
Does Buffelgrass Have a Long Permanence in an Established Pasture? An Analysis of the Population Dynamics of This Exotic Grass in Central Sonora, Mexico

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
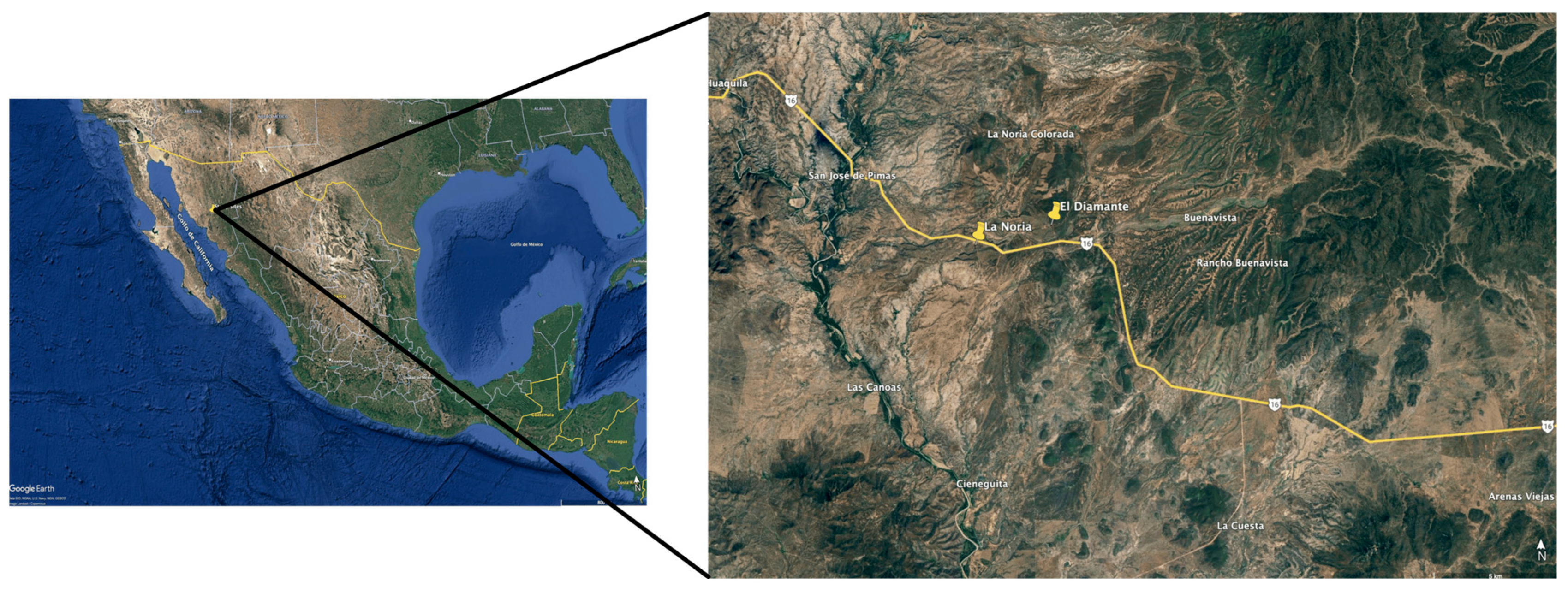
2.1. Study Area
2.2. Field Experiment
2.3. Demographic Model
3. Results
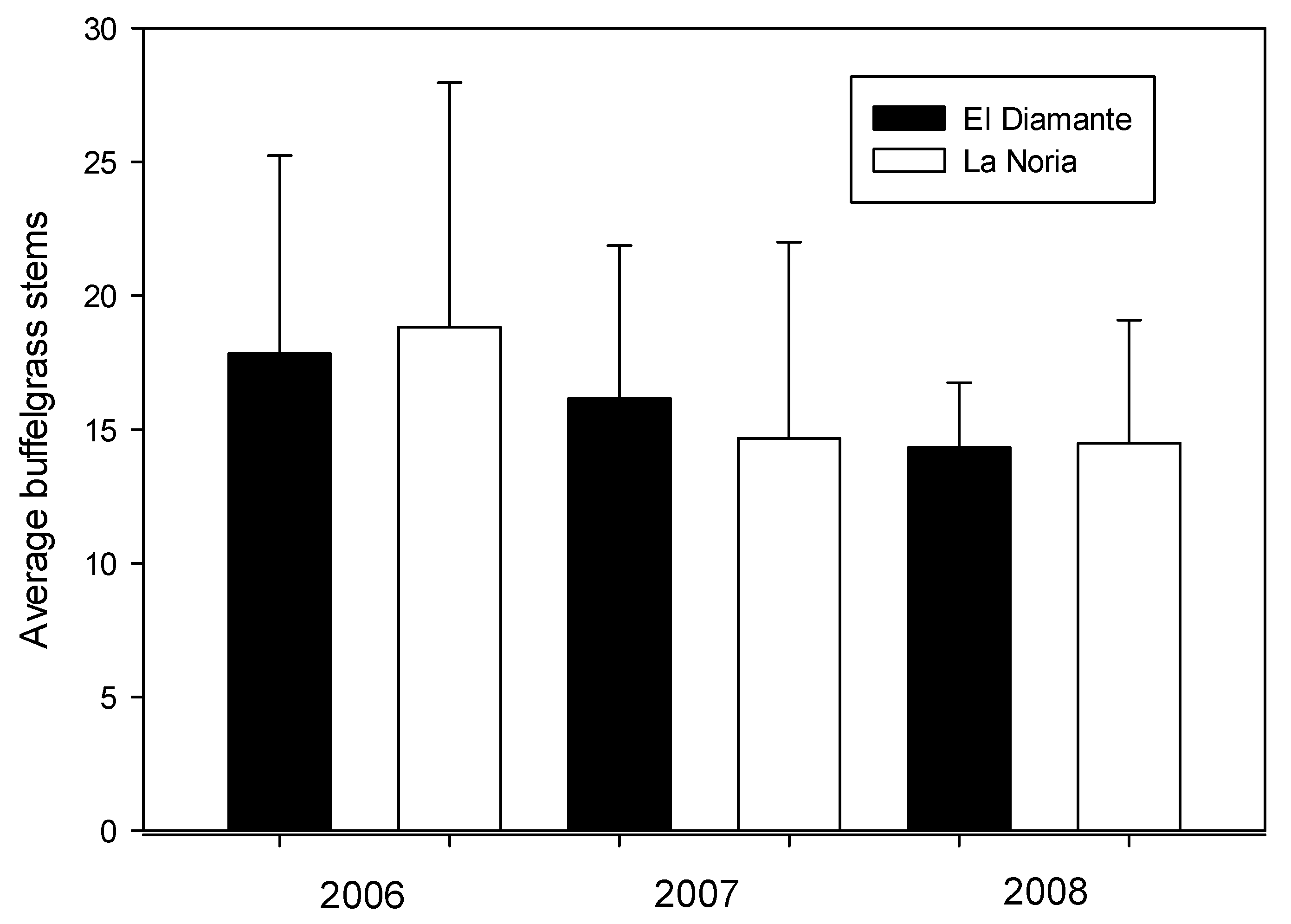
3.1. Comparison of the Total Number of Stems per Plant
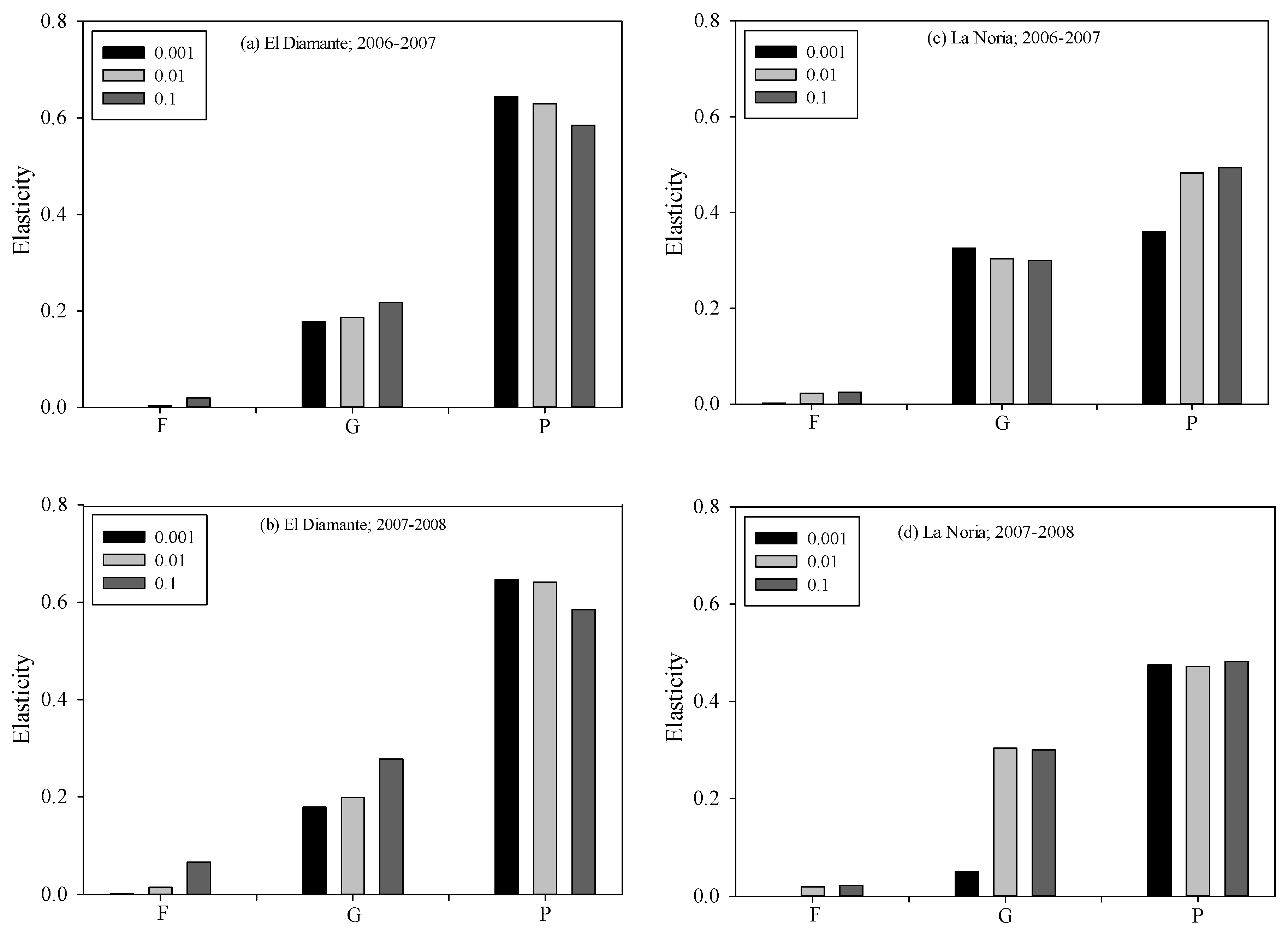
3.2. Population Growth Rates
4. Discussion
Elasticity Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vitousek, P.M.; D’Antonio, C.M.; Loope, L.L.; Rejmánek, M.; Westbrooks, R. Introduced species: A significant component of human-caused global change. N. Z. J. Ecol. 1997, 21, 1–16. [Google Scholar]
- Maass, J.M. Tropical deciduous forest conversion to pasture and agriculture. In Seasonally Dry Tropical Forests; Bullock, S.H., Mooney, H.A., Medina, E., Eds.; Cambridge University Press: Cambridge, UK, 1995; pp. 399–422. [Google Scholar]
- Asner, G.P.; Elmore, A.J.; Olander, L.P.; Martin, R.E.; Harris, A.T. Grazing systems, ecosystem responses, and global change. Annu. Rev. Environ. Resour. 2004, 29, 261–299. [Google Scholar] [CrossRef]
- Ghorbel, M.; Alghamdi, A.; Brini, F.; Hawamda, A.I.; Mseddi, K. Mitigating water loss in arid lands: Buffelgrass as a potential replacement for alfalfa in livestock feed. Agronomy 2025, 15, 371. [Google Scholar] [CrossRef]
- Miller, P.M. Coppice shoot and foliar crown growth after disturbance of a tropical deciduous forest in Mexico. For. Ecol. Manag. 1999, 116, 163–173. [Google Scholar] [CrossRef]
