
Heat Wave, Cone Crops, Forest-Floor Small Mammals, and Mustelid Predation in Coniferous Forests of Southern British Columbia

Abstract
1. Introduction
2. Methods
2.1. Study Areas
2.2. Forest-Floor Small Mammals
2.3. Demographic Analysis
2.4. Activity of Weasels
2.5. Statistical Analyses
3. Results
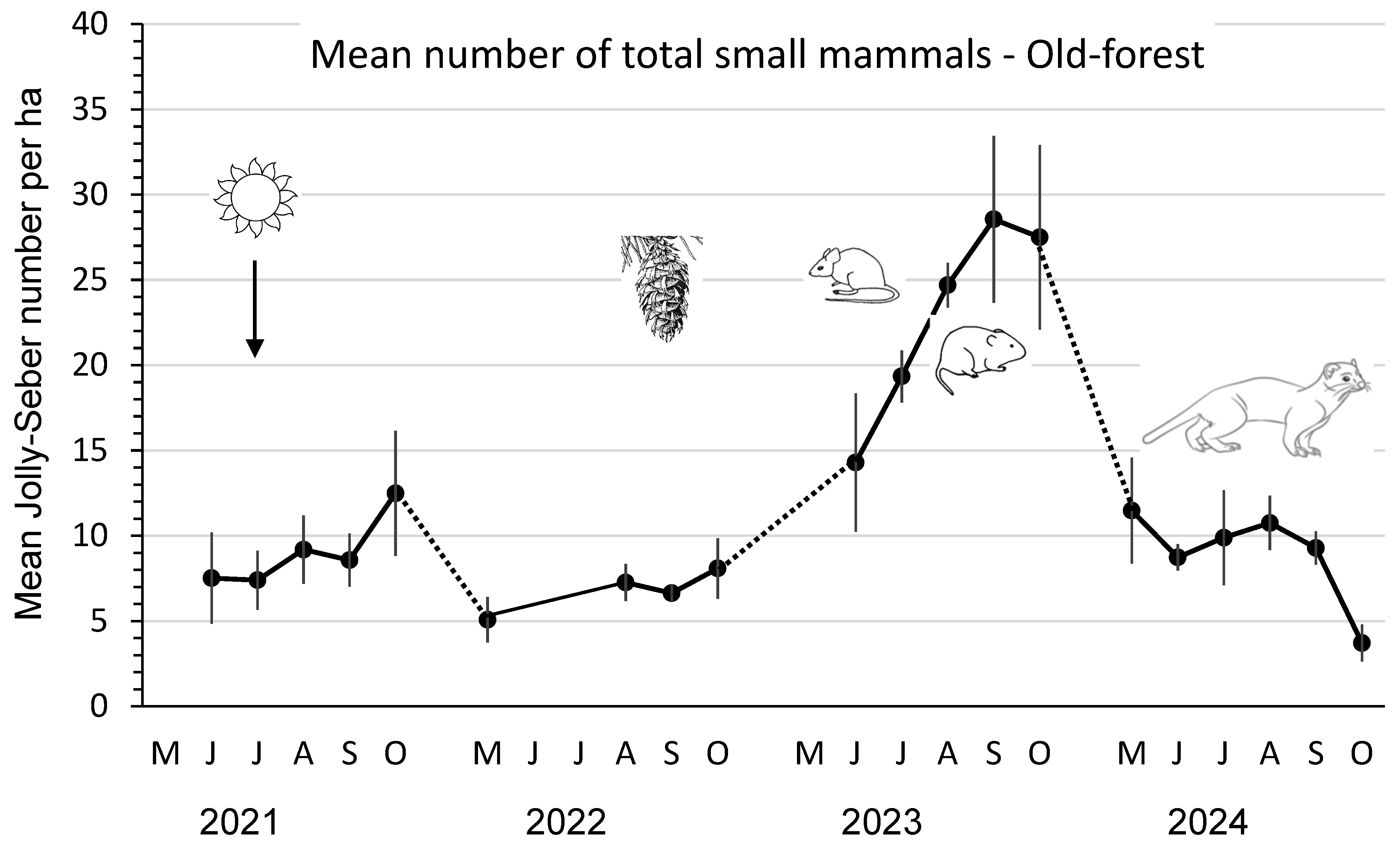
3.1. Forest-Floor Small Mammals
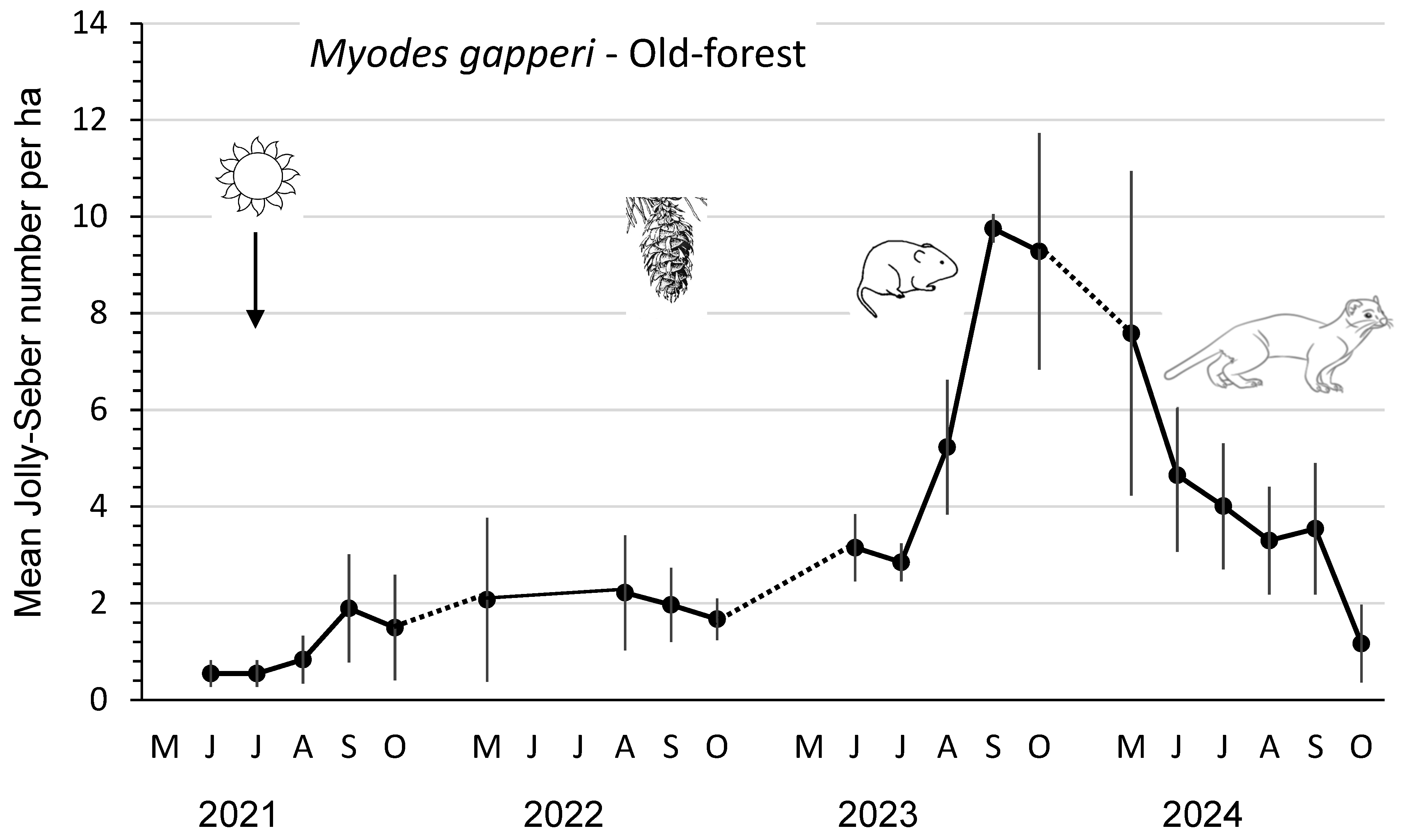
3.2. Small Mammals in Old-Forest Sites
3.3. Small Mammals in Second-Growth Forest Sites
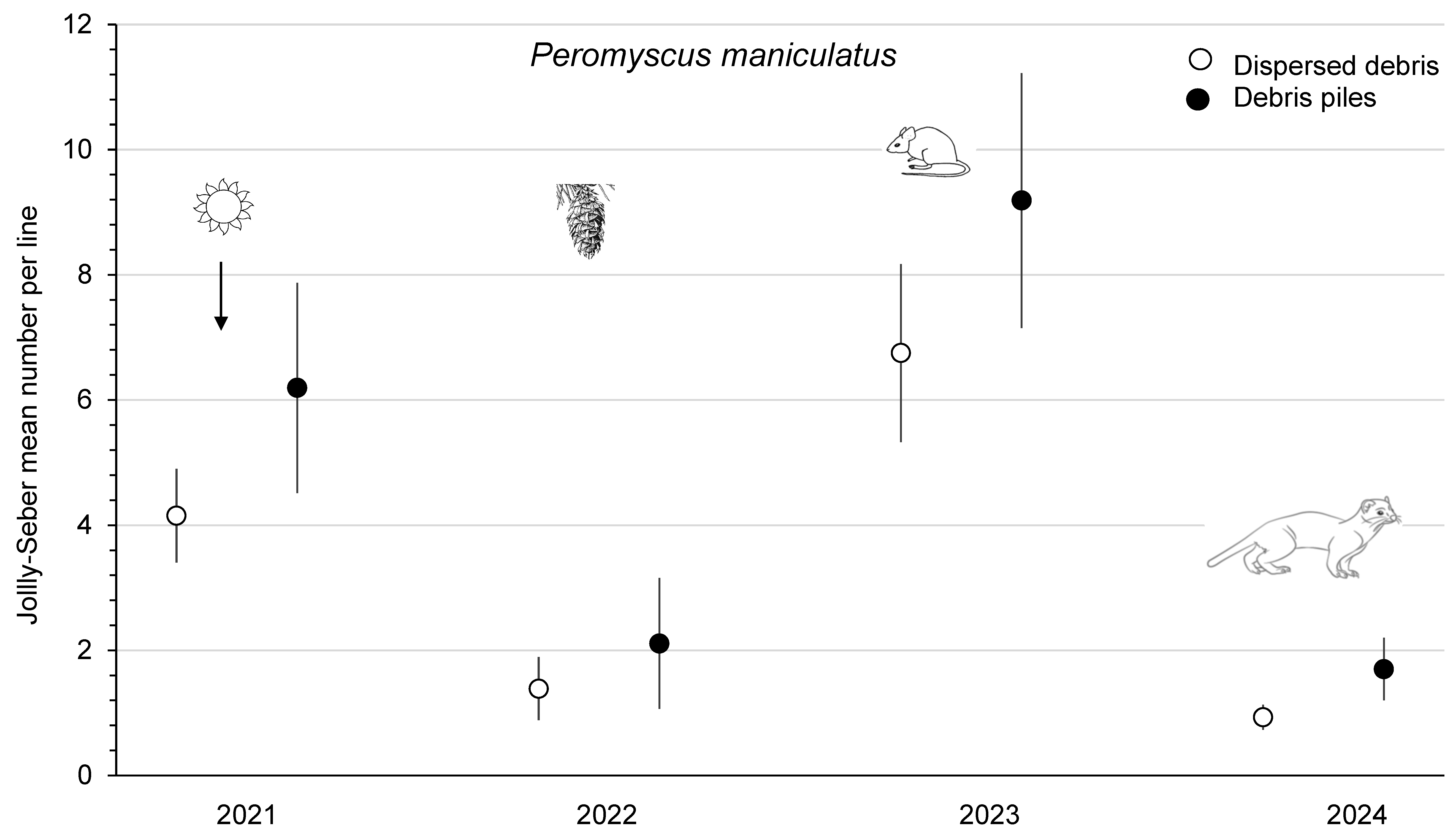
3.4. Small Mammals on Clearcuts
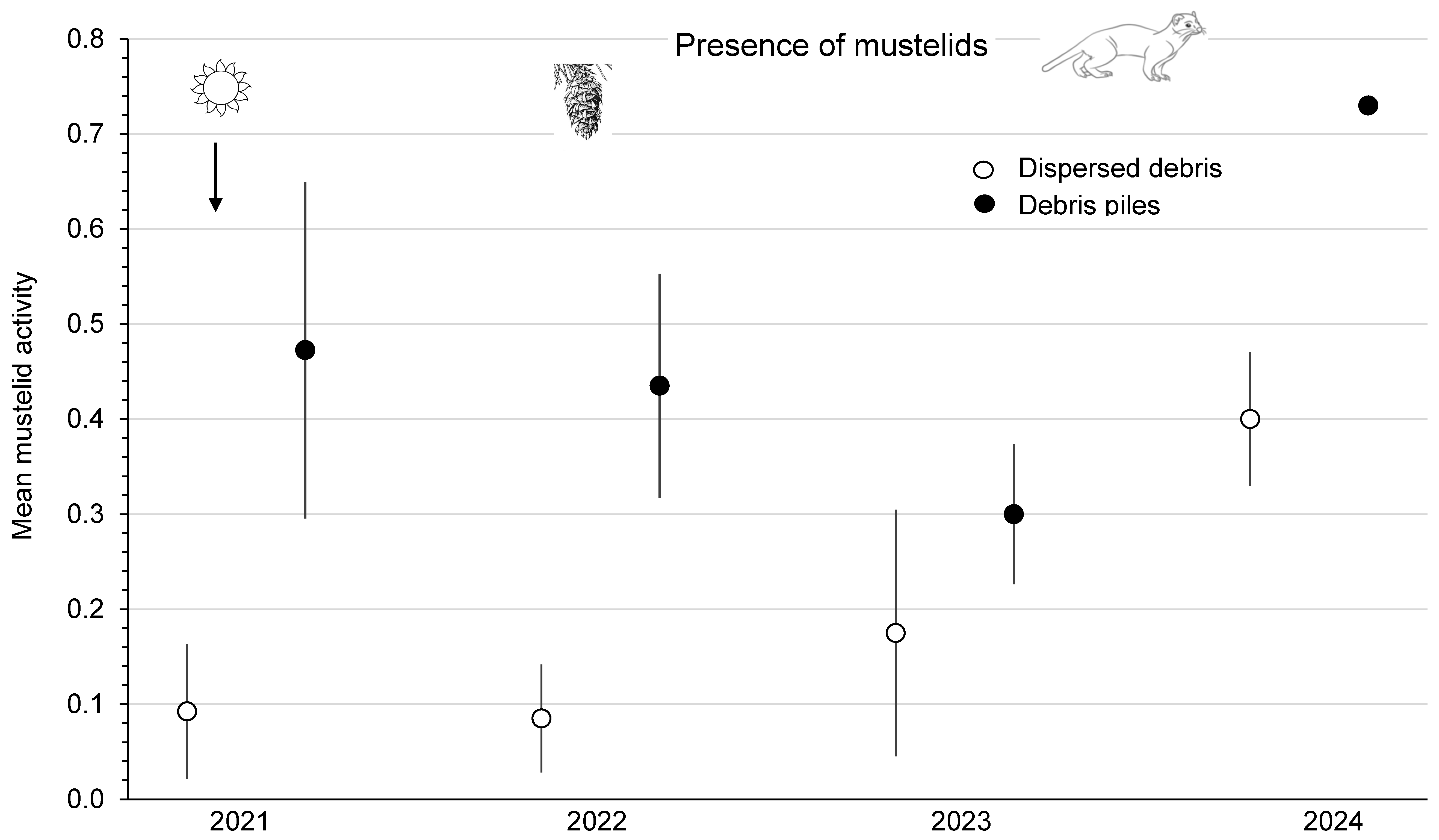
3.5. Activity of Weasels
4. Discussion
4.1. Cone Crops and Small Mammals
4.2. Activity of Mustelids
4.3. Tree Seeds, Small Mammals, and Mustelids
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Koenig, W.D.; Knops, J.M.H. Scale of mast-seeding and tree-ring growth. Nature 1998, 396, 225–226. [Google Scholar] [CrossRef]
