

Temporal Changes in Fishing Yields, Trophic Dynamics, and Fisheries in Three Mediterranean Lagoons: Logarou and Rodia-Tsoukalio (Greece) and Mar Menor (Spain)

 ,
,  ,
,  , , ,
, , , 

Abstract
1. Introduction
2. Materials and Methods
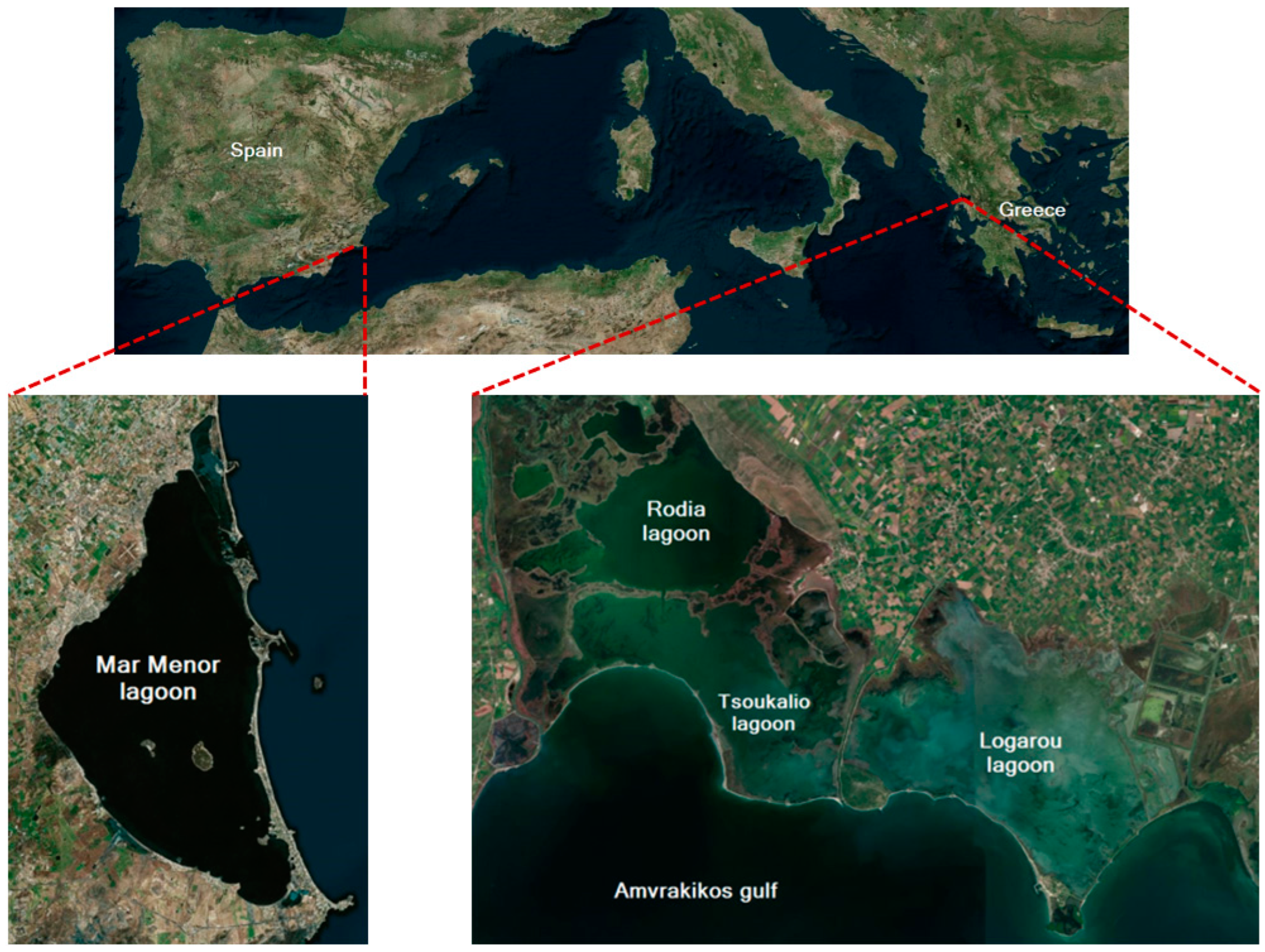
2.1. Target Area
2.2. Datasets
2.3. Data Analysis
3. Results
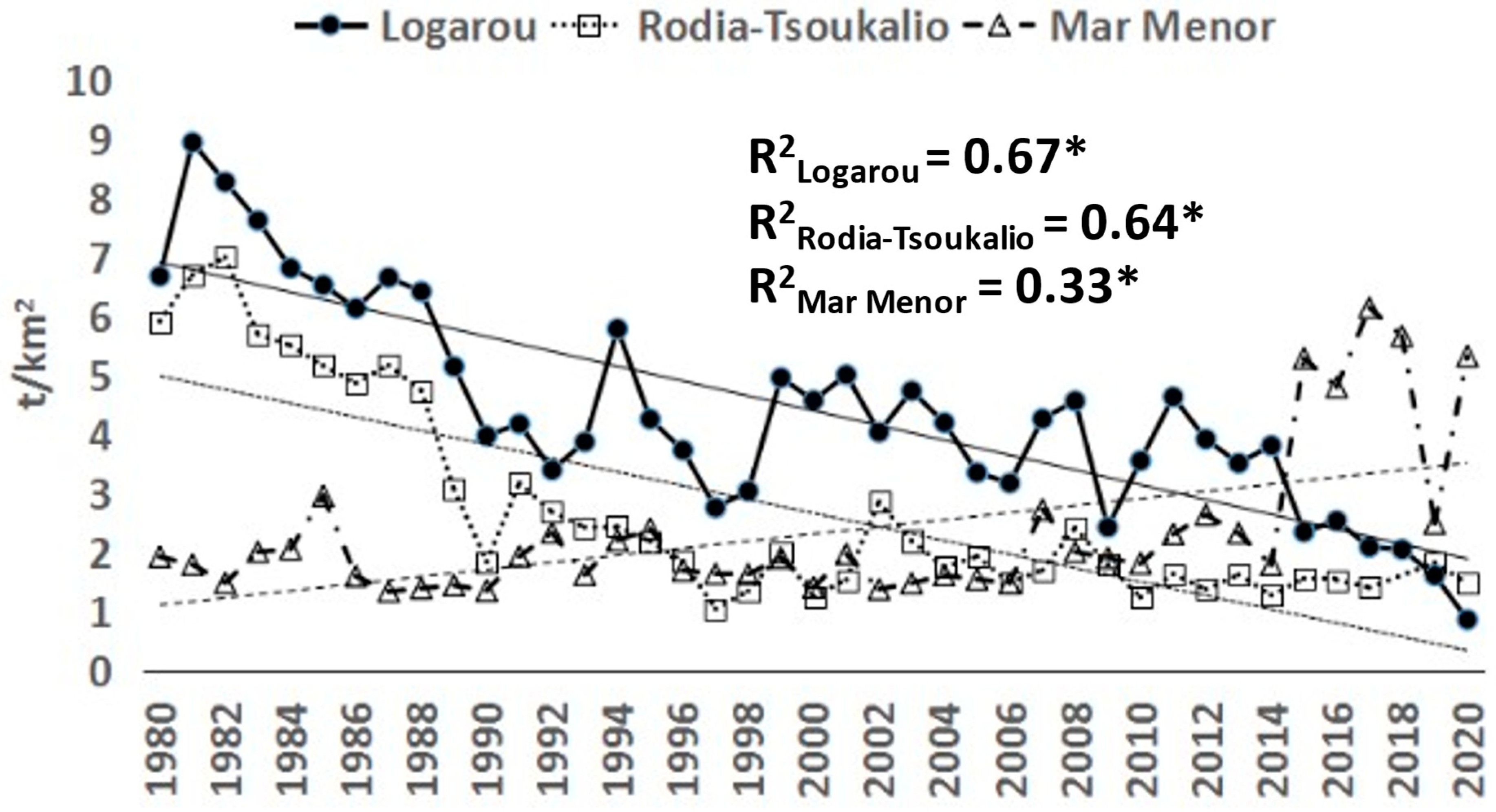
3.1. Time Series Analysis
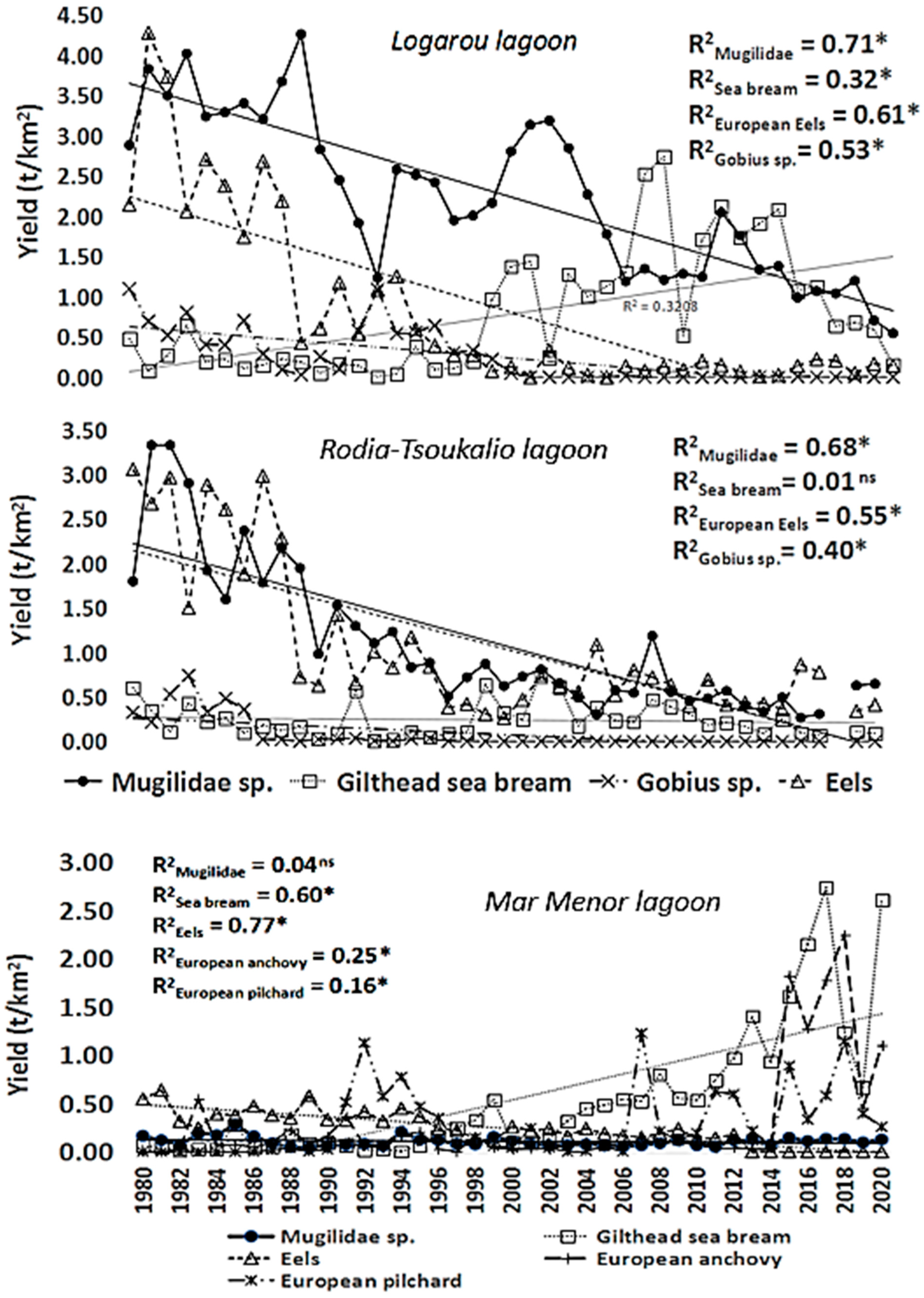
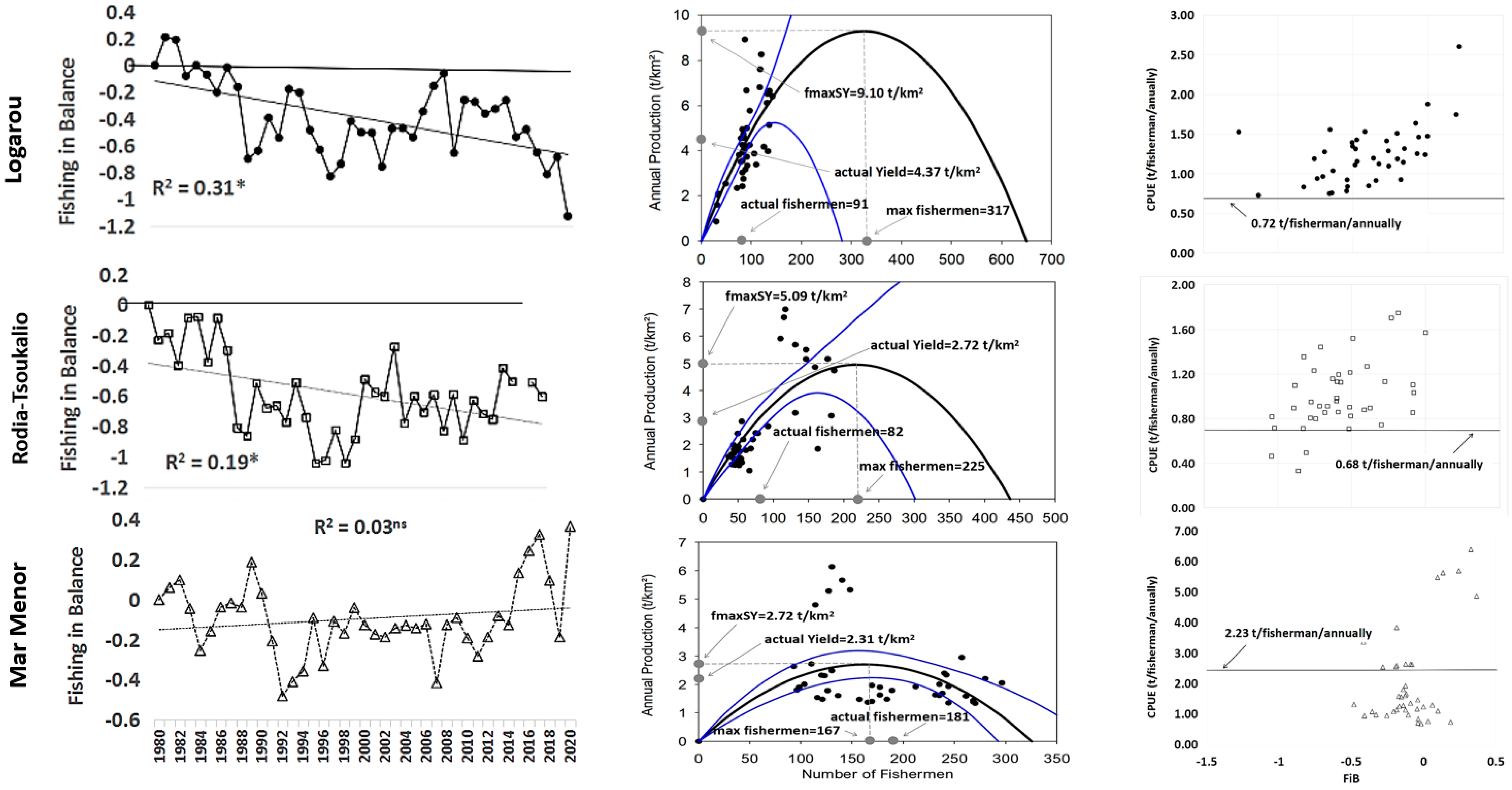
3.2. Logarou Lagoon
3.3. Rodia-Tsoukalio Lagoon
3.4. Mar Menor Lagoon
3.5. Comparison of the Systems Studied
4. Discussion
4.1. Fishing Yield and Studied Lagoon Characteristics
4.2. Species Landing Analysis
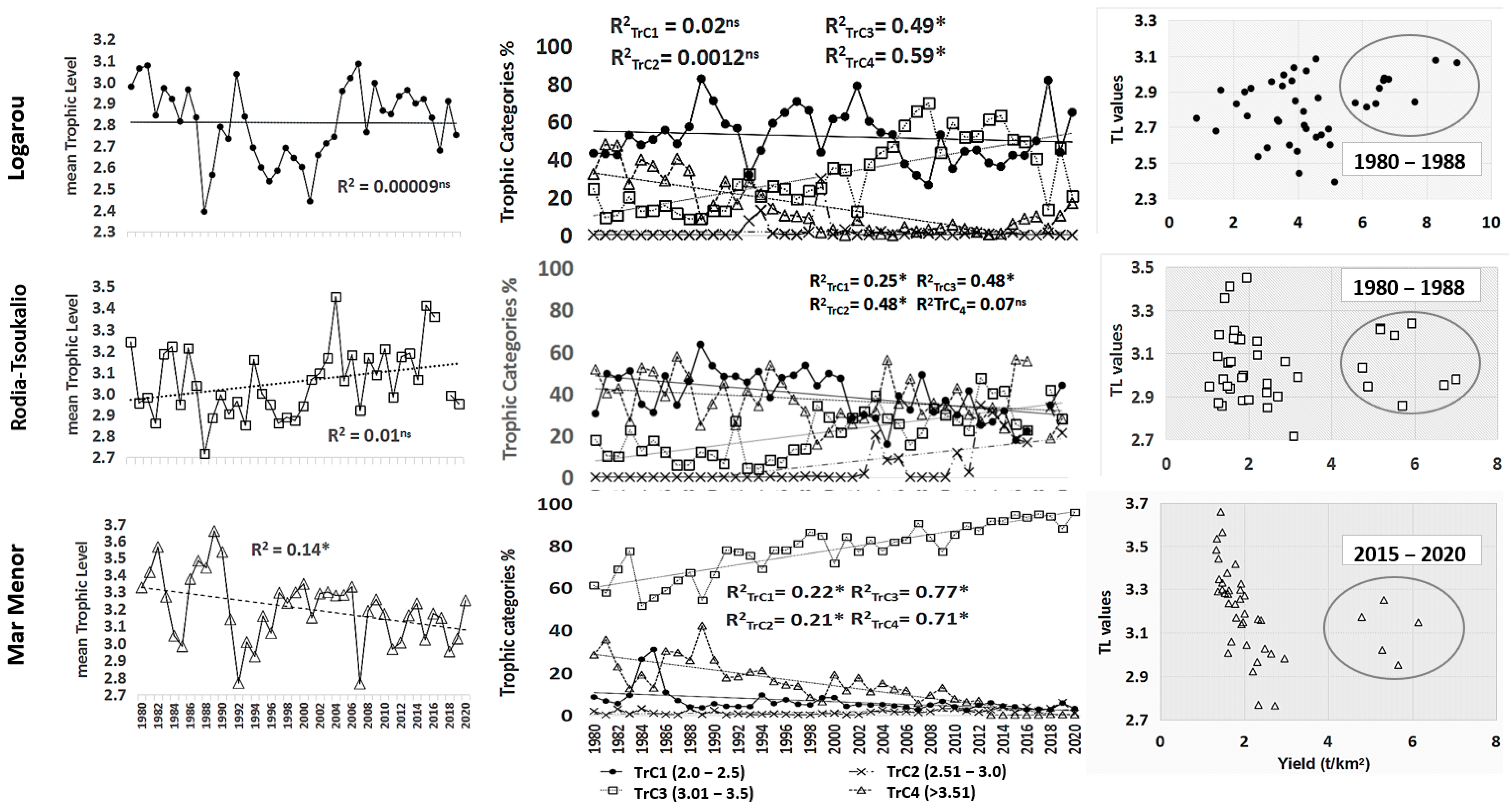
4.3. BTLS, Mean Trophic Level (mTrL), and Trophic Category (TrC)
4.4. Fishing Pressure (FiB and CPUE)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pérez-Ruzafa, A.; Pérez-Ruzafa, I.M.; Newton, A.; Marcos, C. Coastal Lagoons: Environmental Variability, Ecosystem Complexity, and Goods and Services Uniformity. In Coasts and Estuaries; Elsevier: Amsterdam, The Netherlands, 2019; pp. 253–276. [Google Scholar]
- Rodrigues-Filho, J.L.; Macêdo, R.L.; Sarmento, H.; Pimenta, V.R.A.; Alonso, C.; Teixeira, C.R.; Pagliosa, P.R.; Netto, S.A.; Santos, N.C.L.; Daura-Jorge, F.G. From Ecological Functions to Ecosystem Services: Linking Coastal Lagoons Biodiversity with Human Well-Being. Hydrobiologia 2023, 850, 2611–2653. [Google Scholar] [CrossRef]
- Cataudella, S.; Crosetti, D.; Massa, F. Mediterranean Coastal Lagoons: Sustainable Management and Interactions among Aquaculture, Capture Fisheries and the Environment. Gen. Fish. Comm. Mediterr. Stud. Rev. 2015, 95, 278. [Google Scholar]
- Mariani, S. Can Spatial Distribution of Ichthyofauna Describe Marine Influence on Coastal Lagoons? A Central Mediterranean Case Study. Estuar. Coast. Shelf Sci. 2001, 52, 261–267. [Google Scholar] [CrossRef]
- Coleman, J.M.; Huh, O.K.; Braud Jr, D. Wetland Loss in World Deltas. J. Coast. Res. 2008, 24, 1–14. [Google Scholar] [CrossRef]
- de Leiva Moreno, J.I.; Agostini, V.N.; Caddy, J.F.; Carocci, F. Is the Pelagic-Demersal Ratio from Fishery Landings a Useful Proxy for Nutrient Availability? A Preliminary Data Exploration for the Semi-Enclosed Seas around Europe. ICES J. Mar. Sci. 2000, 57, 1091–1102. [Google Scholar] [CrossRef]
- Garrison, L.P.; Link, J.S. Fishing Effects on Spatial Distribution and Trophic Guild Structure of the Fish Community in the Georges Bank Region. ICES J. Mar. Sci. 2000, 57, 723–730. [Google Scholar] [CrossRef]
