cirrhosis. Hepatology 2002, 35, 1010–1021. [Google Scholar] [CrossRef]
- Mingyuan, X.; Qianqian, P.; Shengquan, X.; Chenyi, Y.; Rui, L.; Yichen, S.; Jinghong, X. Hypoxia-inducible factor-1α activates transforming growth factor-β1/smad s.signaling and increases collagen deposition in dermal fibroblasts. Oncotarget 2017, 9, 3188–3197. [Google Scholar] [CrossRef] [Green Version]
- Cai, C.X.; Buddha, H.; Castelino-Prabhu, S.; Zhang, Z.; Britton, R.S.; Bacon, B.R.; Neuschwander-Tetri, B.A. Activation of insulin-pi3k/akt-p70s6k pathway in hepatic stellate cells contributes to fibrosis in nonalcoholic steatohepatitis. Dig. Dis. Sci. 2017, 62, 968–978. [Google Scholar] [CrossRef] [PubMed]
- Reif, S.; Lang, A.; Lindquist, J.N.; Yata, Y.; Gabele, E.; Scanga, A.; Brenner, D.A.; Rippe, R.A. The role of focal adhesion kinase-phosphatidylinositol 3-kinase-akt signaling in hepatic stellate cell proliferation and type i collagen expression. J. Biol. Chem. 2003, 278, 8083–8090. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, S.; Kobayashi, M.; Kitagishi, Y. Roles for pi3k/akt/pten pathway in cell signaling of nonalcoholic fatty liver disease. ISRN Endocrinol. 2013, 2013, 472432. [Google Scholar] [CrossRef] [PubMed]
- Foglia, B.; Cannito, S.; Bocca, C.; Parola, M.; Novo, E. Erk pathway in activated, myofibroblast-like, hepatic stellate cells: A critical signaling crossroad sustaining liver fibrosis. Int. J. Mol. Sci. 2019, 20, 2700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ying, H.Z.; Chen, Q.; Zhang, W.Y.; Zhang, H.H.; Ma, Y.; Zhang, S.Z.; Fang, J.; Yu, C.H. Pdgf signaling pathway in hepatic fibrosis pathogenesis and therapeutics (review). Mol. Med. Rep. 2017, 16, 7879–7889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandramouli, S.; Yu, C.Y.; Yusoff, P.; Lao, D.H.; Leong, H.F.; Mizuno, K.; Guy, G.R. Tesk1 interacts with spry2 to abrogate its inhibition of erk phosphorylation downstream of receptor tyrosine kinase signaling. J. Biol. Chem. 2008, 283, 1679–1691. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; Guo, M.-G.; Lou, X.-L.; Li, X.-Y.; Xu, Y.; Ji, W.-D.; Huang, X.-D.; Yang, J.-H.; Duan, J.-C. Hepatocyte nuclear factor 4α induces a tendency of differentiation and activation of rat hepatic stellate cells. World J. Gastroenterol. 2015, 21, 5856–5866. [Google Scholar] [CrossRef]
- Yang, T.; Poenisch, M.; Khanal, R.; Hu, Q.; Dai, Z.; Li, R.; Song, G.; Yuan, Q.; Yao, Q.; Shen, X.; et al. Therapeutic hnf4a mrna attenuates liver fibrosis in a preclinical model. J. Hepatol. 2021, 75, 1420–1433. [Google Scholar] [CrossRef]
- Monga, S.P. Β-catenin signaling and roles in liver homeostasis, injury, and tumorigenesis. Gastroenterology 2015, 148, 1294–1310. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Lamouille, S.; Derynck, R. Tgf-β-induced epithelial to mesenchymal transition. Cell Res. 2009, 19, 156–172. [Google Scholar] [CrossRef]
- Pelullo, M.; Zema, S.; Nardozza, F.; Checquolo, S.; Screpanti, I.; Bellavia, D. Wnt, notch, and tgf-β pathways impinge on hedgehog signaling complexity: An open window on cancer. Front. Genet. 2019, 10, 711. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Leng, X.-S.; Zhu, J.-Y.; Wang, G. Suppression of hedgehog signaling regulates hepatic stellate cell activation and collagen secretion. Int. J. Clin. Exp. Pathol. 2015, 8, 14574–14579. [Google Scholar]
- Tsagakis, I.; Douka, K.; Birds, I.; Aspden, J.L. Long non-coding rnas in development and disease: Conservation to mechanisms. J. Pathol. 2020, 250, 480–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhanoa, J.K.; Sethi, R.S.; Verma, R.; Arora, J.S.; Mukhopadhyay, C.S. Long non-coding rna: Its evolutionary relics and biological implications in mammals: A review. J. Anim. Sci. Technol. 2018, 60, 25. [Google Scholar] [CrossRef] [Green Version]
- Derrien, T.; Johnson, R.; Bussotti, G.; Tanzer, A.; Djebali, S.; Tilgner, H.; Guernec, G.; Martin, D.; Merkel, A.; Knowles, D.G.; et al. The gencode v7 catalog of human long noncoding rnas: Analysis of their gene structure, evolution, and expression. Genome Res. 2012, 22, 1775–1789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Wang, Y.; Xie, S.; Liu, Y.; Xie, Z. Global and cell-type specific properties of lincrnas with ribosome occupancy. Nucleic Acids Res. 2017, 45, 2786–2796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Zhang, Y.H.; Pan, X.; Liu, M.; Wang, S.; Huang, T.; Cai, Y.D. Tissue expression difference between mrnas and lncrnas. Int. J. Mol. Sci. 2018, 19, 3416. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Sun, Z. Novel lincrna discovery and tissue-specific gene expression across 30 normal human tissues. Genes 2021, 12, 614. [Google Scholar] [CrossRef]