- Koenig, W.D.; Knops, J.M.H. The mystery of masting in trees. Am. Sci. 2005, 93, 340–347. [Google Scholar] [CrossRef]
- Thompson, V.; Kennedy-Asser, A.T.; Vosper, E.; Lo, Y.E.; Huntingford, C.; Andrews, O.; Collins, M.; Hegerl, G.C.; Mitchell, D. The 2021 western North America heat wave among the most extreme events ever recorded globally. Sci. Adv. 2022, 8, eabm6860. [Google Scholar] [CrossRef] [PubMed]
- Heeter, K.J.; Harley, G.L.; Abatzoglou, J.T.; Anchukaitis, K.J.; Cook, E.R.; Coulthard, B.L.; Dye, L.A.; Homfeld, I.K. Unprecedented 21st century heat across the Pacific Northwest of North America. Clim. Atmosph. Sci. 2023, 6, 5. [Google Scholar] [CrossRef]
- White, R.H.; Anderson, S.; Booth, J.F.; Braich, G.; Draeger, C.; Fei, C.; Harley, C.D.; Henderson, S.B.; Jakob, M.; Lau, C.A.; et al. The unprecedented Pacific Northwest heat-wave of June 2021. Nat. Commun. 2023, 14, 727. [Google Scholar] [CrossRef]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2015, 6, 1–55. [Google Scholar] [CrossRef]
- Halofsy, J.E.; Peterson, D.L.; Harvey, B.J. Changing wildfire, changing forests: The effects of climate change on fire regimes and vegetation in the Pacific Northwest, USA. Fire Ecol. 2020, 16, 4. [Google Scholar] [CrossRef]
- British Columbia Ministry of Forests. Tree Seed Centre; Kalamalka Lake Tree Seed Orchards: Vernon, BC, Canada, 2023; Available online: https://forestgeneticsbc.ca/wp-content/uploads/2024/02/9-Dave-Kolotelo-Tree-Seed-Centre-Update.pdf (accessed on 20 February 2024).
- Feick, J.L. Conifer Cornucopia. The Great Divide Trail Association. 22 October 2022. Available online: https://www.greatdividetrail.com/conifer-cornucopia/ (accessed on 23 January 2025).