- Pauly, D.; Christensen, V.; Walters, C. Ecopath, Ecosim, and Ecospace as Tools for Evaluating Ecosystem Impact of Fisheries. ICES J. Mar. Sci. 2000, 57, 697–706. [Google Scholar] [CrossRef]
- Oczkowski, A.; Nixon, S. Increasing Nutrient Concentrations and the Rise and Fall of a Coastal Fishery; a Review of Data from the Nile Delta, Egypt. Estuar. Coast. Shelf Sci. 2008, 77, 309–319. [Google Scholar] [CrossRef]
- Erzini, K.; Sadat, Z.; Bentes, L.; Coelho, R.; Lino, P.G.; Monteiro, P.; Oliveira, F.; Ribeiro, J.; Gonçalves, J.M.S. The Potential Fish Provisioning Services of Vegetated and Unvegetated Habitat in a Lagoon Nursery. Fish. Res. 2024, 278, 107115. [Google Scholar] [CrossRef]
- Pérez-Ruzafa, A.; Molina-Cuberos, G.J.; García-Oliva, M.; Umgiesser, G.; Marcos, C. Why Coastal Lagoons Are so Productive? Physical Bases of Fishing Productivity in Coastal Lagoons. Sci. Total Environ. 2024, 922, 171264. [Google Scholar] [CrossRef]
- Pérez-Ruzafa, A.; Marcos, C. Fisheries in Coastal Lagoons: An Assumed but Poorly Researched Aspect of the Ecology and Functioning of Coastal Lagoons. Estuar. Coast. Shelf Sci. 2012, 110, 15–31. [Google Scholar] [CrossRef]
- Libralato, S.; Pranovi, F.; Raicevich, S.; Da Ponte, F.; Giovanardi, O.; Pastres, R.; Torricelli, P.; Mainardi, D. Ecological Stages of the Venice Lagoon Analysed Using Landing Time Series Data. J. Mar. Syst. 2004, 51, 331–344. [Google Scholar] [CrossRef]
- Pauly, D.; Christensen, V.; Dalsgaard, J.; Froese, R.; Torres Jr, F. Fishing down Marine Food Webs. Science 1998, 279, 860–863. [Google Scholar] [CrossRef] [PubMed]
- Pinnegar, J.K.; Jennings, S.; O’brien, C.M.; Polunin, N.V.C. Long-Term Changes in the Trophic Level of the Celtic Sea Fish Community and Fish Market Price Distribution. J. Appl. Ecol. 2002, 39, 377–390. [Google Scholar] [CrossRef]
- Pauly, D.; Palomares, M.-L. Fishing down Marine Food Web: It Is Far More Pervasive than We Thought. Bull. Mar. Sci. 2005, 76, 197–212. [Google Scholar]
- Sosa-López, A.; Mouillot, D.; Do Chi, T.; Ramos-Miranda, J. Ecological Indicators Based on Fish Biomass Distribution along Trophic Levels: An Application to the Terminos Coastal Lagoon, Mexico. ICES J. Mar. Sci. 2005, 62, 453–458. [Google Scholar] [CrossRef]
- Caddy, J.F.; Garibaldi, L. Apparent Changes in the Trophic Composition of World Marine Harvests: The Perspective from the FAO Capture Database. Ocean Coast. Manag. 2000, 43, 615–655. [Google Scholar] [CrossRef]
- Schaefer, M.B. Some Aspects of the Dynamics of Populations Important to the Management of the Commercial Marine Fisheries. Bull. Math. Biol. 1991, 53, 253–279. [Google Scholar] [CrossRef]