- Moore, J.B.T.; Uchida, S. Functional characterization of long noncoding rnas. Curr. Opin. Cardiol. 2020, 35, 199–206. [Google Scholar] [CrossRef]
- Zhao, Y.; Wu, J.; Liangpunsakul, S.; Wang, L. Long non-coding rna in liver metabolism and disease: Current status. Liver Res. 2017, 1, 163–167. [Google Scholar] [CrossRef]
- Peng, H.; Wan, L.-Y.; Liang, J.-J.; Zhang, Y.-Q.; Ai, W.-B.; Wu, J.-F. The roles of lncrna in hepatic fibrosis. Cell Biosci. 2018, 8, 63. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Huang, S.; Zheng, X.; Gu, S.; Xu, Q.; Gong, Y.; Zhang, J.; Fu, B.; Tang, L. Regulatory long non-coding rnas of hepatic stellate cells in liver fibrosis (review). Exp. Ther. Med. 2021, 21, 351. [Google Scholar] [CrossRef]
- Zhang, K.; Han, X.; Zhang, Z.; Zheng, L.; Hu, Z.; Yao, Q.; Cui, H.; Shu, G.; Si, M.; Li, C.; et al. The liver-enriched lnc-lfar1 promotes liver fibrosis by activating tgfβ and notch pathways. Nat. Commun. 2017, 8, 144. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Luo, Z.; Pan, Y.; Zheng, W.; Li, W.; Zhang, Z.; Xiong, P.; Xu, D.; Du, M.; Wang, B.; et al. H19/mir-148a/usp4 axis facilitates liver fibrosis by enhancing tgf-β signaling in both hepatic stellate cells and hepatocytes. J. Cell. Physiol. 2019, 234, 9698–9710. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhou, F.; Drabsch, Y.; Gao, R.; Snaar-Jagalska, B.E.; Mickanin, C.; Huang, H.; Sheppard, K.A.; Porter, J.A.; Lu, C.X.; et al. Usp4 is regulated by akt phosphorylation and directly deubiquitylates tgf-β type i receptor. Nat. Cell Biol. 2012, 14, 717–726. [Google Scholar] [CrossRef]
- Li, Z.; Wang, J.; Zeng, Q.; Hu, C.; Zhang, J.; Wang, H.; Yan, J.; Li, H.; Yu, Z. Long noncoding rna hottip promotes mouse hepatic stellate cell activation via downregulating mir-148a. Cell Physiol. Biochem. 2018, 51, 2814–2828. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Mao, Y.; Dong, P.; Huang, Z.; Yu, F. Long noncoding rna hottip mediates srf expression through sponging mir-150 in hepatic stellate cells. J. Cell. Mol. Med. 2019, 23, 1572–1580. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Liu, X.; Zhou, Q.; Huang, C.; Meng, X.; Xu, F.; Li, J. Silent information regulator 1 (sirt1) ameliorates liver fibrosis via promoting activated stellate cell apoptosis and reversion. Toxicol. Appl. Pharmacol. 2015, 289, 163–176. [Google Scholar] [CrossRef]
- Liao, J.; Zhang, Z.; Yuan, Q.; Liu, Q.; Kuang, J.; Fang, Y.; Hu, X. A lncrna gpr137b-ps/mir-200a-3p/cxcl14 axis modulates hepatic stellate cell (hsc) activation. Toxicol. Lett. 2021, 336, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Chen, B.; Dong, P.; Zheng, J. Hotair epigenetically modulates pten expression via microrna-29b: A novel mechanism in regulation of liver fibrosis. Mol. Ther. 2017, 25, 205–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bian, E.B.; Wang, Y.Y.; Yang, Y.; Wu, B.M.; Xu, T.; Meng, X.M.; Huang, C.; Zhang, L.; Lv, X.W.; Xiong, Z.G.; et al. Hotair facilitates hepatic stellate cells activation and fibrogenesis in the liver. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 674–686. [Google Scholar] [CrossRef]
- Wu, J.-C.; Luo, S.-Z.; Liu, T.; Lu, L.-G.; Xu, M.-Y. Linc-scrg1 accelerates liver fibrosis by decreasing rna-binding protein tristetraprolin. FASEB J. 2019, 33, 2105–2115. [Google Scholar] [CrossRef] [Green Version]
- Fu, N.; Zhao, S.X.; Kong, L.B.; Du, J.H.; Ren, W.G.; Han, F.; Zhang, Q.S.; Li, W.C.; Cui, P.; Wang, R.Q.; et al. Lncrna-atb/microrna-200a/β-catenin regulatory axis involved in the progression of hcv-related hepatic fibrosis. Gene 2017, 618, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Lu, Z.; Cai, J.; Huang, K.; Chen, B.; Li, G.; Dong, P.; Zheng, J. Malat1 functions as a competing endogenous rna to mediate rac1 expression by sequestering mir-101b in liver fibrosis. Cell Cycle 2015, 14, 3885–3896. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Lin, H.; Chen, X.; Li, G.; Zhao, Y.; Zheng, L.; Shi, Z.; Zhang, K.; Hong, W.; Han, T.; et al. Lncrna meg8 suppresses activation of hepatic stellate cells and epithelial-mesenchymal transition of hepatocytes via the notch pathway. Biochem. Biophys. Res. Commun. 2020, 521, 921–927. [Google Scholar] [CrossRef]
