
Rhizosphere Microbiomes Mediating Abiotic Stress Mitigation for Improved Plant Nutrition

 , , ,
, , ,  , ,
, ,
Abstract
1. Introduction
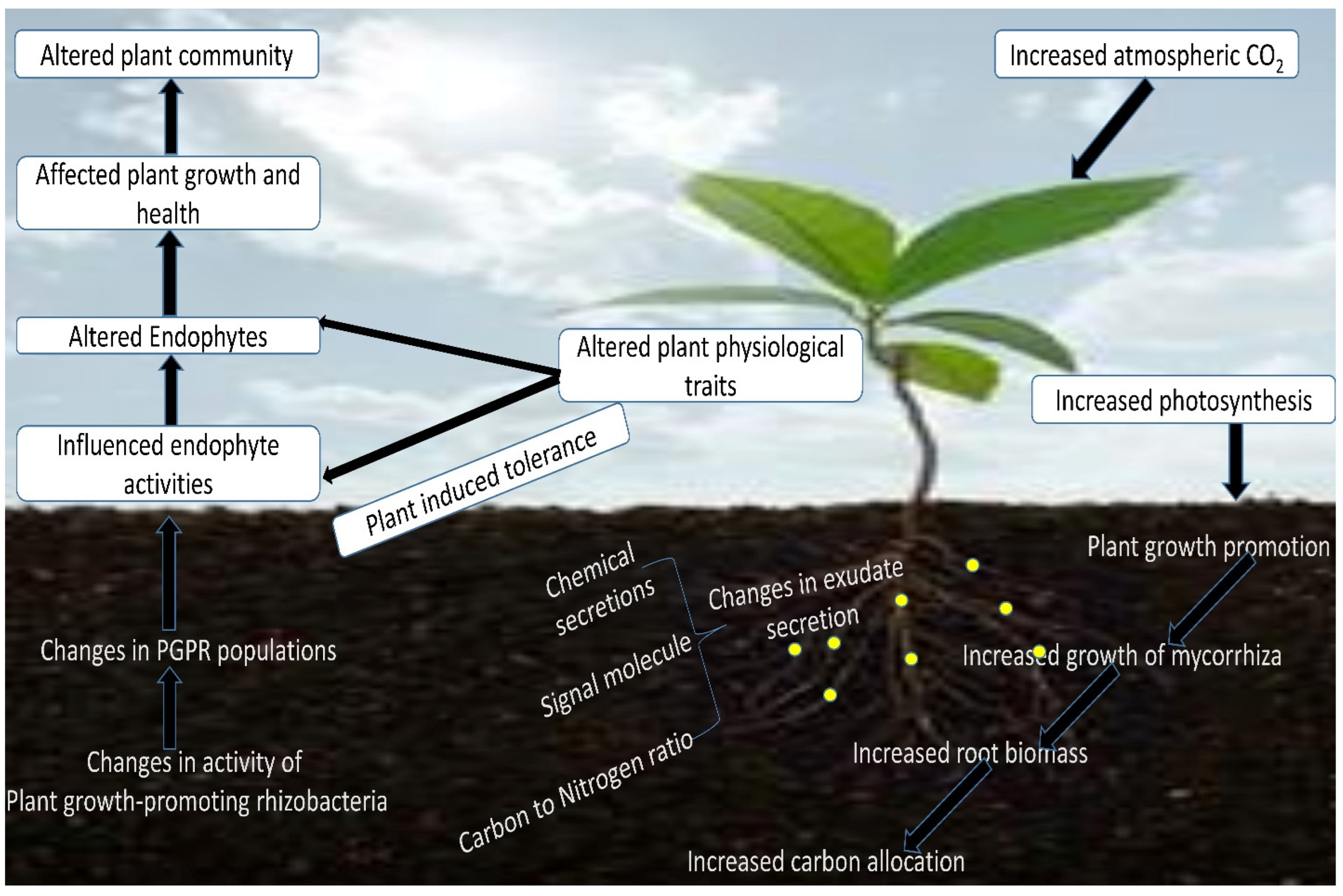
2. Microbiomes in the Rhizosphere Environments
3. Soil Fertility Reduction Alters Soil Microbiome Interactions Due to Climate Change
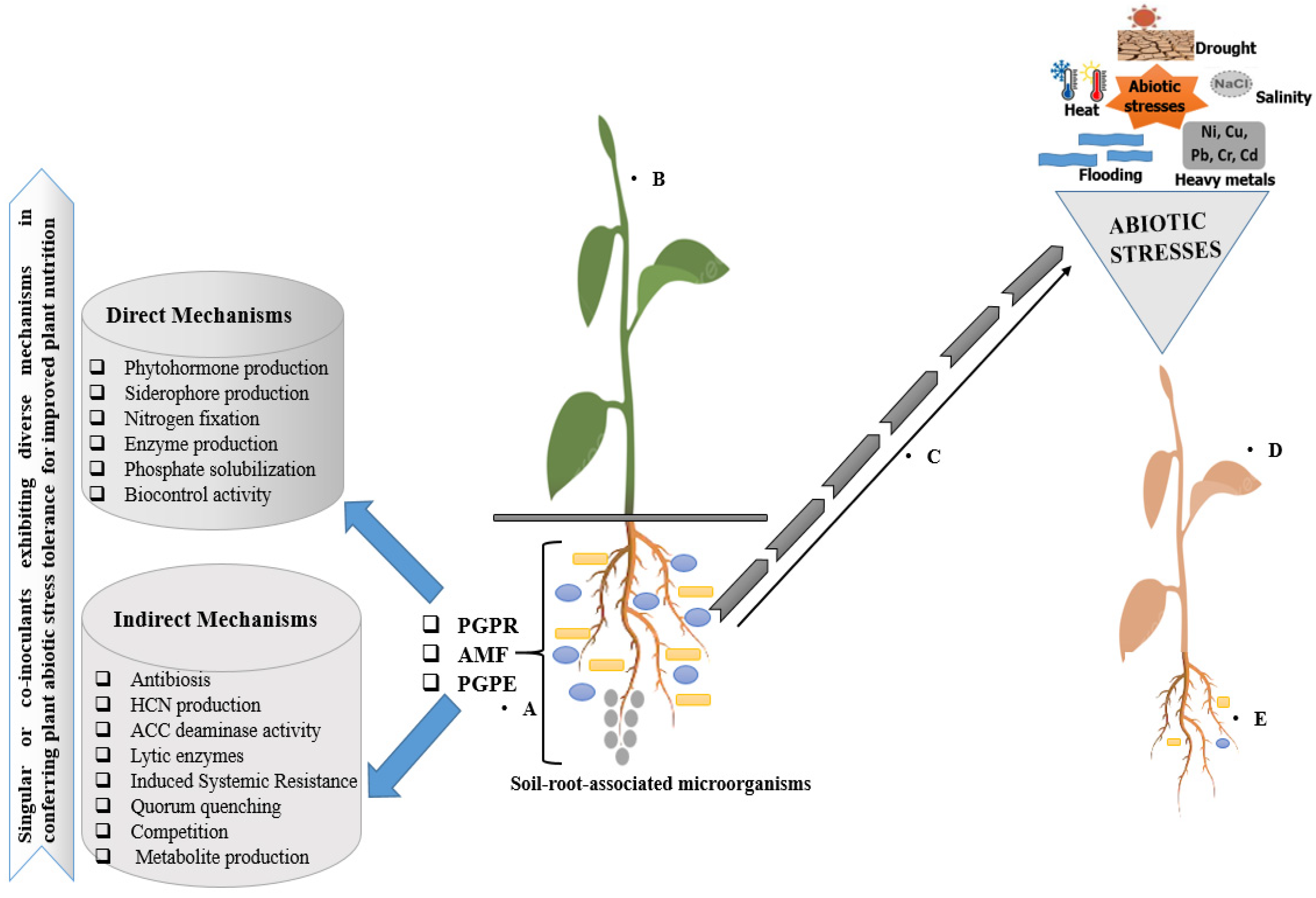
4. Rhizobiome-Mediated Alleviation of Abiotic Stresses
5. Abiotic Stresses Mitigation and Ecological Impacts on Plants

{kind=link}
{kind=link}
{kind=link}
| Abiotic Stress/Metal Toxicity | Microbial Inoculant | Host Plant | Tolerance Type | References |
|---|---|---|---|---|
| Zn, Pb, Cu, AS, and Cd toxicity | Pseudomonas koreensis AGB-1 | Miscanthus sinensis | IAA production and ACC deaminase | [106] |
| Hg toxicity | Photobacterium spp. | Phragmites australis | Mercury reductase and IAA activity. | [107] |
| Zn toxicity | Pseudomonas aeruginosa | Triticum aestivum | Soluble protein, N and P uptake, and improved biomass. | [108] |
| Arsenic toxicity | Staphylococcus arlettae | Brassica juncea | Phosphorus bioavailability in soil through an increased dehydrogenase and phosphatase activities | [109] |
| Heat | Bacillus amyloliquefaciens, Azospirillum brasilence | Triticum aestivum | Decreased generation of ROS, alterations in the metabolome, and preactivation of heat shock transcription factors | [110] |
| Drought | Pseudomonas chlororaphis O6 | Arabidopsis thaliana | Production of volatile compounds, i.e., 2R, 3R butanediol | [111] |
| Drought | Burkholderia phytofirmans Enterobacter sp. FD17 | Zea mays | Increased root and shoot biomass and photosynthesis under drought conditions. | [112] |
| Salinity | Azospirillum brasilense strain Cd | Phaseolus vulgaris | Production of root exudate and flavonoids | [113] |