- Joyeux, J.-C.; Ward, A.B. Constraints on Coastal Lagoon Fisheries. In Advances in Marine Biology; Elsevier: Amsterdam, The Netherlands, 1998; Volume 34, pp. 73–199. ISBN 0065-2881. [Google Scholar]
- HCMR. Coastal and Transitional Waters Monitoring Program According to Article 8 of the Directive 2000/60/EC. Annual Report of the 14 Water Districts of Greece for the Year 2014; HCMR: Anavissos, Greece, 2015. [Google Scholar]
- HCMR. “State-of-the-Lagoon Report” for Amvrakikos Lagoon Complex, Western Greece. In ARCH Project (282748) Work Package Report; Conides, A.J., Klaoudatos, D.S., Eds.; Hellenic Center for Marine Research: Anavyssos, Greece, 2012; p. 186. [Google Scholar]
- García-Oliva, M.; Pérez-Ruzafa, Á.; Umgiesser, G.; McKiver, W.; Ghezzo, M.; De Pascalis, F.; Marcos, C. Assessing the Hydrodynamic Response of the Mar Menor Lagoon to Dredging Inlets Interventions through Numerical Modelling. Water 2018, 10, 959. [Google Scholar] [CrossRef]
- Umgiesser, G.; Ferrarin, C.; Cucco, A.; De Pascalis, F.; Bellafiore, D.; Ghezzo, M.; Bajo, M. Comparative Hydrodynamics of 10 Mediterranean Lagoons by Means of Numerical Modeling. J. Geophys. Res. Ocean. 2014, 119, 2212–2226. [Google Scholar] [CrossRef]
- HCMR. Study of Fisheries Management of Lakes (Natural and Artificial), Utilization of Water Resources of Mountainous and Disadvantaged Areas of the Prefectures of Aitoloakarnania. Evritania, Karditsa, Boeotia, Arcadia, Ilia and Achaia. In Final Published Technical Report, in Greek, Hellenic Center for Marine Research—Institute of Inland Waters—Greek Ministry of Agriculture; EU PESCA Programme. Main Document; Hellenic Center for Marine Research: Anavyssos, Greece, 2001; 599p, ISBN 960-86651-2-4. [Google Scholar]
- MAPA. Anuario de Pesca Marítima [Maritime Fisheries Yearbook]; Dirección General de Pesca Marítima. Ministerio de Agricultura, Pesca y Alimentación: Madrid, Spain, 1986. [Google Scholar]
- Katselis, G.N.; Moutopoulos, D.K.; Dimitriou, E.N.; Koutsikopoulos, C. Long-Term Changes of Fisheries Landings in Enclosed Gulf Lagoons (Amvrakikos Gulf, W Greece): Influences of Fishing and Other Human Impacts. Estuar. Coast. Shelf Sci. 2013, 131, 31–40. [Google Scholar] [CrossRef]
- Chou, Y.-L. Statistical Analysis: With Business and Economic Applications; Holt, Rinehart and Winston: New York, NY, USA, 1963; ISBN 0039100618. [Google Scholar]
- Pauly, D.; Christensen, V. Primary Production Required to Sustain Global Fisheries. Nature 1995, 374, 255–257. [Google Scholar] [CrossRef]