- Hagen, J. A Cone Crop for the Ages. Great Divide’s Fall Newsletter 2022. Great Divide Nature Interpretation. 2022. Available online: https://www.greatdivide.ca/category/newsletters/ (accessed on 23 January 2025).
- Merritt, J.F. Clethrionomys gapperi; Mamm. Species. 1981, No. 146, 1–9. Available online: https://academic.oup.com/mspecies/article/doi/10.2307/3503900/2600558 (accessed on 23 January 2025).
- Sutton, D.A. Tamias amoenus. Mamm. Species 1992, No. 390, 1–8. Available online: https://www.science.smith.edu/departments/biology/VHAYSSEN/msi/pdf/i0076-3519-390-01-0001.pdf (accessed on 23 January 2025).
- Lobo, N.; Duong, M.; Millar, J.S. Conifer-seed preferences of small mammals. Can. J. Zool 2009, 87, 773–780. [Google Scholar] [CrossRef]
- Nagorsen, D.W. Rodents and Lagomorphs of British Columbia; Royal BC Museum: Victoria, BC, Canada, 2005. [Google Scholar]
- Maser, C.; Claridge, A.W.; Trappe, J.M. Trees, Truffles, and Beasts: How Forests Function; Rutgers University Press: Piscataway, NJ, USA, 2008. [Google Scholar]
- Lobo, N.; Millar, J.S. The efficacy of conifer seeds as major food resources to deer mice (Peromyscus maniculatus) and southern red-backed voles (Myodes gapperi). Mammal. Biol. 2011, 76, 274–284. [Google Scholar] [CrossRef]
- Gashwiler, J.S. Tree seed abundance vs. deer mouse populations in Douglas-Fir clearcuts. Proc. Soc. Am. For. 1965, 1965, 219–222. [Google Scholar]
- Gashwiler, J.S. Deer mouse reproduction and its relationship to the tree seed crop. Am. Midl. Nat. 1979, 102, 95–104. [Google Scholar] [CrossRef]
- Sullivan, T.P.; Sullivan, D.S.; Boonstra, R.; Krebs, C.J.; Vyse, A. Population regulation in the deer mouse (Peromyscus maniculatus) in old-growth coniferous forests of southern British Columbia: Insights from a long-term study. Mam. Res. 2023, 68, 37–51. [Google Scholar] [CrossRef]
- Lobo, N.; Millar, J.S. Indirect and mitigated effects of pulsed resources on the population dynamics of a northern rodent. J. Anim. Ecol. 2013, 82, 814–825. [Google Scholar] [CrossRef]
- Elias, S.P.; Witham, J.W.; Hunter, M.L. A cyclic red-backed vole (Clethrionomys gapperi) population and seedfall over 22 years in Maine. J. Mammal. 2006, 87, 440–445. [Google Scholar] [CrossRef]
- Fauteux, D.; Cheveau, M.; Imbeau, L.; Drapeau, P. Cyclic dynamics of a boreal southern red-backed vole population in northwestern Quebec. J. Mammal. 2015, 96, 1–6. [Google Scholar] [CrossRef]
- Sullivan, T.P.; Sullivan, D.S.; Boonstra, R.; Krebs, C.J.; Vyse, A. Mechanisms of population limitation in the southern red-backed vole in conifer forests of western North America: Insights from a long-term study. J. Mammal. 2017, 98, 1367–1378. [Google Scholar] [CrossRef]
- Vander Wall, S.B. Cache site selection by chipmunks (Tamias spp.) and its influence on the effectiveness of seed dispersal in Jeffrey pine (Pinus jeffreyi). Oecologia 1993, 96, 246–252. [Google Scholar] [CrossRef]
- Buskirk, S.W.; Zielinski, W.J. Small and mid-sized carnivores. In Mammal Community Dynamics: Management and Conservation in the Coniferous Forests of Western North America; Zabel, C.J., Anthony, R.G., Eds.; Cambridge University Press: Cambridge, UK, 2003; pp. 207–249. [Google Scholar]
- Sheffield, S.R.; Thomas, H.H. Mustela frenata; Mamm. Species. 1997. No. 570; 1–9. Available online: https://www.science.smith.edu/departments/biology/VHAYSSEN/msi/pdf/i0076-3519-570-01-0001.pdf (accessed on 23 January 2025).