- Yu, F.; Jiang, Z.; Chen, B.; Dong, P.; Zheng, J. Neat1 accelerates the progression of liver fibrosis via regulation of microrna-122 and kruppel-like factor 6. J. Mol. Med. 2017, 95, 1191–1202. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Huang, F.; Zhang, R.; Luo, H. Lncrna neat1 expedites the progression of liver fibrosis in mice through targeting mir-148a-3p and mir-22-3p to upregulate cyth3. Cell Cycle 2021, 20, 490–507. [Google Scholar] [CrossRef]
- Wang, Q.; Wei, S.; Li, L.; Bu, Q.; Zhou, H.; Su, W.; Liu, Z.; Wang, M.; Lu, L. Mir-139-5p sponged by lncrna neat1 regulates liver fibrosis via targeting β-catenin/sox9/tgf-β1 pathway. Cell Death Discov. 2021, 7, 243. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Yu, F.; Dong, P.; Wu, L.; Zhang, Y.; Hu, Y.; Zheng, L. Long non-coding rna pvt1 activates hepatic stellate cells through competitively binding microrna-152. Oncotarget 2016, 7, 62886–62897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, F.; Lu, Z.; Chen, B.; Wu, X.; Dong, P.; Zheng, J. Salvianolic acid b-induced microrna-152 inhibits liver fibrosis by attenuating dnmt1-mediated patched1 methylation. J. Cell. Mol. Med. 2015, 19, 2617–2632. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Han, Y.; Hu, Z.; Zhang, Z.; Shao, S.; Yao, Q.; Zheng, L.; Wang, J.; Han, X.; Zhang, Y.; et al. Scarna10, a nuclear-retained long non-coding rna, promotes liver fibrosis and serves as a potential biomarker. Theranostics 2019, 9, 3622–3638. [Google Scholar] [CrossRef]
- Xie, Z.; Wu, Y.; Liu, S.; Lai, Y.; Tang, S. Lncrna-snhg7/mir-29b/dnmt3a axis affects activation, autophagy and proliferation of hepatic stellate cells in liver fibrosis. Clin. Res. Hepatol. Gastroenterol. 2021, 45, 101469. [Google Scholar] [CrossRef]
- Yu, F.; Dong, P.; Mao, Y.; Zhao, B.; Huang, Z.; Zheng, J. Loss of lncrna-snhg7 promotes the suppression of hepatic stellate cell activation via mir-378a-3p and dvl2. Mol. Ther. Nucleic Acids 2019, 17, 235–244. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Hong, Y.; Zhang, K. Tug1 is involved in liver fibrosis and activation of hscs by regulating mir-29b. Biochem. Biophys. Res. Commun. 2018, 503, 1394–1400. [Google Scholar] [CrossRef]
- Xie, Z.Y.; Wang, F.F.; Xiao, Z.H.; Liu, S.F.; Lai, Y.L.; Tang, S.L. Long noncoding rna xist enhances ethanol-induced hepatic stellate cells autophagy and activation via mir.r.r.r-29b/hmgb1 axis. IUBMB Life 2019, 71, 1962–1972. [Google Scholar] [CrossRef]
- Yu, F.; Zheng, J.; Mao, Y.; Dong, P.; Lu, Z.; Li, G.; Guo, C.; Liu, Z.; Fan, X. Long non-coding rna growth arrest-specific transcript 5 (gas5) inhibits liver fibrogenesis through a mechanism of competing endogenous rna. J. Biol. Chem. 2015, 290, 28286–28298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.-Q.; Xu, M.-Y.; Qu, Y.; Hu, J.-J.; Li, Z.-H.; Zhang, Q.-D.; Lu, L.-G. Tet3 mediates the activation of human hepatic stellate cells via modulating the expression of long non-coding rna hif1a-as1. Int. J. Clin. Exp. Pathol. 2014, 7, 7744–7751. [Google Scholar] [PubMed]
- Yu, F.; Lu, Z.; Chen, B.; Dong, P.; Zheng, J. Identification of a novel lincrna-p21-mir-181b-pten signaling cascade in liver fibrosis. Mediat. Inflamm. 2016, 2016, 9856538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, F.; Guo, Y.; Chen, B.; Shi, L.; Dong, P.; Zhou, M.; Zheng, J. Lincrna-p21 inhibits the wnt/β-catenin pathway in activated hepatic stellate cells via sponging microrna-17-5p. Cell. Physiol. Biochem. 2017, 41, 1970–1980. [Google Scholar] [CrossRef]
- Dong, Z.; Li, S.; Si, L.; Ma, R.; Bao, L.; Bo, A. Identification lncrna loc102551149/mir-23a-5p pathway in hepatic fibrosis. Eur. J. Clin. Investig. 2020, 50, e13243. [Google Scholar] [CrossRef]