- Quesada, M.; Stoner, K.E.; Rosas-Guerrero, V.; Palacios-Guevara, C.; Lobo, J.A. Effects of habitat disruption on the activity of nectarivorous bats (Chiroptera: Phyllostomidae) in a dry tropical forest: Implications for the reproductive success of the neotropical tree Ceiba grandiflora. Oecologia 2003, 135, 400–406. [Google Scholar] [CrossRef]
- Holl, K.D. Factors limiting tropical rain forest regeneration in abandoned pasture: Seed rain, seed germination, microclimate, and soil 1. Biotropica 1999, 31, 229–242. [Google Scholar] [CrossRef]
- Van Groenendael, J.; Kroon, H.; Caswell, H. Projection matrices in population biology. Trends Ecol. Evol. 1988, 3, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Horvitz, C.C.; Schemske, D.W. Spatio temporal variation in demographic transitions of a tropical understory herb: Projection matrix analysis. Ecol. Monogr. 1995, 65, 155–527. [Google Scholar] [CrossRef]
- Caswell, H. Matrix Populations Models. Construction, Analysis and Interpretation, 2nd ed.; Sinauer Associates: Sunderland, MA, USA, 2001. [Google Scholar]
- Silvertown, J.; Franco, M.; Pisanty, I.; Mendoza, A. Comparative plant demography relative importance of life-cycle components to the finite rate of increase in woody and herbaceous perennials. J. Ecol. 1993, 81, 465–476. [Google Scholar] [CrossRef]
- Schemske, D.W.; Husband, B.C.; Ruckelshaus, M.H.; Goodwillie, C.; Parker, I.M.; Bishop, J.G. Evaluating approaches to the conservation of rare and endangered plants. Ecology 1994, 75, 584–606. [Google Scholar] [CrossRef]