- Simms, D.A. North American weasels: Resource utilization and distribution. Can. J. Zool. 1979, 57, 504–520. [Google Scholar]
- Hargis, C.D.; Bissonette, J.A.; Turner, D.L. The influence of forest fragmentation and landscape pattern on American martens. J. Appl. Ecol. 1999, 36, 157–172. [Google Scholar] [CrossRef]
- King, C.M.; Powell, R.A. The Natural History of Weasels and Stoats: Ecology, Behavior, Management; Oxford University Press: New York, NY, USA, 2007. [Google Scholar]
- Linnell, M.A.; Epps, C.W.; Forsman, E.D.; Zielinski, W.J. Survival and predation of weasels (Mustela erminea, Mustela frenata) in North America. Northwest Sci. 2017, 91, 15–26. [Google Scholar] [CrossRef]
- Evans, B.E.; Mortelliti, A. Forest disturbance and occupancy patterns of American ermine (Mustela richardsonii) and long-tailed weasel (Neogale frenata): Results from a large-scale natural experiment in Maine, United States. J. Mammal. 2022, 103, 1338–1349. [Google Scholar] [CrossRef]
- Lisgo, K.A.; Bunnell, F.L.; Harestad, A.S. Summer and fall use of logging residue piles by female short-tailed weasels. USDA For. Serv. Gen. Tech. Rep. PSW-GTR 2002, 181, 319–329. [Google Scholar]
- Sullivan, T.P.; Sullivan, D.S.; Lindgren, P.M.F.; Ransome, D.B. If we build habitat, will they come? Woody debris structures and conservation of forest mammals. J. Mammal. 2012, 93, 1456–1468. [Google Scholar] [CrossRef]
- Sullivan, T.P.; Sullivan, D.S.; Sullivan, J.H. Mammalian responses to windrows of woody debris on clearcuts: Abundance and diversity of forest-floor small mammals and presence of small mustelids. For. Ecol. Manag. 2017, 399, 143–154. [Google Scholar] [CrossRef]
- Seip, C.; Hodder, D.; Crowley, S.; Johnson, C. Use of constructed coarse woody debris corridors in a clearcut by American martens (Martes americana) and their prey. Forestry 2018, 91, 506–513. [Google Scholar] [CrossRef]
- Ogawa, R.; Mortelliti, A.; Witham, J.W.; Hunter, M.L. Demographic mechanisms linking tree seeds and rodent population fluctuations: Insights from a 33-year study. J. Mammal 2017, 98, 419–427. [Google Scholar] [CrossRef]
- Zwolak, R.; Witczuk, J.; Bogdiewicz, M.; Rychlik, L.; Pagacz, S. Simultaneous population fluctuations of rodents in montane forests and alpine meadows suggest indirect effects of tree masting. J. Mammal. 2018, 99, 586–595. [Google Scholar] [CrossRef]
- Ostfeld, R.S.; Keesing, F. Pulsed resources and community dynamics of consumers in terrestrial ecosystems. Trends Ecol. Evol. 2000, 15, 232–237. [Google Scholar] [CrossRef]
- Pace, M.L.; Cole, J.J.; Carpenter, S.R.; Kitchell, J.F. Trophic cascades revealed in diverse ecosystems. Tree 1999, 14, 483–488. [Google Scholar] [CrossRef]
- Jędrzejewska, B.; Jędrzejewski, W. Predation in Vertebrate Communities: The Bialowieźa Primeval Forest as a Case Study; Springer: Berlin, Germany, 1998. [Google Scholar]
- King, C.M. The relationship between beech (Nothofagus spp.) seedfall and populations of mice (Mus musculus), and the demographic and dietary responses of stoats (Mustela erminea), in three New Zealand forests. J. Anim. Ecol. 1983, 52, 141–166. [Google Scholar] [CrossRef]
- O’Donnell, C.F.J.; Phillipson, S.M. Predicting the incidence of mohua predation from the seedfall, mouse, and predator fluctuations in beech forests. N. Z. J. Zool. 1996, 23, 287–293. [Google Scholar] [CrossRef]
- Johnson, D.R.; Swanson, B.J.; Eger, J.L. Cyclic dynamics of eastern Canadian ermine populations. Can. J. Zool. 2000, 78, 835–839. [Google Scholar] [CrossRef]
- Cheveau, M.; Drapeau, P.; Imbeau, L.; Bergeron, Y. Owl winter irruptions as an indicator of small mammal population cycles in the boreal forest of eastern North America. Oikos 2004, 107, 190–198. [Google Scholar] [CrossRef]
- Schnurr, J.L.; Ostfeld, R.S.; Canham, C.D. Direct and indirect effects of masting on rodent populations and tree seed survival. Oikos 2002, 96, 402–410. [Google Scholar] [CrossRef]
- Zwolak, R.; Celebias, P.; Bogdziewicz, M. Global patterns in the predator satiation effect of masting: A meta-analysis. Proc. Nat. Acad. Sci. USA 2022, 119, e2105655119. [Google Scholar] [CrossRef]
- Bogdziewicz, M.; Zwolak, R.; Crone, E.E. How do vertebrates respond to mast seeding? Oikos 2016, 125, 300–307. [Google Scholar] [CrossRef]
- Meidinger, D.; Pojar, J. Ecosystems of British Columbia; Special Report Series No. 6; Research Branch, BC Ministry of Forests: Victoria, BC, Canada, 1991. [Google Scholar]
- Sullivan, T.P.; Sullivan, D.S. Woody debris structures on large clearcut openings: Oases for small mustelids and prey species? For. Ecol. Manag. 2023, 543, 121117. [Google Scholar] [CrossRef]
- Krebs, C.J.; Keller, B.L.; Tamarin, R.H. Microtus population biology: Demographic changes in fluctuating populations of M. ochrogaster and M. pennsylvanicus in southern Indiana. Ecology 1969, 50, 587–607. [Google Scholar] [CrossRef]