- He, Y.; Wu, Y.; Huang, C.; Meng, X.M.; Ma, T.; Wu, B.M.; Xu, F.; Zhang, L.; Lv, X.W.; Li, J. Inhibitory effects of long noncoding rna meg3 on hepatic stellate cells activation and liver fibrogenesis. Biochim. Biophys. Acta 2014, 1842, 2204–2215. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.; Geng, W.; Dong, P.; Huang, Z.; Zheng, J. Lncrna-meg3 inhibits activation of hepatic stellate cells through smo protein and mir-212. Cell Death Dis. 2018, 9, 1014. [Google Scholar] [CrossRef]
- Yu, F.; Lu, Z.; Huang, K.; Wang, X.; Xu, Z.; Chen, B.; Dong, P.; Zheng, J. Microrna-17-5p-activated wnt/β-catenin pathway contributes to the progression of liver fibrosis. Oncotarget 2016, 7, 81–93. [Google Scholar] [CrossRef] [Green Version]
- Machado, M.V.; Diehl, A.M. Hedgehog signalling in liver pathophysiology. J. Hepatol. 2018, 68, 550–562. [Google Scholar] [CrossRef] [Green Version]
- Patop, I.L.; Wüst, S.; Kadener, S. Past, present, and future of circrnas. Embo J. 2019, 38, e100836. [Google Scholar] [CrossRef] [PubMed]
- Santer, L.; Bär, C.; Thum, T. Circular rnas: A novel class of functional rna molecules with a therapeutic perspective. Mol. Ther. 2019, 27, 1350–1363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu, N.; Jamal, R. Circular rnas as promising biomarkers: A mini-review. Front. Physiol. 2016, 7, 355. [Google Scholar] [CrossRef]
- Ragan, C.; Goodall, G.J.; Shirokikh, N.E.; Preiss, T. Insights into the biogenesis and potential functions of exonic circular rna. Sci. Rep. 2019, 9, 2048. [Google Scholar] [CrossRef]
- Sulaiman, S.A.; Abdul Murad, N.A.; Mohamad Hanif, E.A.; Abu, N.; Jamal, R. Prospective advances in circular rna investigation. Adv. Exp. Med. Biol. 2018, 1087, 357–370. [Google Scholar] [PubMed]
- Zeng, X.; Yuan, X.; Cai, Q.; Tang, C.; Gao, J. Circular rna as an epigenetic regulator in chronic liver diseases. Cells 2021, 10, 1945. [Google Scholar] [CrossRef]
- Fu, L.Y.; Wang, S.W.; Hu, M.Y.; Jiang, Z.L.; Shen, L.L.; Zhou, Y.P.; Guo, J.M.; Hu, Y.R. Circular rnas in liver diseases: Mechanisms and therapeutic targets. Life Sci. 2021, 264, 118707. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Xia, L.; Sun, M.; Yang, C.; Wang, F. Circular rna in liver: Health and diseases. Adv. Exp. Med. Biol. 2018, 1087, 245–257. [Google Scholar]
- Zhu, L.; Ren, T.; Zhu, Z.; Cheng, M.; Mou, Q.; Mu, M.; Liu, Y.; Yao, Y.; Cheng, Y.; Zhang, B.; et al. Thymosin-β4 mediates hepatic stellate cell activation by interfering with circrna-0067835/mir-155/foxo3 signaling pathway. Cell. Physiol. Biochem. 2018, 51, 1389–1398. [Google Scholar] [CrossRef]
- Chen, Y.; Yuan, B.; Wu, Z.; Dong, Y.; Zhang, L.; Zeng, Z. Microarray profiling of circular rnas and the potential regulatory role of hsa_circ_0071410 in the activated human hepatic stellate cell induced by irradiation. Gene 2017, 629, 35–42. [Google Scholar] [CrossRef]
- Liu, W.; Feng, R.; Li, X.; Li, D.; Zhai, W. Tgf-β- and lipopolysaccharide-induced upregulation of circular rna pwwp2a promotes hepatic fibrosis via sponging mir-203 and mir-223. Aging 2019, 11, 9569–9580. [Google Scholar] [CrossRef]
- Chen, Y.; Yuan, B.; Chen, G.; Zhang, L.; Zhuang, Y.; Niu, H.; Zeng, Z. Circular rna rsf1 promotes inflammatory and fibrotic phenotypes of irradiated hepatic stellate cell by modulating mir-146a-5p. J. Cell. Physiol. 2020, 235, 8270–8282. [Google Scholar] [CrossRef] [PubMed]
- Niu, H.; Zhang, L.; Chen, Y.H.; Yuan, B.Y.; Wu, Z.F.; Cheng, J.C.; Lin, Q.; Zeng, Z.C. Circular rna tubd1 acts as the mir-146a-5p sponge to affect the viability and pro-inflammatory cytokine production of lx-2 cells through the tlr4 pathway. Radiat. Res. 2020, 193, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Chen, X.; Wang, J.N.; Xu, J.J.; Wang, A.; Li, J.J.; Wu, S.; Wu, Y.Y.; Li, X.F.; Huang, C.; et al. Circular rna circube2k promotes hepatic fibrosis via sponging mir-149-5p/tgf-β2 axis. FASEB J. 2021, 35, e21622. [Google Scholar] [CrossRef]
- Li, S.; Song, F.; Lei, X.; Li, J.; Li, F.; Tan, H. Hsa_circ_0004018 suppresses the progression of liver fibrosis through regulating the hsa-mir-660-3p/tep1 axis. Aging 2020, 12, 11517–11529. [Google Scholar] [CrossRef]
- Jin, H.; Li, C.; Dong, P.; Huang, J.; Yu, J.; Zheng, J. Circular rna cmto1 promotes pten expression through sponging mir-181b-5p in liver fibrosis. Front. Cell Dev. Biol. 2020, 8, 714. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Dong, R.; Guo, Y.; He, J.; Shao, C.; Yi, P.; Yu, F.; Gu, D.; Zheng, J. Circmto1 inhibits liver fibrosis via regulation of mir-17-5p and smad7. J. Cell. Mol. Med. 2019, 23, 5486–5496. [Google Scholar] [CrossRef] [Green Version]