- Marshall, V.M.; Lewis, M.M.M.; Ostendorf, B. Buffelgrass (Cenchrus ciliaris) as an invader and threat to biodiversity in arid environments: A review. J. Arid. Environ. 2012, 78, 1–12. [Google Scholar] [CrossRef]
- Olsson, A.D.; Betancourt, J.L.; Crimmins, M.A.; Marsh, S.E. Constancy of local spread rates for buffelgrass (Pennisetum ciliare L.) in the Arizona upland of the Sonoran Desert. J. Arid. Environ. 2012, 87, 136–143. [Google Scholar] [CrossRef]
- Franklin, K.A.; Lyons, K.; Nagler, P.L.; Lampkin, D.; Glenn, E.P.; Molina-Freaner, F.; Markow, T.; Huete, A.R. Buffelgrass (Pennisetum ciliare) land conversion and productivity in the plains of Sonora, Mexico. Biol. Conserv. 2006, 127, 62–71. [Google Scholar] [CrossRef]
- Hovanes, K.A.; Lien, A.M.; Baldwin, E.; Li, Y.M.; Franklin, K.; Gornish, E.S. Relationship between local-scale topography and vegetation on the invasive C4 perennial bunchgrass buffelgrass (Pennisetum ciliare) size and reproduction. Invasive Plant Sci. Manag. 2023, 16, 38–46. [Google Scholar] [CrossRef]
- Morales-Romero, D.; Rosas-Becerra, R.; Ortega-Rosas, C.I.; Molina-Freaner, F. Can legume seeds establish after land degradation by buffelgrass? Evidence to initiate restoration in Central Sonora, Mexico. Land. Degrad. Dev. 2023, 34, 5430–5437. [Google Scholar] [CrossRef]