| Salinity | Bacillus subtilis | Arabidopsis | Decreased root transcriptional expression of a high-affinity potassium ion (K+) transporter (AtHKT1) and decreasing root sodium ion (Na+) import | [114] |
| Salinity | Pantoea dispersa, Azospirillum brasilense | Capsicum annuum | High stomatal conductance and photosynthesis | [115] |
| Salt | Burkholderia, Arthrobacter Bacillus | Vitis vinifera, Capsicum Annuum | Increased accumulation of proline | [116] |
| Osmotic stress | Bacillus megaterium | Zea mays | Increased root expression, high hydraulic conductance, and ZmPIP isoforms | [117] |
| Salt | Cyanobacteria and cyanobacterial extracts | Zea mays, Oryza sativa, Triticum aestivum, Gossypium hirsutum | Production of phytohormones, eliciting molecules | [118] |
| Salt | Bacillus megaterium, Pseudomonas jaduguda, Paenibacillus cookii | Typha angustifolia | Production of growth-promoting and abiotic induction stress genes | [119] |
| Salt | Pseudomonas simiae | Glycine max | Secondary metabolite biosynthesis, i.e., 4-nitroguaiacol and quinolone and stimulation of growth factors | [120] |
| Salt | Bacillus subtilis GB03 | Arabidopsis thaliana | Upregulation of genes involved in sodium transport | [114] |
| Type of Microbes | Abiotic Stress | Plant Host | Mechanism of Action | References |
|---|---|---|---|---|
| Azospirillum lipoferum | Salinity | Cicer arietinum | It helps in the expression of genes related to antioxidants, osmolyte production, and stress | [131] |
| Bacillus sp. | Salinity | Solanum tuberosum | The bacteria increased the production of antioxidant enzymes and auxin. They also increased the uptake of K+, Ca2+, and Na+ | [132] |
| Kocuria rhizophila, Cronobacter sakazakii | Salinity | Triticum aestivum | The microbes enhanced the production of AAC deaminase and antioxidant enzymes | [133] |