- Ji, D.; Chen, G.F.; Wang, J.C.; Ji, S.H.; Wu, X.W.; Lu, X.J.; Chen, J.L.; Li, J.T. Hsa_circ_0070963 inhibits liver fibrosis via regulation of mir-223-3p and lemd3. Aging 2020, 12, 1643–1655. [Google Scholar] [CrossRef]
- Chen, X.; Li, H.D.; Bu, F.T.; Li, X.F.; Chen, Y.; Zhu, S.; Wang, J.N.; Chen, S.Y.; Sun, Y.Y.; Pan, X.Y.; et al. Circular rna circfbxw4 suppresses hepatic fibrosis via targeting the mir-18b-3p/fbxw7 axis. Theranostics 2020, 10, 4851–4870. [Google Scholar] [CrossRef]
- Bu, F.T.; Zhu, Y.; Chen, X.; Wang, A.; Zhang, Y.F.; You, H.M.; Yang, Y.; Yang, Y.R.; Huang, C.; Li, J. Circular rna circpsd3 alleviates hepatic fibrogenesis by regulating the mir-92b-3p/smad7 axis. Mol. Ther. Nucleic Acids 2021, 23, 847–862. [Google Scholar] [CrossRef] [PubMed]
- Fatima, A.; Irmak, D.; Noormohammadi, A.; Rinschen, M.M.; Das, A.; Leidecker, O.; Schindler, C.; Sánchez-Gaya, V.; Wagle, P.; Pokrzywa, W.; et al. The ubiquitin-conju.ugating enzyme ube2k determines neurogenic potential through histone h3 in human embryonic stem cells. Commun. Biol. 2020, 3, 262. [Google Scholar] [CrossRef] [PubMed]
- Shang, H.; Liu, X.; Guo, H. Knockdown of fstl1 attenuates hepatic stellate cell activation through the tgf-β1/smad3 signaling pathway. Mol. Med. Rep. 2017, 16, 7119–7123. [Google Scholar] [CrossRef] [Green Version]
- Seki, E.; De Minicis, S.; Österreicher, C.H.; Kluwe, J.; Osawa, Y.; Brenner, D.A.; Schwabe, R.F. Tlr4 enhances tgf-β signaling and hepatic fibrosis. Nat. Med. 2007, 13, 1324–1332. [Google Scholar] [CrossRef] [PubMed]
- Chambers, D.M.; Moretti, L.; Zhang, J.J.; Cooper, S.W.; Chambers, D.M.; Santangelo, P.J.; Barker, T.H. Lem domain-containing protein 3 antagonizes tgfβ-smad2/3 signaling in a stiffness-dependent manner in both the nucleus and cytosol. J. Biol. Chem. 2018, 293, 15867–15886. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.; Wang, G.; Zhang, J.; Cao, W.; Chen, X. Resveratrol decreases foxo protein expression through pi3k-akt-dependent pathway inhibition in H2O2-treated synoviocytes. Histol. Histopathol. 2017, 32, 1305–1315. [Google Scholar]
- Li, L.; Yang, J.; Li, F.; Zhu, L.; Hao, J.; Gao, F. Fbxw7 mediates high glucose-induced srebp-1 expression in renal tubular cells of diabetic nephropathy under pi3k/akt pathway regulation. Mol. Med. Rep. 2021, 23, 233. [Google Scholar] [CrossRef]
- Sundqvist, A.; Bengoechea-Alonso, M.T.; Ye, X.; Lukiyanchuk, V.; Jin, J.; Harper, J.W.; Ericsson, J. Control of lipid metabolism by phosphorylation-dependent degradation of the srebp family of transcription factors by scf(fbw7). Cell Metab. 2005, 1, 379–391. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Guo, M.; Li, Y.; Shen, M.; Kong, D.; Shao, J.; Ding, H.; Tan, S.; Chen, A.; Zhang, F.; et al. Rna-binding protein zfp36/ttp protects against ferroptosis by regulating autophagy signaling pathway in hepatic stellate cells. Autophagy 2020, 16, 1482–1505. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, L.; Wang, Y.; Zhang, M.; Li, L.; Zhu, D.; Li, X.; Gu, H.; Zhang, C.Y.; Zen, K. Protective role of estrogen-induced mirna-29 expression in carbon tetrachloride-induced mouse liver injury. J. Biol. Chem. 2012, 287, 14851–14862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; He, Y.; Mackowiak, B.; Gao, B. Micrornas as regulators, biomarkers and therapeutic targets in liver diseases. Gut 2021, 70, 784–795. [Google Scholar] [CrossRef] [PubMed]
- Janssen, H.L.; Reesink, H.W.; Lawitz, E.J.; Zeuzem, S.; Rodriguez-Torres, M.; Patel, K.; van der Meer, A.J.; Patick, A.K.; Chen, A.; Zhou, Y.; et al. Treatment of hcv infection by targeting microrna. N. Engl. J. Med. 2013, 368, 1685–1694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ottosen, S.; Parsley, T.B.; Yang, L.; Zeh, K.; van Doorn, L.J.; van der Veer, E.; Raney, A.K.; Hodges, M.R.; Patick, A.K. In vitro antiviral activity and preclinical and clinical resistance profile of miravirsen, a novel anti-hepatitis c virus therapeutic targeting the human factor mir-122. Antimicrob. Agents Chemother. 2015, 59, 599–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, S.H.; Yu, B.; Wang, X.; Lu, Y.; Schmidt, C.R.; Lee, R.J.; Lee, L.J.; Jacob, S.T.; Ghoshal, K. Cationic lipid nanoparticles for therapeutic delivery of sirna and mirna to murine liver tumor. Nanomedicine 2013, 9, 1169–1180. [Google Scholar] [CrossRef] [Green Version]