- Vivekanandan, E.; Srinath, M.; Kuriakose, S. Fishing the Marine Food Web along the Indian Coast. Fish. Res. 2005, 72, 241–252. [Google Scholar] [CrossRef]
- Stergiou, K.I.; Karpouzi, V.S. Feeding Habits and Trophic Levels of Mediterranean Fish. Rev. Fish Biol. Fish. 2002, 11, 217–254. [Google Scholar] [CrossRef]
- FishBase: World Wide Web Electronic Publication. Available online: https://www.fishbase.org (accessed on 24 April 2025).
- Verdiell-Cubedo, D.; Torralva, M.; Ruiz-Navarro, A.; Oliva-Paterna, F.J. Fish Assemblages in Different Littoral Habitat Types of a Hypersaline Coastal Lagoon (Mar Menor, Mediterranean Sea). Ital. J. Zool. 2013, 80, 104–116. [Google Scholar] [CrossRef]
- Sparre, P.; Venema, S.C. Introduction to Fish Stock Assessment. Part 1: Manual; FAO Fisheries Technical Paper; FAO: Rome, Italy, 1998; Volume 306. [Google Scholar]
- Zar, J.H. Biostatistical Analysis, 4th ed.; Pearson Education: Uttar Pradesh, India, 1999. [Google Scholar]
- Pérez-Ruzafa, A.; Mompeán, M.C.; Marcos, C. Hydrographic, Geomorphologic and Fish Assemblage Relationships in Coastal Lagoons. In Proceedings of the Lagoons and Coastal Wetlands in the Global Change Context: Impacts and Management Issues: Selected papers of the International Conference “CoastWetChange”, Venice, Italy, 26–28 April 2004; Springer: Berlin/Heidelberg, Germany, 2007; pp. 107–125. [Google Scholar]
- Andrisoa, A.; Stieglitz, T.C.; Rodellas, V.; Raimbault, P. Primary Production in Coastal Lagoons Supported by Groundwater Discharge and Porewater Fluxes Inferred from Nitrogen and Carbon Isotope Signatures. Mar. Chem. 2019, 210, 48–60. [Google Scholar] [CrossRef]
- Al-Khalidy, H.I.; Al-Haidarey, M.J.S. Impact of Salinity on Primary Production in the Marshes. Indian J. Ecol. 2019, 46, 614–618. [Google Scholar]
- Reizopoulou, S.; Nicolaidou, A. Benthic Diversity of Coastal Brackish-water Lagoons in Western Greece. Aquat. Conserv. Mar. Freshw. Ecosyst. 2004, 14, S93–S102. [Google Scholar] [CrossRef]
- Ligorini, V.; Garrido, M.; Malet, N.; Simon, L.; Alonso, L.; Bastien, R.; Aiello, A.; Cecchi, P.; Pasqualini, V. Response of Phytoplankton Communities to Variation in Salinity in a Small Mediterranean Coastal Lagoon: Future Management and Foreseen Climate Change Consequences. Water 2023, 15, 3214. [Google Scholar] [CrossRef]
- Ersoy, Z.; Abril, M.; Cañedo-Argüelles, M.; Espinosa, C.; Vendrell-Puigmitja, L.; Proia, L. Experimental Assessment of Salinization Effects on Freshwater Zooplankton Communities and Their Trophic Interactions under Eutrophic Conditions. Environ. Pollut. 2022, 313, 120127. [Google Scholar] [CrossRef]