- Morales-Romero, D.; Godinez-Alvarez, H.; Campo, J.; Molina-Freaner, F. Effects on land conversion on the regeneration of Pachycereus pecten-aboriginum and its consequences on the population dynamics in Northwestern Mexico. J. Arid. Environ. 2012, 77, 123–129. [Google Scholar] [CrossRef]
- Morales-Romero, D.; Molina-Freaner, F. Influence of buffelgrass pasture conversion on the regeneration and reproduction of the columnar cactus, Pachycereus pecten-aboriginum, in northwestern Mexico. J. Arid. Environ. 2008, 72, 228–237. [Google Scholar] [CrossRef]
- Grigulis, K.; Sheppard, A.W.; Ash, J.E.; Groves, R.H. The comparative demography of the pasture weed Echium plantagineum between its native and invaded ranges. J. Appl. Ecol. 2001, 38, 281–290. [Google Scholar] [CrossRef]
- INEGI. Atlas Nacional del Medio Físico; Instituto Nacional de Estadistica, Geografia e Informatica: Sonora, Mexico, 1988. [Google Scholar]
- Martínez-Yrízar, A.; Felger, R.S.; Búrquez, A. Los Ecosistemas Terrestres: Un Diverso Capital Natural; Molina Freaner, F.E., Van-Devender, T.R., Eds.; Diversidad Biologica de Sonora; UNAM: Mexico City, Mexico, 2010; pp. 129–156. [Google Scholar]
- Vega, E.; Montaña, C. Spatio-temporal variation in the demography of a bunch grass in a patchy semiarid environment. Plant Ecol. 2004, 175, 107–120. [Google Scholar] [CrossRef]
- Commander, L.E.; Golos, P.J.; Miller, B.P.; Merritt, D.J. Seed germination traits of desert perennials. Plant Ecol. 2017, 218, 1077–1091. [Google Scholar] [CrossRef]
- Nantel, P. Démographe, Version 2.0; An Excel add-in for building and running matrix population models; Environment Canada: Ottawa, ON, Canada, 2004.