- He, S.; Guo, W.; Deng, F.; Chen, K.; Jiang, Y.; Dong, M.; Peng, L.; Chen, X. Targeted delivery of microrna 146b mimic to hepatocytes by lactosylated pdmaema nanoparticles for the treatment of nafld. Artif. Cells Nanomed. Biotechnol. 2018, 46, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Newman, L.A.; Sorich, M.J.; Rowland, A. Role of extracellular vesicles in the pathophysiology, diagnosis and tracking of non-alcoholic fatty liver disease. J. Clin. Med. 2020, 9, 2032. [Google Scholar] [CrossRef]
- Dorairaj, V.; Sulaiman, S.A.; Abu, N.; Abdul Murad, N.A. Extracellular vesicles in the development of the non-alcoholic fatty liver disease: An update. Biomolecules 2020, 10, 1494. [Google Scholar] [CrossRef]
- Li, X.; Chen, R.; Kemper, S.; Brigstock, D.R. Extracellular vesicles from hepatocytes are therapeutic for toxin-mediated fibrosis and gene expression in the liver. Front. Cell Dev. Biol. 2019, 7, 368. [Google Scholar] [CrossRef]
- Li, X.; Chen, R.; Kemper, S.; Brigstock, D.R. Structural and functional characterization of fibronectin in extracellular vesicles from hepatocytes. Front. Cell Dev. Biol. 2021, 9, 640667. [Google Scholar] [CrossRef] [PubMed]
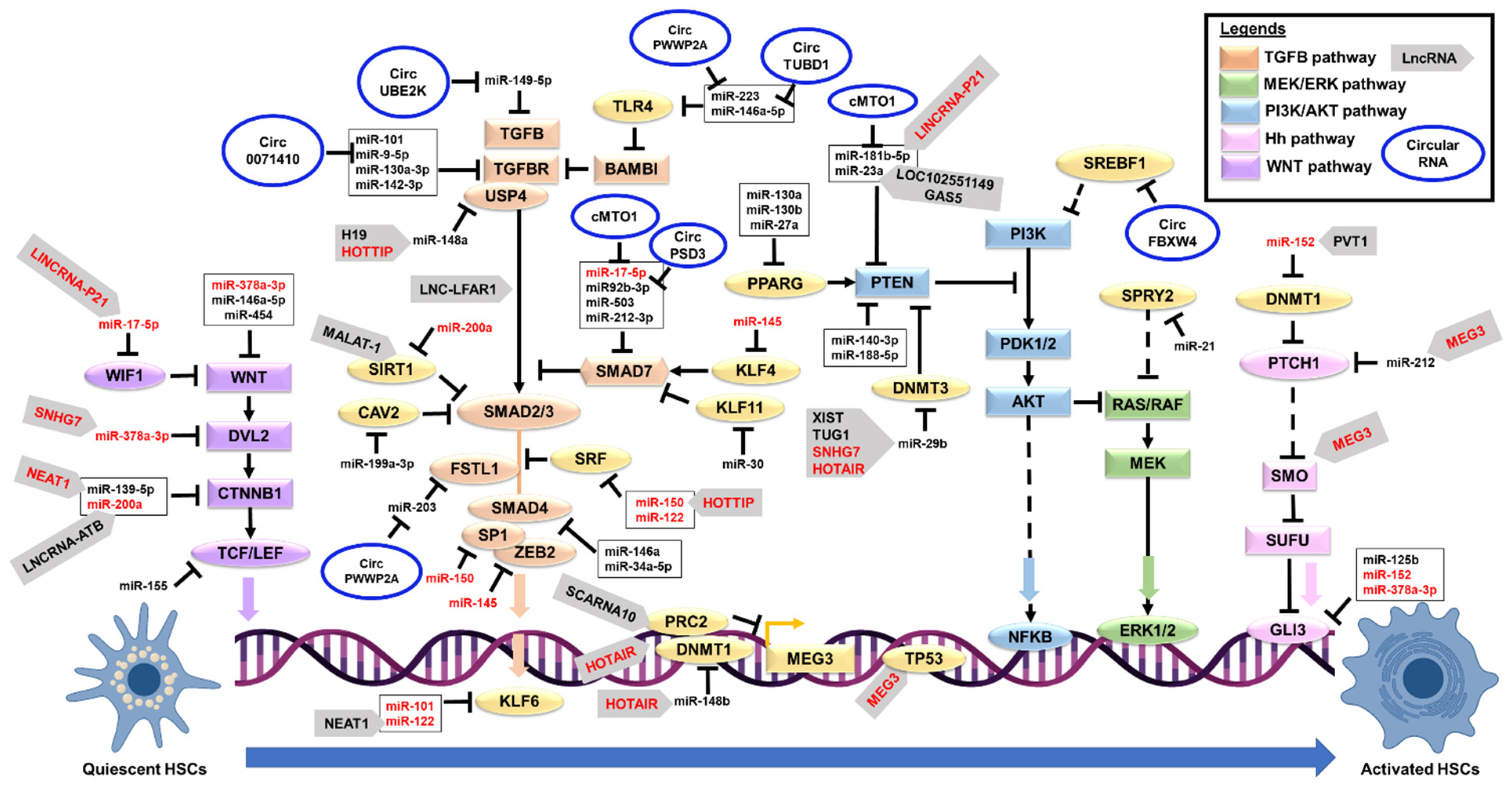
| MicroRNA | Expression | Target Gene | Disease Model | Function | Role in Fibrosis | References |
|---|---|---|---|---|---|---|
| miR-125a-5p | Up | Hif1a | Mouse CCI(4)-treated, Mouse HSC cell line | Activates HSCs proliferation | Profibrotic | [22] |
| miR-126 | Up | NFKBIA | Human LX-2 cell line | Activates HSCs proliferation | Profibrotic | [23] |
| miR-130a | Up | Pparg | Rat CCI(4)-treated, Rat HSC-T6 cell line | Activates HSCs proliferation | Profibrotic | [24] |
| miR-130b | Up | Pparg | Rat CCI(4)-treated, Rat HSC-T6 cell line | Activates HSCs proliferation | Profibrotic | [24] |
| miR-130b-5p | Up | SIRT4 | Patient samples, Rat HSC-T6 cell line | Activates HSCs proliferation | Profibrotic | [25] |
| miR-140-3p | Up | Pten | Rat HSC-T6 cell line | Activates HSCs proliferation | Profibrotic | [26] |
| miR-141 | Up | PTEN | Patient samples, Human LX-2 cell line, Rat HSC-T6 cell line | Activates HSCs proliferation | Profibrotic | [27] |
| miR-145 | Up | KLF4 | Patient samples, Rat CCI(4)-treated, Human LX-2 cell line, Rat HSC cell line | Activates HSCs proliferation | Profibrotic | [28] |
| miR-17-5p | Up | Smad7 | Patient samples, Rat CCI(4)-treated, Rat HSC cell line | Activates HSCs proliferation upon TGFB1 treatment | Profibrotic | [29] |
| miR-181b | Up | Pten | Human LX-2 cell line, Rat CCI(4)-treated | Promotes the profibrotic factors (Acta2 and Col1a2) | Profibrotic | [30] |
| Up | Cdkn1b | Patient samples, Rat HSC-T6 cell line | Activates HSCs proliferation | Profibrotic | [31] | |
| miR-188-5p | Up | Pten | Patient samples, Mouse CCI(4)-treated, Rat CCI(4)-treated | Activates HSCs proliferation | Profibrotic | [32] |
| miR-199a-3p | Up | Cav2 | Mouse CCI(4)-treated, Rat CCI(4)-treated | HSC activation | Profibrotic | [33] |
| miR-21 | Up | SPRY2 and HNF4A | Patient samples, HEK-293 cell line, Rat HSC-T6 cell line, Hepatocyte cell line | HSC activation via ERK1 signaling | Profibrotic | [34] |
| miR-212-3p | Up | Smad7 | Human LX-2 cell line, HEK293T cell line, Mouse CCI(4)-treated | Activates HSCs proliferation upon TGFB1 treatment | Profibrotic | [35] |
| miR-214 | Up | SUFU | Human LX-2 cell line, Rat HSC cell line | HSC activation | Profibrotic | [36] |
| miR-221 | Up | Col1a1 | Patient samples, Mouse TAA-treated, Mouse MCDD-treated | HSC activation | Profibrotic | [37] |
| Up | LAMP2 | Human LX-2 cell line, Mouse CCI(4)-treated | HSC activation | Profibrotic | [38] | |
| miR-23a | Up | Pten | Rat CCI(4)-treated | HSC activation | Profibrotic | [39] |
| miR-27a | Up | PPARG | Patient samples, Human LX-2 and HepG2 cell lines, Rat DMN-treated | Activates HSC proliferation and profibrotic factors | Profibrotic | [40] |
| miR-503 | Up | Smad7 | Human LX-2 cell line, Rat CCI(4)-treated | Activates HSC proliferation | Profibrotic | [41] |