- Shadrin, N.V.; Anufriieva, E.V.; Kipriyanova, L.M.; Kolesnikova, E.A.; Latushkin, A.A.; Romanov, R.E.; Sergeeva, N.G. The Political Decision Caused the Drastic Ecosystem Shift of the Sivash Bay (the Sea of Azov). Quat. Int. 2018, 475, 4–10. [Google Scholar] [CrossRef]
- Anufriieva, E.V.; El-Shabrawy, G.M.; Shadrin, N.V. Copepoda in the Shallow Hypersaline Bardawil Coastal Lake (Egypt): Are There Long-Term Changes in Composition and Abundance? Oceanol. Hydrobiol. Stud. 2018, 47, 219–229. [Google Scholar] [CrossRef]
- Anufriieva, E.; Kolesnikova, E.; Revkova, T.; Latushkin, A.; Shadrin, N. Human-Induced Sharp Salinity Changes in the World’s Largest Hypersaline Lagoon Bay Sivash (Crimea) and Their Effects on the Ecosystem. Water 2022, 14, 403. [Google Scholar] [CrossRef]
- Meredith, W.; Casamitjana, X.; Quintana, X.D.; Menció, A. Effects of Morphology and Sediment Permeability on Coastal Lagoons’ Hydrological Patterns. J. Hydrol. 2022, 612, 128259. [Google Scholar] [CrossRef]
- Jin, H.; van Leeuwen, C.H.A.; Van de Waal, D.B.; Bakker, E.S. Impacts of Sediment Resuspension on Phytoplankton Biomass Production and Trophic Transfer: Implications for Shallow Lake Restoration. Sci. Total Environ. 2022, 808, 152156. [Google Scholar] [CrossRef]
- Franco, T.P.; Neves, L.M.; Araújo, F.G. Better with More or Less Salt? The Association of Fish Assemblages in Coastal Lagoons with Different Salinity Ranges. Hydrobiologia 2019, 828, 83–100. [Google Scholar] [CrossRef]
- Gray, J.S. Animal-Sediment Relationships. Oceanogr. Mar. Biol. Ann. Rev. 1974, 12, 223–261. [Google Scholar]
- Mouillot, D.; Laune, J.; Tomasini, J.-A.; Aliaume, C.; Brehmer, P.; Dutrieux, E.; Do Chi, T. Assessment of Coastal Lagoon Quality with Taxonomic Diversity Indices of Fish, Zoobenthos and Macrophyte Communities. Hydrobiologia 2005, 550, 121–130. [Google Scholar] [CrossRef]
- Kormas, K.A.; Nicolaidou, A.; Reizopoulou, S. Temporal Variations of Nutrients, Chlorophyll a and Particulate Matter in Three Coastal Lagoons of Amvrakikos Gulf (Ionian Sea, Greece). Mar. Ecol. 2001, 22, 201–213. [Google Scholar] [CrossRef]
- Nicolaidou, A.; Petrou, K.; Kormas, K.A.; Reizopoulou, S. Inter-Annual Variability of Soft Bottom Macrofaunal Communities in Two Ionian Sea Lagoons. Mar. Biodivers. Patterns Process. Assess. Threat. Manag. Conserv. 2006, 183, 89–98. [Google Scholar]
- Guelorget, O.; Perthuisot, J.P. PARALIC ECOSYSTEMS Biological Organization and Functionning. Vie Milieu/Life Environ. 1992, 42, 215–251. [Google Scholar]