- Nagorsen, D.W. Opossums, Shrews, and Moles of British Columbia. In The Mammals of British Columbia; University of British Columbia Press: Vancouver, BC, Canada, 1996; Volume 2. [Google Scholar]
- Sikes, R.S. The Animal Care and Use Committee of the American Society of Mammalogists. Guidelines of the American Society of Mammalogists for the use of wild mammals in research and education. J. Mammal. 2016, 97, 663–688. [Google Scholar] [CrossRef]
- Pearson, D.E.; Ruggiero, L.F. Transect versus grid trapping arrangements for sampling small-mammal communities. Wildl. Soc. Bull. 2003, 31, 454–459. [Google Scholar]
- Sullivan, T.P.; Sullivan, D.S. Forecasting vole population outbreaks: The rise and fall of a major mammalian pest. For. Ecol. Manag. 2010, 260, 983–993. [Google Scholar] [CrossRef]
- Seber, G.A.F. The Estimation of Animal Abundance and Related Parameters, 2nd ed.; Charles Griffin: London, UK, 1982. [Google Scholar]
- Krebs, C.J. Ecological Methodology; Addison Wesley Longman, Inc.: White Plains, NY, USA, 1999; 624p. [Google Scholar]
- Krebs, C.J.; Boonstra, R.; Gilbert, S.; Reid, D.; Kenney, A.J.; Hofer, E.J. Density estimation for small mammals from livetrapping grids: Rodents in northern Canada. J. Mammal. 2011, 92, 974–981. [Google Scholar] [CrossRef]
- Krebs, C.J.; Boonstra, R. Trappability estimates for mark-recapture data. Can. J. Zool. 1984, 62, 2440–2444. [Google Scholar] [CrossRef]
- Murie, O. A Field Guide to Animal Tracks; The Peterson Field Guide Series 9; Houghton Mifflin Co.: Boston, MA, USA, 1954. [Google Scholar]
- Zielinski, W.J.; Kucera, T.E. (Eds.) American Marten, Fisher, Lynx, and Wolverine: Survey Methods for Their Detection; Gen Tech Rep PSW-GTR-157; USDA Forest Service: Ogden, UT, USA, 1995. [Google Scholar]
- British Columbia Ministry of Environment. Inventory Methods for Marten and Weasels; Standards for Components of British Columbia’s Biodiversity No. 24 Version 2.0; British Columbia Ministry of Environment: Victoria, BC, Canada, 1998. [Google Scholar]
- Rezendes, P. Tracking and the Art of Seeing: How to Read Animal Tracks and Sign; Firefly Books: Willowdale, ON, Canada, 1999. [Google Scholar]
- IBM Corp. IBM SPSS Statistics for Windows, version 29.0; IBM Corp.: Armonk, NY, USA, 2023. [Google Scholar]
- Littel, R.C. Statistical analysis of experiments with repeated measures. HortScience 1989, 24, 36–40. [Google Scholar]
- Kuehl, R.C. Statistical Principles of Research Design and Analysis; Duxbury Press: Belmont, CA, USA, 1994. [Google Scholar]
- Huynh, H.; Feldt, L.S. Estimation of the Box correction for degrees of freedom from sample data in the randomized block and split-plot designs. J. Educ. Stat. 1976, 1, 69–82. [Google Scholar] [CrossRef]
- Fowler, J.; Cohen, L.; Jarvis, P. Practical Statistics for Field Biology, 2nd ed.; John Wiley and Sons: Hoboken, NJ, USA, 1998; 259p. [Google Scholar]
- Saville, D.J. Multiple comparison procedures: The practical solution. Am. Stat. 1990, 44, 174–180. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatistical Analysis; Prentice-Hall, Inc.: Englewood Cliffs, NJ, USA, 1999; 663p. [Google Scholar]
- Banfield, A.W.F. The Mammals of Canada; University of Toronto Press: Toronto, ON, Canada, 1974. [Google Scholar]
- McShea, W.J. The influence of acorn crops on annual variation in rodent and bird populations. Ecology 2000, 81, 228–238. [Google Scholar] [CrossRef]
- Elias, S.P.; Witham, J.W.; Hunter, M.L. Peromyscus leucopus abundance and acorn mast: Population fluctuation patterns over 20 years. J. Mammal. 2004, 85, 743–747. [Google Scholar] [CrossRef]
- Boonstra, R.; Krebs, C.J. Population limitation of the northern red-backed vole in the boreal forests of northern Canada. J. Anim. Ecol. 2006, 75, 1269–1284. [Google Scholar] [CrossRef]
- Sullivan, T.P.; Sullivan, D.S. The use of alternative foods to reduce lodgepole pine seed predation by small mammals. J. Appl. Ecol. 1982, 19, 33–45. [Google Scholar] [CrossRef]
- Zwolak, R. A meta-analysis of the effects of wildfire, clearcutting, and partial harvest on the abundance of North American small mammals. For. Ecol. Manag. 2009, 258, 539–545. [Google Scholar] [CrossRef]
- Rosenberg, D.K.; Anthony, R.G. Differences in Townsend’s chipmunk populations between second- and old-growth forests in western Oregon. J. Wildl. Manag. 1993, 57, 365–373. [Google Scholar] [CrossRef]
- Hayes, J.P.; Horvath, E.G.; Hounihan, P. Townsend’s chipmunk populations in Douglas-fir plantations and mature forests in the Oregon coast range. Can. J. Zool. 1995, 73, 67–73. [Google Scholar] [CrossRef]
- Gashwiler, J.S. Seed fall of three conifers in west-central Oregon. For. Sci. 1969, 15, 290–295. [Google Scholar]