| miR-708 | Up | TMEM88 | Patient samples, Human LX-2 cell line | Activates HSC proliferation via WNT signaling | Profibrotic | [42] |
| Up | Zeb1 | Human LX-2 cell line, Mouse CCI(4)-treated | Activates HSC proliferation via WNT signaling | Profibrotic | [43] | |
| miR-942 | Up | BAMBI | Patient samples, Human LX-2 cell line | HSC activation | Profibrotic | [44] |
| miR-101 | Down | Tgfbr1 and Klf6 | Human 293A cell line, Mouse AML-12 cell line, Rat HSC-T6 cell line, Mouse CCI(4)-treated | Suppresses HSC activation and proliferation | Anti-fibrotic | [45] |
| miR-122 | Down | P4HA1 | Patient samples, Human LX-2 cell line, Mouse CCI(4)-treated, Rat HSC cell line | Suppresses HSC activation and collagen production | Anti-fibrotic | [46,47] |
| Down | SRF and FN1 | Patient samples, Human LX-2 and HEK293T cell lines, Mouse CCI(4)-treated | Suppresses HSC activation and collagen production | Anti-fibrotic | [48] | |
| miR-125b | Down | Gli3 | Patient samples, Human LX-2, AML12, LO2, LX2, THP-1 and 293T cell lines, Rat CCI(4)-treated | Suppresses HSC activation and ECM proteins | Anti-fibrotic | [49] |
| miR-126 * | Down | Vegfa | Rat CCI(4)-treated | Suppresses HSC activation and proliferation | Anti-fibrotic | [50] |
| miR-130a-3p | Down | Tgfbr1/Tgfbr2 | Patient samples, Mouse (NASH) | Suppresses HSC activation | Anti-fibrotic | [51] |
| miR-139-5p | Down | PMP22 | Human LX-2 cell line, Mouse CCI(4)-treated | Suppresses HSC activation | Anti-fibrotic | [52] |
| miR-142-3p | Down | Tgfbr1 | Patient samples, Rat HSC cell line | Suppresses HSC activation | Anti-fibrotic | [53] |
| miR-145 | Down | ZEB2 | Human LX-2 cell line, Mouse CCI(4)-treated, Rat HSC-T6 cell line | Suppresses HSC activation and proliferation | Anti-fibrotic | [54] |
| miR-146a | Down | Smad4 | Rat CCI(4)-treated, Rat HSC-T6 cell line | Suppresses HSC activation upon TGFB1 treatment | Anti-fibrotic | [55] |
| miR-146a-5p | Down | Wnt1 and Wnt5a | Mouse (NASH), Mouse primary HSC cells, Human LX-2 and Rat HSC-T6 cell lines | Suppresses HSC activation | Anti-fibrotic | [56] |
| Down | PTPRA | Human LX-2 cell line | Suppresses HSC activation | Anti-fibrotic | [57] | |
| miR-150 | Down | SP1 and COL4A4 | Human LX-2 cell line | Suppresses HSC proliferation and ECM proteins | Anti-fibrotic | [58] |
| miR-152 | Down | Gli3 | Patient samples, Rat CCI(4)-treated, HEK293T, LX-2, THP-1, AML12 and L02 cell lines | Suppresses HSC activation | Anti-fibrotic | [59] |
| miR-155 | Down | TCF4 and AGTR1 | Patient samples, Rat DMN-treated, Rat HSC cells, Rat HSC-T6 and Human HEK-293 cell lines | Suppresses HSC activation via ERK1 signaling | Anti-fibrotic | [60] |
| miR-193a/b-3p | Down | CAPRIN1 and TGFB2 | Mouse ConA-treated, Human LX-2 cell line | Suppresses HSC activation and proliferation | Anti-fibrotic | [61] |
| miR-194 | Down | AKT2 | Patient samples, Mouse CCI(4)-treated, Human LX-2 cell line | Suppresses HSC proliferation and collagen production | Anti-fibrotic | [62] |
| miR-195 | Down | CCNE1 | Human LX-2 cell line | Suppresses HSC proliferation upon IFNB1 treatment | Anti-fibrotic | [63] |
| miR-200a | Down | Ctnnb1 and Tgfb2 | Rat CCI(4)-treated, Rat HSC-T6 and Human HEK-293 cell lines | Suppresses HSC proliferation via TGFB and WNT/β-catenin pathways | Anti-fibrotic | [64] |
| Down | Sirt1 | Rat CCI(4)-treated, Rat HSC-T6 cell line | Suppresses HSC activation and proliferation | Anti-fibrotic | [65] | |
| miR-29b | Down | Col1a1 | Mouse HSC cells, Human LX-2 cell line | Suppresses HSC activation | Anti-fibrotic | [66] |
| miR-30 | Down | Klf11 | Mouse CCI(4)-treated, Rat HSC-T6 and HEK-293 cell lines | Suppresses HSC activation | Anti-fibrotic | [67] |
| miR-30a | Down | Becn1 | Mouse CCI(4)-treated, Rat HSC-T6 and LX-2 cell lines | Suppresses HSC activation and ECM proteins | Anti-fibrotic | [68] |
| miR-335 | Down | Tnc | Rat HSC cells | Suppresses HSC proliferation and collagen production | Anti-fibrotic | [69] |
| miR-338-3p | Down | Cdk4 | Rat HSC cells, Rat HSC-T6 and Human HEK-293 cell lines | Suppresses HSC activation and proliferation | Anti-fibrotic | [70] |
| miR-34a-5p | Down | Smad4 | Patient samples, Mouse CCI(4)-treated, Human LX-2 cell line | Suppresses HSC activation | Anti-fibrotic | [71] |