- Valiela, I.; McClelland, J.; Hauxwell, J.; Behr, P.J.; Hersh, D.; Foreman, K. Macroalgal Blooms in Shallow Estuaries: Controls and Ecophysiological and Ecosystem Consequences. Limnol. Oceanogr. 1997, 42, 1105–1118. [Google Scholar] [CrossRef]
- Vasileiadou, K.; Pavloudi, C.; Kalantzi, I.; Apostolaki, E.T.; Chatzigeorgiou, G.; Chatzinikolaou, E.; Pafilis, E.; Papageorgiou, N.; Fanini, L.; Konstas, S. Environmental Variability and Heavy Metal Concentrations from Five Lagoons in the Ionian Sea (Amvrakikos Gulf, W Greece). Biodivers. Data J. 2016, 1, e8233. [Google Scholar] [CrossRef] [PubMed]
- Mazzella, L.; Buia, M.C.; Gambi, M.C.; Lorenti, M.; Russo, G.F.; Scipione, M.B.; Zupo, V. Plant-Animal Trophic Relationships in the Posidonia Oceanica Ecosystem of the Mediterranean Sea: A Review. In Plant-Animal Interactions in the Marine Benthos; John, D.M., Howkins, S.J., Price, J.H., Eds.; Claredon Press: Oxford, UK, 1992; pp. 165–187. [Google Scholar]
- Panagopoulos, I.; Mimikou, M. Assessment of the Changes in the Arachtos River Flow and Sediment DISCHARGES Due to Anthropogenic Interventions. In Proceedings of the Protection and Restoration of the Environment VIII, Chania, Greece, 3–7 July 2006. [Google Scholar]
- Moutopolos, D.K.; Ramfos, A.; Spala, K.; Koutsikopoulos, C.; Katselis, G. Long Term Changes of Fisheries Landings Patterns of Most Important Species in Amvrakikos Lagoonal System. In Proceedings of the 9th Symposium on Oceanography and Fisheries, Patra, Greece, 13–16 May 2009; Volume 2, pp. 995–1000. [Google Scholar]
- Ferentinos, G.; Papatheodorou, G.; Geraga, M.; Iatrou, M.; Fakiris, E.; Christodoulou, D.; Dimitriou, E.; Koutsikopoulos, C. Fjord Water Circulation Patterns and Dysoxic/Anoxic Conditions in a Mediterranean Semi-Enclosed Embayment in the Amvrakikos Gulf, Greece. Estuar. Coast. Shelf Sci. 2010, 88, 473–481. [Google Scholar] [CrossRef]
- Pérez-Ruzafa, A.; Marcos-Diego, C.; Ros, J.D. Environmental and Biological Changes Related to Recent Human Activities in the Mar Menor (SE of Spain). Mar. Pollut. Bull. 1991, 23, 747–751. [Google Scholar] [CrossRef]
- Marcos, C.; Torres, I.; López-Capel, A.; Pérez-Ruzafa, A. Long Term Evolution of Fisheries in a Coastal Lagoon Related to Changes in Lagoon Ecology and Human Pressures. Rev. Fish Biol. Fish. 2015, 25, 689–713. [Google Scholar] [CrossRef]
- Giovos, I.; Gonzalvo, J.; Ciprian, M.; Gaentlich, M.; Gavriel, E.; Konstas, S.; Kordopatis, P.; Koutsikopoulos, C.; MaGiovo, I.; Gonzalvo, J.; et al. Amvrakikos Gulf: Biodiversity and Threats. Project “Contributing to the Effective Management of the Amvrakikos Gulf National Park. Available online: https://amvrakikosalliance.net/about/reports/ (accessed on 24 April 2025).