- Martinez-Lopez, J.R. Influencia de la Precipitación y la Carga Animal Sobre la Productividad Del Buffel (Cenchrus ciliaris L.) Utilizando Analisis de Sistemas y Simulación. Master’s Thesis, Universidad Autonoma de Nuevo Leon, San Nicolás de los Garza, Mexico, 2000. [Google Scholar]
- Franklin, K.; Molina-Freaner, F. Consequences of buffelgrass pasture development for primary productivity, perennial plant richness, and vegetation structure in the drylands of Sonora, Mexico. Conserv. Biol. 2010, 24, 1664–1673. [Google Scholar] [CrossRef] [PubMed]
- Ibarra, F.; Cox, J.R.; Martin-Rivera, M.; Crowl, T.A.; Norton., B.E.; Banner, R.E.; Miller, R.W. Soil physicochemical changes following buffelgrass establishment in Mexico. Arid. Soil. Res. Rehab. 1999, 13, 39–52. [Google Scholar] [CrossRef]
- You, Y.; Liu, Y.; Fujiwara, K. Effects of life-history components on population dynamics of the rare endangered plant Davidia involucrate. Nat. Sci. 2013, 5, 62–70. [Google Scholar]
- Enright, N.J.; Ogden, J. Application of transition matrix models in forest dynamics: Araucaria in New Guinea, and Nothofagus in New Zealand. Aust. J. Ecol. 1979, 4, 3–23. [Google Scholar] [CrossRef]
- Oyama, K. Conservation biology of tropical trees: Demographic and genetic considerations. Environment 1993, 1, 17–32. [Google Scholar]
- Alvarez-Buylla, E.; García-Barrios, R.; Lara-Moreno, C.; Martínez-Ramos, M. Demographic and genetic models in conservation biology: Applications and perspectives for tropical rain forest tree species. Annu. Rev. Ecol. Syst. 1996, 27, 387–421. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Period | Probability | El Diamante (10 yrs After Conversion) | La Noria (50 yrs After Conversion) |
|---|---|---|---|
| 2006–2007 | λ (95% confidence interval) | λ (95% confidence interval) | |
| 0.001 | 1.0018 (1.0016–1.0019) | 1.0015 (1.0014–1.0016) | |
| 0.01 | 1.0166 (1.0164–1.0168) | 1.0376 (1.0373–1.0378) | |
| 0.1 | 1.1050 (1.1048–1.1052) | 1.0414 (1.0412–1.0416) | |
| 2007–2008 | |||
| 0.001 | 1.0002 (1.0001–1.0004) | 1.0013 (1.0011–1.0015) | |
| 0.01 | 1.0033 (1.0030–1.0034) | 1.0317 (1.0315–1.0319) | |
| 0.1 | 1.0233 (1.0232–1.0235) | 1.0355 (1.0352–1.0357) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morales-Romero, D.; Angulo-Cota, R.M.; Ortega-Rosas, C.I.; Cota-Arriola, O.; Molina-Freaner, F. Does Buffelgrass Have a Long Permanence in an Established Pasture? An Analysis of the Population Dynamics of This Exotic Grass in Central Sonora, Mexico. Ecologies 2025, 6, 48. https://doi.org/10.3390/ecologies6030048
Morales-Romero D, Angulo-Cota RM, Ortega-Rosas CI, Cota-Arriola O, Molina-Freaner F. Does Buffelgrass Have a Long Permanence in an Established Pasture? An Analysis of the Population Dynamics of This Exotic Grass in Central Sonora, Mexico. Ecologies. 2025; 6(3):48. https://doi.org/10.3390/ecologies6030048
Chicago/Turabian StyleMorales-Romero, Daniel, Rosa María Angulo-Cota, Carmen Isela Ortega-Rosas, Octavio Cota-Arriola, and Francisco Molina-Freaner. 2025. "Does Buffelgrass Have a Long Permanence in an Established Pasture? An Analysis of the Population Dynamics of This Exotic Grass in Central Sonora, Mexico" Ecologies 6, no. 3: 48. https://doi.org/10.3390/ecologies6030048
APA StyleMorales-Romero, D., Angulo-Cota, R. M., Ortega-Rosas, C. I., Cota-Arriola, O., & Molina-Freaner, F. (2025). Does Buffelgrass Have a Long Permanence in an Established Pasture? An Analysis of the Population Dynamics of This Exotic Grass in Central Sonora, Mexico. Ecologies, 6(3), 48. https://doi.org/10.3390/ecologies6030048