| Down | Snai1 | Patient samples, Rat CCI(4)-treated, HEK-293 cell line | Suppresses HSC proliferation and collagen production | Anti-fibrotic | [72] | |
| miR-375 | Down | RAC1 | Patient samples, Mouse CCI(4)-treated | Suppresses HSC activation | Anti-fibrotic | [73] |
| miR-378a | Down | TGFB2 | Patient samples, Rat CCI(4)-treated, Human LX-2 cell line | Suppresses HSC proliferation | Anti-fibrotic | [74] |
| miR-378a-3p | Down | Gli3 | Patient samples, Mouse CCI(4)-treated, LX-2, HepG2 and N2a cell lines | Suppresses HSC activation | Anti-fibrotic | [75] |
| Down | Wnt10a | Patient samples, Rat CCI(4)-treated, HSC-T6 cell line | Suppresses HSC proliferation | Anti-fibrotic | [76] | |
| miR-454 | Down | Wnt10a | Rat CCI(4)-treated, HSC-T6 cell line | Suppresses HSC proliferation | Anti-fibrotic | [77] |
| miR-455-3p | Down | HSF1 | Patient samples, Mouse CCI(4)-treated, LX-2 and HSC-T6 cell lines | Suppresses HSC proliferation and ECM proteins | Anti-fibrotic | [78] |
| miR-9 | Down | ABCC1 | Patient samples, Human LX-2, Mouse CCI(4)-treated and primary HSC cells | Suppresses HSC proliferation | Anti-fibrotic | [79] |
| miR-9-5p | Down | TGFBR1/TGFBR2 | Patient samples, Mouse CCI(4)-treated, LX-2 cell line | Suppresses HSC activation | Anti-fibrotic | [80] |
| miR-98 | Down | HLF | Patient samples, Mouse CCI(4)-treated, LX-2 cell line | Suppresses HSC activation | Anti-fibrotic | [81] |
| CircRNA | Expression | Target Molecule | Disease Model | Function | Role in Fibrosis | References |
|---|---|---|---|---|---|---|
| circ_0067835 | Up | miR-155/FOXO3 | Human LX-2 cell line, Mouse CCI(4)-treated | HSC activation via the PI3K/Akt signaling pathway | Profibrotic | [153] |
| circ_0071410 | Up | miR-9-5p | Human LX-2 cell line | HSC activation | Profibrotic | [154] |
| circPWWP2A | Up | miR-203/FSTL1 | Human LX-2 cell line, Mouse CCI(4)-treated | Activates HSC proliferation | Profibrotic | [155] |
| Up | miR-223/TLR4 | Human LX-2 cell line, Mouse CCI(4)-treated | Activates HSC proliferation | Profibrotic | [155] | |
| circRSF1 | Up | miR-146a-5p/RAC1 | Human LX-2 cell line | HSC activation | Profibrotic | [156] |
| circTUBD1 | Up | miR-146a-5p | Human LX-2 cell line | Activates HSC proliferation via TLR4 pathway | Profibrotic | [157] |
| circUBE2K | Up | miR-149-5p/TGFB2 | Human LX-2 cell line, Mouse CCI(4)-treated | Activates HSC proliferation | Profibrotic | [158] |
| circ_0004018 | Down | miR-660-3p/Tep1 | Mouse CCI(4)-treated | Suppresses the HSC proliferation and activation | Anti-fibrotic | [159] |
| circ_0007874/cMTO1 | Down | miR-181b-5p/Pten | Patient samples, Mouse CCI(4)-treated | Suppresses the HSC activation | Anti-fibrotic | [160] |
| Down | miR-17-5p/SMAD7 | Patient samples, LX-2 cell line | Suppresses the HSC activation | Anti-fibrotic | [161] | |
| circ_0070963 | Down | miR-223-3p/LEMD3 | Human LX-2 cell line, Mouse CCI(4)-treated | Suppresses the HSC proliferation | Anti-fibrotic | [162] |
| circFBXW4 | Down | miR-18b-3p/FBXW7 | Patient samples, Human LX-2 cell line, Mouse CCI(4)-treated | Suppresses the HSC proliferation and activation | Anti-fibrotic | [163] |
| circPSD3 | Down | miR-92b-3p/SMAD7 | Patient samples, Human LX-2 cell line, Mouse CCI(4)-treated | Suppresses the HSC proliferation and activation | Anti-fibrotic | [164] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sulaiman, S.A.; Dorairaj, V.; Abdul Ghafar, K.N.; Abdul Murad, N.A. Noncoding RNAs Interactions in Hepatic Stellate Cells during Hepatic Fibrosis. Livers 2021, 1, 263-285. https://doi.org/10.3390/livers1040021
Sulaiman SA, Dorairaj V, Abdul Ghafar KN, Abdul Murad NA. Noncoding RNAs Interactions in Hepatic Stellate Cells during Hepatic Fibrosis. Livers. 2021; 1(4):263-285. https://doi.org/10.3390/livers1040021
Chicago/Turabian StyleSulaiman, Siti Aishah, Vicneswarry Dorairaj, Khairun Nur Abdul Ghafar, and Nor Azian Abdul Murad. 2021. "Noncoding RNAs Interactions in Hepatic Stellate Cells during Hepatic Fibrosis" Livers 1, no. 4: 263-285. https://doi.org/10.3390/livers1040021
APA StyleSulaiman, S. A., Dorairaj, V., Abdul Ghafar, K. N., & Abdul Murad, N. A. (2021). Noncoding RNAs Interactions in Hepatic Stellate Cells during Hepatic Fibrosis. Livers, 1(4), 263-285. https://doi.org/10.3390/livers1040021