- Dimarchopoulou, D.; Keramidas, I.; Sylaios, G.; Tsikliras, A.C. Ecotrophic Effects of Fishing across the Mediterranean Sea. Water 2021, 13, 482. [Google Scholar] [CrossRef]
- Moutopoulos, D.K.; Libralato, S.; Solidoro, C.; Erzini, K.; Stergiou, K.I. Effect of Landings Data Disaggregation on Ecological Indicators. Mar. Ecol. Prog. Ser. 2014, 509, 27–38. [Google Scholar] [CrossRef]
- Pérez-Ruzafa, A.; Morkune, R.; Marcos, C.; Pérez-Ruzafa, I.M.; Razinkovas-Baziukas, A. Can an Oligotrophic Coastal Lagoon Support High Biological Productivity? Sources and Pathways of Primary Production. Mar. Environ. Res. 2020, 153, 104824. [Google Scholar] [CrossRef]
- Costa, M.J.; Cabral, H.N.; Drake, P.; Economou, A.N.; Fernandez-Delgado, C.; Gordo, L.; Marchand, J.; Thiel, R. Recruitment and Production of Commercial Species in Estuaries. In Fishes in Estuaries; Wiley: Hoboken, NJ, USA, 2002; pp. 54–123. [Google Scholar] [CrossRef]
- Pombo, L.; Elliott, M.; Rebelo, J.E. Changes in the Fish Fauna of the Ria de Aveiro Estuarine Lagoon (Portugal) during the Twentieth Century. J. Fish Biol. 2002, 61, 167–181. [Google Scholar] [CrossRef]
- Malavasi, S.; Fiorin, R.; Franco, A.; Franzoi, P.; Granzotto, A.; Riccato, F.; Mainardi, D. Fish Assemblages of Venice Lagoon Shallow Waters: An Analysis Based on Species, Families and Functional Guilds. J. Mar. Syst. 2004, 51, 19–31. [Google Scholar] [CrossRef]
- Zoulias, T.; Papadopoulos, A.; Conides, A. An Ecological Evaluation Using Fisheries Landings Time-Series Data: A Case Study of Two Coastal Lagoons in the Ionian Sea (E. Mediterranean, Greece). Estuar. Coast. Shelf Sci. 2019, 216, 229–239. [Google Scholar] [CrossRef]
- Malavasi, S.; Franco, A.; Fiorin, R.; Franzoi, P.; Torricelli, P.; Mainardi, D. The Shallow Water Gobiid Assemblage of the Venice Lagoon: Abundance, Seasonal Variation and Habitat Partitioning. J. Fish Biol. 2005, 67, 146–165. [Google Scholar] [CrossRef]
- Ribeiro, J.; Monteiro, C.C.; Monteiro, P.; Bentes, L.; Coelho, R.; Gonçalves, J.M.S.; Lino, P.G.; Erzini, K. Long-Term Changes in Fish Communities of the Ria Formosa Coastal Lagoon (Southern Portugal) Based on Two Studies Made 20 Years Apart. Estuar. Coast. Shelf Sci. 2008, 76, 57–68. [Google Scholar] [CrossRef]
- ICES. Report of the 2006 Session of the Joint European Inland Fisheries Advisory Commision/International Council for the Exploration of the Sea; EIFAC Occasional Paper, 38; Food and Agriculture Organization of the United Nations: Rome, Italy, 2006. [Google Scholar]
- Álvarez-Rogel, J.; Barberá, G.G.; Maxwell, B.; Guerrero-Brotons, M.; Díaz-García, C.; Martínez-Sánchez, J.J.; Sallent, Á.; Martínez-Ródenas, J.; González-Alcaraz, M.N.; Jiménez-Cárceles, F.J. The Case of Mar Menor Eutrophication: State of the Art and Description of Tested Nature-Based Solutions. Ecol. Eng. 2020, 158, 106086. [Google Scholar] [CrossRef]
- Pérez-Ruzafa, A.; Gilabert, J.; Gutiérrez, J.M.; Fernández, A.I.; Marcos, C.; Sabah, S. Evidence of a Planktonic Food Web Response to Changes in Nutrient Input Dynamics in the Mar Menor Coastal Lagoon, Spain. In Proceedings of the Nutrients and Eutrophication in Estuaries and Coastal Waters: Proceedings of the 31st Symposium of the Estuarine and Coastal Sciences Association (ECSA), Bilbao, Spain, 3–7 July 2000; Springer: Berlin/Heidelberg, Germany, 2002; pp. 359–369. [Google Scholar]
- Yunev, O.A.; Carstensen, J.; Moncheva, S.; Khaliulin, A.; Ærtebjerg, G.; Nixon, S. Nutrient and Phytoplankton Trends on the Western Black Sea Shelf in Response to Cultural Eutrophication and Climate Changes. Estuar. Coast. Shelf Sci. 2007, 74, 63–76. [Google Scholar] [CrossRef]
- Zamora-López, A.; Guerrero-Gómez, A.; Torralva, M.; Zamora-Marín, J.M.; Guillén-Beltrán, A.; Oliva-Paterna, F.J. Shallow Waters as Critical Habitats for Fish Assemblages under Eutrophication-Mediated Events in a Coastal Lagoon. Estuar. Coast. Shelf Sci. 2023, 291, 108447. [Google Scholar] [CrossRef]
- Apromar. Aquaculture in Spain; Apromar: Cádiz, Spain, 2020. [Google Scholar]
- FAO. The State of Mediterranean and Black Sea Fisheries; General Fisheries Commission for the Mediterranean: Rome, Italy, 2016; Volume 511. [Google Scholar]
- Dekker, W. Did Lack of Spawners Cause the Collapse of the European Eel, Anguilla Anguilla? Fish. Manag. Ecol. 2003, 10, 365–376. [Google Scholar] [CrossRef]
- Essington, T.E.; Beaudreau, A.H.; Wiedenmann, J. Fishing through Marine Food Webs. Proc. Natl. Acad. Sci. USA 2006, 103, 3171–3175. [Google Scholar] [CrossRef]
- Steneck, R.S. Human Influences on Coastal Ecosystems: Does Overfishing Create Trophic Cascades? Trends Ecol. Evol. 1998, 13, 429–430. [Google Scholar] [CrossRef]
- Pinnegar, J.K.; Polunin, N.V.C.; Badalamenti, F. Long-Term Changes in the Trophic Level of Western Mediterranean Fishery and Aquaculture Landings. Can. J. Fish. Aquat. Sci. 2003, 60, 222–235. [Google Scholar] [CrossRef]
- Pauly, D.; Yáñez-Arancibia, A. Fisheries in Coastal Lagoons. In Elsevier Oceanography Series; Elsevier: Amsterdam, The Netherlands, 1994; Volume 60, pp. 377–399. ISBN 0422-9894. [Google Scholar]
- Kleisner, K.; Pauly, D. The Marine Trophic Index (MTI), the Fishing in Balance (FIB) Index. Fish. Cent. Res. Rep. 2011, 19, 41. [Google Scholar]
- Karageorgis, A.; Krasakopoulou, E.; Pappas, G.; Papageorgiou, A.; Taxiarchi, M. Geochemical Characteristics of Surface Sediments from the Lagoons of Amvrakikos Gulf, Ionian Sea. In Proceedings of the 8th Hellenic Symposium in Oceanography and Fisheries, Thessaloniki, Greece, 4–8 June 2006; pp. 793–799. [Google Scholar]