- McCaughey, W.W.; Schmidt, W.C.; Shearer, R.C. ; Shearer, R.C. (Compiler) Seed-dispersal characteristic of conifers in the inland mountain west. In Proceedings of the Conifer Tree Seed in the Inland Mountain West Symposium, Missoula, MT, USA, 5–6 August 1985; Forest Service, U.S. Department of Agriculture, Intermountain Research Station, Northern Region: Albany, CA, USA, 1985. [Google Scholar]
- Weetman, G.; Vyse, A. Natural Regeneration. In Regenerating B.C.’s Forests; Lavender, D.P., Parish, R., Johnson, C., Montgomery, G., Vyse, A., Willis, R., Winston, D., Eds.; UBC Press: Vancouver, BC, Canada, 1990; pp. 118–129. [Google Scholar]
- Greene, D.F.; Zasada, J.C.; Sirois, L.; Kneeshaw, D.; Morin, H.; Charron, I.; Simard, M.-J. A review of the regeneration dynamics of North American boreal forest tree species. Can. J. For. Res. 1999, 29, 824–839. [Google Scholar] [CrossRef]
- Huggard, D.J.; Arsenault, A.; Lloyd, D.; Vyse, A.; Klenner, W. The Opax Mountain Silviculture Systems Project: Preliminary results for managing complex, dry interior Douglas-fir forests. Min. For. Res. Program Ext. Note 2005, 72, 16. [Google Scholar]
- Sachser, F.; Pesendorfer, M.; Gratzer, G.; Nopp-Mayr, U. Differential spatial responses of rodents to masting on forest sites with differing disturbance history. Ecol. Evol. 2021, 11, 11890–11902. [Google Scholar] [CrossRef]
- Sullivan, T.P.; Sullivan, D.S. Influence of stand thinning on wildlife in managed second-growth forests: Tree squirrels, small mustelids, and mammalian species richness. Wildl. Res. 2024, 51, WR24063. [Google Scholar] [CrossRef]
- Henttonen, H.; Oksanen, T.; Jortikka, A.; Haukisalmi, V. How much do weasels shape microtine cycles in the northern Fennoscandian taiga? Oikos 1987, 50, 353–365. [Google Scholar] [CrossRef]
- Hanski, I.; Hansson, L.; Henttonen, H. Specialist predators, generalist predators, and the microtine rodent cycle. J. Anim. Ecol. 1991, 60, 353–367. [Google Scholar] [CrossRef]
- Korpimaki, E.; Norrdahl, K.; Rinta-Jaskari, T. Responses of stoats and least weasels to fluctuating food abundances: Is the low phase of the vole cycle due to mustelid predation? Oecologia 1991, 88, 552–561. [Google Scholar] [CrossRef] [PubMed]
- Cano-Martinez, R.; Carricondo-Sanchez, D.; Devineau, O.; Odden, M. Small rodent cycles influence interactions among predators in a boreal forest ecosystem. Mammal Res. 2021, 66, 592–593. [Google Scholar] [CrossRef]
- Powell, R.A. Structure and spacing of Martes populations. In Martens, Sables, and Fishers: Biology and Conservation; Buskirk, S.W., Harestad, A.S., Raphael, M.G., Powell, R.A., Eds.; Comstock Publishing Associates, Cornell University Press: Ithaca, NY, USA, 1994; pp. 101–121. [Google Scholar]
- Edwards, M.A.; Forbes, G.J. Food habits of ermine, Mustela erminea, in a forested landscape. Can. Field Nat. 2003, 117, 245–248. [Google Scholar] [CrossRef]
- Wilson, P.R.; Karl, B.J.; Toft, R.J.; Beggs, J.R.; Taylor, R.H. The role of introduced predators and competitors in the decline of kaka (Nestor meridionalis) populations in New Zealand. Biol. Conserv. 1998, 83, 175–185. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Old Forest | Second-Growth Forest | Dispersed + Piles of Debris | Dispersed + Piles of Debris |
|---|---|---|---|---|
| Years | 2021–2024 | 2023–2024 | 2021–2024 | 2021–2023 |
| Location | SUM | SUM | SUM | GOLD |
| No. of replicate sites | 3 | 4 | 2 | 2 |
| Monitoring of small mammals | Yes | Yes | Yes | Yes |
| Monitoring of weasel presence | Captures | Captures | Overall activity | Overall activity |
| Methodology | Grids | Grids | Index lines | Index lines |
| Species and Parameter | Year | |||||
|---|---|---|---|---|---|---|
| P. maniculatus | ||||||
| Parameter | 2021 | 2022 | 2023 | 2024 | F3,8 | p |
| Abundance/ha | 6.1 ± 0.1 b | 2.1 ± 0.5 bc | 12.4 ± 1.8 a | 3.1 ± 0.5 b | 22.14 | <0.01 |
| Annual peak abundance/ha | 8.6 ± 0.8 b | 3.8 ± 0.8 b | 17.2 ± 2.7 a | 5.3 ± 1.3 b | 14.06 | <0.01 |
| Number of lactating females | 3.0 ± 1.7 | 1.3 ± 0.9 | 3.7 ± 1.7 | 2.7 ± 2.2 | 0.35 | 0.80 |
| Total number recruits | 25.0 ± 3.5 ab | 6.7 ± 2.8 bc | 40.3 ± 8.1 a | 14.7 ± 1.8 b | 9.49 | <0.01 |
| Total number of juvenile recruits | 19.3 ± 3.4 b | 5.3 ± 2.2 c | 34.3 ± 4.8 a | 12.0 ± 1.0 bc | 15.24 | <0.01 |
| Index of juvenile | 2.72 ± 1.64 | 3.44 ± 2.38 | 13.60 ± 5.13 | 4.33 ± 3.38 | 2.25 | 0.16 |
| productivity | ||||||
| Summer J-S survival | 0.58 ± 0.13 | 0.77 ± 0.12 | 0.66 ± 0.10 | 0.53 ± 0.15 | 1.01 | 0.44 |
| (rate per 28 days) | ||||||
| F2,6 | p | |||||
| Winter J-S survival | 0.78 ± 0.02 | 0.83 ± 0.05 | 0.72 ± 0.02 | 2.03 | 0.21 | |
| (rate per 28 days) | ||||||
| M. gapperi | ||||||
| Parameter | 2021 | 2022 | 2023 | 2024 | F3,8 | p |
| Abundance/ha | 1.1 ± 0.3 c | 2.0 ± 0.8 bc | 6.1 ± 0.4 a | 4.1 ± 1.4 ab | 6.98 | 0.01 |
| Peak abundance/ha | 2.8 ± 1.0 b | 3.5 ± 1.0 b | 11.0 ± 1.5 a | 8.2 ± 3.2 ab | 4.58 | 0.04 |