- Pérez-Ruzafa, A.; Marcos, C.; Bernal, C.M.; Quintino, V.; Freitas, R.; Rodrigues, A.M.; García-Sánchez, M.; Pérez-Ruzafa, I.M. Cymodocea Nodosa vs. Caulerpa Prolifera: Causes and Consequences of a Long Term History of Interaction in Macrophyte Meadows in the Mar Menor Coastal Lagoon (Spain, Southwestern Mediterranean). Estuar. Coast. Shelf Sci. 2012, 110, 101–115. [Google Scholar] [CrossRef]
- Pérez-Ruzafa, A.; Campillo, S.; Fernández-Palacios, J.M.; García-Lacunza, A.; García-Oliva, M.; Ibañez, H.; Navarro-Martínez, P.C.; Pérez-Marcos, M.; Pérez-Ruzafa, I.M.; Quispe-Becerra, J.I. Long-Term Dynamic in Nutrients, Chlorophyll a, and Water Quality Parameters in a Coastal Lagoon during a Process of Eutrophication for Decades, a Sudden Break and a Relatively Rapid Recovery. Front. Mar. Sci. 2019, 6, 26. [Google Scholar] [CrossRef]
- Pérez-Ruzafa, A.; Dezileau, L.; Martínez-Sánchez, M.J.; Pérez-Sirvent, C.; Pérez-Marcos, M.; von Grafenstein, U.; Marcos, C. Long-Term Sediment Records Reveal over Three Thousand Years of Heavy Metal Inputs in the Mar Menor Coastal Lagoon (SE Spain). Sci. Total Environ. 2023, 902, 166417. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trophic Categories | Scientific Names | English Common Name | Lagoon Presence | TL |
|---|---|---|---|---|
| Herbivores–Detrivores (TL: 2.00–2.50) (TrC1) | Sarpa salpa (Linnaeus, 1758) | Salema | ALL | 2.00 |
| Ostrea edulis Linnaeus, 1758 | European flat oyster | MM | 2.06 | |
| Mugilidae | Mullets | ALL | 2.14 | |
| Omnivores (TL: 2.51–3.00) (TrC2) | Crangon crangon (Linnaeus, 1758) | Common shrimp | MM | 2.85 |
| Palaemon adspersus Rathke, 1836 | Prawn | ALL | 2.85 | |
| Gobiidae | Gobies | ALL | 3.00 | |
| Mid-level carnivores (TL: 3.01–3.50) (TrC3) | Diplodus sargus sargus (Linnaeus, 1758) | White sea bream | ALL | 3.10 |
| Engraulis encrasicolus (Linnaeus, 1758) | European anchovy | MM | 3.10 | |
| Sardina pilchardus (Walbaum, 1792) | European pilchard | MM | 3.10 | |
| Bolinus brandaris (Linnaeus, 1758) | Purple dye murex | MM | 3.23 | |
| Solea solea (Linnaeus, 1758) | Common sole | ALL | 3.28 | |
| Atherina boyeri Risso, 1810 | Big-scale sand smelt | MM | 3.30 | |
| Diplodus annularis (Linnaeus, 1758) | Annular sea bream | ALL | 3.30 | |
| Diplodus puntazzo (Walbaum, 1792) | Sharp snout sea bream | ALL | 3.40 | |
| Lithognathus mormyrus (Linnaeus, 1758) | Sand stean bras | ALL | 3.40 | |
| Serranus cabrilla (Linnaeus, 1758) | Comber | MM | 3.40 | |
| Mullidae | Surmullet | ALL | 3.42 | |
| Sparus aurata Linnaeus, 1758 | Gilthead sea bream | ALL | 3.45 | |
| Carcinus aestuarii Nardo, 1847 | Shore green crab | MM | 3.45 | |
| Dicentrarchus labrax (Linnaeus, 1758) | Sea bass | ALL | 3.47 | |
| High-level carnivores (TL:3.51–4.00) (TrC4) | Callinectes sapidus Rathbun, 1896 | Blue crab | MM | 3.73 |
| Serranus scriba (Linnaeus, 1758) | Painted comber | MM | 3.80 | |
| Anguilla anguilla (Linnaeus, 1758) | European eel | ALL | 3.85 |
| Indicator | Logarou | Rodia-Tsoukalio | Mar Menor | Interpretation |
|---|---|---|---|---|
| Yield (t/km2) ± st.dev Min-max values Time series trend | 4.39 (±1.83) 0.86–8.94 Declining | 2.72 (±1.70) 1.05–6.97 Declining | 2.31 (±1.26) 1.34–6.14 Increasing | Yield differences driven by habitat loss in Greek lagoons, eutrophication in the Mar Menor Lagoon. |
| mTrL ± st.dev Min-max values Species dominant Time series trend | 2.81 (±0.18) 2.39–3.08 Herbivorous–detrivorous Stable | 3.05 (±0.16) 2.72–3.45 Herbivorous–detrivorous and high-level carnivorous Stable | 3.20 (±0.20) 2.76–3.66 Mid-level carnivorous Declining | Ecosystem-specific shifts in trophic structure. |
| FiB ± st.dev Min-max values Time series trend | −0.39 (±0.42) −1.13–0.21 Declining | −0.58 (±0.27) −1.04–0.0 Declining | −0.10 (±0.19) −0.48–0.37 Stable | Under-exploitation in Greek lagoons. Balanced exploitation in the Mar Menor Lagoon |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zoulias, T.; Pérez-Ruzafa, A.; Conides, A.; Marcos, C.; Reizopoulou, S.; Vafidis, D.; Klaoudatos, D. Temporal Changes in Fishing Yields, Trophic Dynamics, and Fisheries in Three Mediterranean Lagoons: Logarou and Rodia-Tsoukalio (Greece) and Mar Menor (Spain). Ecologies 2025, 6, 35. https://doi.org/10.3390/ecologies6020035
Zoulias T, Pérez-Ruzafa A, Conides A, Marcos C, Reizopoulou S, Vafidis D, Klaoudatos D. Temporal Changes in Fishing Yields, Trophic Dynamics, and Fisheries in Three Mediterranean Lagoons: Logarou and Rodia-Tsoukalio (Greece) and Mar Menor (Spain). Ecologies. 2025; 6(2):35. https://doi.org/10.3390/ecologies6020035
Chicago/Turabian StyleZoulias, Theodore, Angel Pérez-Ruzafa, Alexis Conides, Concepción Marcos, Sofia Reizopoulou, Dimitris Vafidis, and Dimitris Klaoudatos. 2025. "Temporal Changes in Fishing Yields, Trophic Dynamics, and Fisheries in Three Mediterranean Lagoons: Logarou and Rodia-Tsoukalio (Greece) and Mar Menor (Spain)" Ecologies 6, no. 2: 35. https://doi.org/10.3390/ecologies6020035
APA StyleZoulias, T., Pérez-Ruzafa, A., Conides, A., Marcos, C., Reizopoulou, S., Vafidis, D., & Klaoudatos, D. (2025). Temporal Changes in Fishing Yields, Trophic Dynamics, and Fisheries in Three Mediterranean Lagoons: Logarou and Rodia-Tsoukalio (Greece) and Mar Menor (Spain). Ecologies, 6(2), 35. https://doi.org/10.3390/ecologies6020035