| Number of lactating females | 1.0 ± 1.0 | 1.7 ± 1.2 | 7.3 ± 3.3 | 5.7 ± 3.7 | 1.40 | 0.31 |
| Total number of recruits | 3.0 ± 0.6 c | 5.0 ± 0.6 bc | 20.7 ± 3.2 a | 11.7 ± 2.7 b | 13.94 | <0.01 |
| Total number of juvenile recruits | 0.7 ± 0.3 c | 2.7 ± 0.3 bc | 13.7 ± 2.7 a | 6.3 ± 0.9 b | 15.53 | <0.01 |
| Index of juvenile | 0.00 ± 0.00 | 0.92 ± 0.58 | 4.90 ± 3.55 | 2.54 ± 1.29 | 1.26 | 0.35 |
| productivity | ||||||
| Summer J-S survival | 0.86 ± 0.14 a | 0.71 ± 0.07 ab | 0.41 ± 0.03 b | 0.48 ± 0.04 b | 5.18 | 0.03 |
| (rate per 28 days) | ||||||
| F2,6 | p | |||||
| Winter J-S survival | 0.94 ± 0.06 | 0.89 ± 0.05 | 0.86 ± 0.05 | 1.36 | 0.33 | |
| (rate per 28 days) | ||||||
| N. amoenus | ||||||
| Parameter | 2021 | 2022 | 2023 | 2024 | F3,8 | p |
| Abundance/ha | 1.3 ± 0.5 | 2.2 ± 0.3 | 3.1 ± 0.6 | 1.3 ± 0.2 | 3.33 | 0.08 |
| Peak abundance/ha | 3.4 ± 1.5 | 3.6 ± 0.8 | 5.2 ± 1.4 | 2.2 ± 0.5 | 1.24 | 0.36 |
| Number of lactating females | 0.7 ± 0.7 | 0.0 ± 0.0 | 0.7 ± 0.7 | 0.3 ± 0.3 | 0.41 | 0.75 |
| Total number of recruits | 5.3 ± 0.9 | 3.3 ± 1.2 | 5.3 ± 0.9 | 2.3 ± 0.9 | 2.38 | 0.15 |
| Total number of juvenile recruits | 1.3 ± 0.9 | 0.0 ± 0.0 | 1.3 ± 0.9 | 0.0 ± 0.0 | 1.52 | 0.28 |
| Index of juvenile | 0.50 ± 0.50 | 0.0 ± 0.0 | 0.50 ± 0.50 | 0.0 ± 0.0 | 0.67 | 0.60 |
| productivity | ||||||
| Summer J-S survival | 0.49 ± 0.29 | 0.95 ± 0.03 | 0.77 ± 0.08 | 0.81 ± 0.06 | 0.44 | 0.73 |
| (rate per 28 days) | ||||||
| F2,6 | p | |||||
| Winter J-S survival | 0.98 ± 0.01 | 0.97 ± 0.03 | 0.96 ± 0.01 | 0.60 | 0.58 | |
| (rate per 28 days) | ||||||
| RM-ANOVA | ||||||||
|---|---|---|---|---|---|---|---|---|
| Treatment | Treatment | Time | Treatment × Time | |||||
| Species + Year | Heavily Thinned | Lightly Thinned | F1,6 | p | F1,6 | p | F1,6 | p |
| P. maniculatus | A | A | 3.29 | 0.12 | 43.45 | <0.01 | 1.61 | 0.25 |
| 2023 | 13.1 ± 2.6 | 8.2 ± 1.6 | ||||||
| 2024 | 2.6 ± 0.5 | 1.1 ± 0.3 | ||||||
| M. gapperi | B | A | 7.57 | 0.03 | 1.57 | 0.26 | 2.24 | 0.19 |
| 2023 | 1.7 ± 0.8 | 4.6 ± 0.4 | ||||||
| 2024 | 1.8 ± 0.9 | 3.6 ± 0.4 | ||||||
| Total number of small mammals | A | A | 0.24 | 0.64 | 62.39 | <0.01 | 0.62 | 0.46 |
| 2023 | 16.9 ± 3.3 | 14.6 ± 1.6 | ||||||
| 2024 | 5.6 ± 1.7 | 5.4 ± 0.4 | ||||||
| RM-ANOVA | ||||||||
|---|---|---|---|---|---|---|---|---|
| Treatment | Treatment | Time | Treatment × Time | |||||
| Parameter and Year | Heavily Thinned | Lightly Thinned | F1,6 | p | F1,6 | p | F1,6 | p |
| P. maniculatus | ||||||||
| Number of lactating females | A | A | 0.09 | 0.78 | 3.77 | 0.10 | 0.79 | 0.08 |
| 2023 | 3.5 ± 1.0 | 3.0 ± 1.2 | ||||||
| 2024 | 1.5 ± 0.6 | 1.5 ± 0.5 | ||||||
| Total number of recruits | A | B | 7.11 | 0.04 | 31.66 | <0.01 | 4.10 | 0.09 |
| 2023 | 36.0 ± 5.8 | 17.0 ± 4.7 | ||||||
| 2024 | 10.5 ± 2.1 | 5.0 ± 2.0 | ||||||
| Total number of juvenile recruits | A | A | 2.82 | 0.14 | 52.09 | <0.01 | 1.72 | 0.24 |
| 2023 | 30.3 ± 5.0 | 21.0 ± 2.9 | ||||||
| 2024 | 7.5 ± 1.7 | 5.3 ± 1.1 | ||||||
| Index of juvenile productivity | 0.14 | 0.72 | 6.63 | 0.04 | 0.22 | 0.65 | ||
| 2023 | 10.98 ± 3.71 | 14.13 ± 6.29 | ||||||
| 2024 | 2.88 ± 1.09 | 2.38 ± 0.80 | ||||||
| M. gapperi | ||||||||
| Number of lactating females | A | A | 4.49 | 0.08 | 4.54 | 0.08 | 0.39 | 0.55 |
| 2023 | 0.5 ± 0.3 | 4.5 ± 1.3 | ||||||
| 2024 | 3.3 ± 1.7 | 6.0 ± 1.6 | ||||||
| Total number of recruits | B | A | 6.94 | 0.04 | 9.48 | 0.02 | 19.36 | <0.01 |
| 2023 | 5.8 ± 2.3 | 17.5 ± 1.0 | ||||||
| 2024 | 7.3 ± 2.9 | 9.0 ± 0.8 | ||||||
| Total number of juvenile recruits | A | A | 5.35 | 0.06 | 7.90 | 0.03 | 20.21 | <0.01 |
| 2023 | 2.5 ± 0.6 | 10.0 ± 1.4 | ||||||
| 2024 | 4.0 ± 1.6 | 3.5 ± 1.2 | ||||||
| Index of juvenile productivity | A | A | 0.68 | 0.44 | 1.83 | 0.23 | 2.10 | 0.20 |
| 2023 | 1.25 ± 0.95 | 2.99 ± 0.93 | ||||||
| 2024 | 1.32 ± 0.63 | 1.09 ± 0.67 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sullivan, T.P.; Sullivan, D.S.; Vyse, A. Heat Wave, Cone Crops, Forest-Floor Small Mammals, and Mustelid Predation in Coniferous Forests of Southern British Columbia. Ecologies 2025, 6, 39. https://doi.org/10.3390/ecologies6020039
Sullivan TP, Sullivan DS, Vyse A. Heat Wave, Cone Crops, Forest-Floor Small Mammals, and Mustelid Predation in Coniferous Forests of Southern British Columbia. Ecologies. 2025; 6(2):39. https://doi.org/10.3390/ecologies6020039
Chicago/Turabian StyleSullivan, Thomas P., Druscilla S. Sullivan, and Alan Vyse. 2025. "Heat Wave, Cone Crops, Forest-Floor Small Mammals, and Mustelid Predation in Coniferous Forests of Southern British Columbia" Ecologies 6, no. 2: 39. https://doi.org/10.3390/ecologies6020039
APA StyleSullivan, T. P., Sullivan, D. S., & Vyse, A. (2025). Heat Wave, Cone Crops, Forest-Floor Small Mammals, and Mustelid Predation in Coniferous Forests of Southern British Columbia. Ecologies, 6(2), 39. https://doi.org/10.3390/ecologies6020039