| Curtobacterium albidum | Salinity | Oryza sativa | The microbe promotes photosynthetic ability and antioxidative properties | [134] |
| Enterobacter cloacae | Salinity | Brassica napus | The microbe promotes hormonal balancing in plants | [135] |
| Burkholderia sp. | Salinity | Oryza sativa | The organism reduces the production of osmolytes, ROS, and stress, while also increasing the morphological and biochemical parameters | [136] |
| Alcaligenes faecalis | Salinity | Arabidopsis thaliana | The microbes modulate ion transporters and hormonal pathways | [137] |
| Pseudomonas frederiksbergensis, Pseudomonas vancouverensis | Salinity | Capsicum annuum | The organisms modulate stress ethylene levels and increase the antioxidant enzyme activities and plant physiological properties | [138] |
| Bacillus Firmus | Salinity | Glycine max | The organism produces antioxidants and works through gene expression | [139] |
| Curtobacterium sp., Pseudomonas putida, Acinetobacter sp. | Salinity | Sulla carnosa | The microbe reduces oxidative stress and increases the acquisition of plant nutrients from the soil | [140] |
| Klebsiella sp. | Salinity | Avena sativa | The organism helps to enhance the modulation of WRKY1 and rbcL genes | [141] |
| Rhizobacter, Azotobacter | Salinity | Cicer arietinum | Boosting plant salt tolerance for improved yield | [142] |
| Bacillus | Salinity | Oryza sativa | Secretion of regulating hormones and antioxidants for plant resilience to salt stress | [143] |
| Aneurinibacillus, aneurinilyticus, Paenibacillus sp. | Salinity | Phaseolus vulgaris | Production of ACC deaminase gene for plant tolerance to salinity | [144] |
| Streptomyces spp. | Salinity | Steva rebaundia | Microbial stability against salt stress and secretion of plant growth hormones | [145] |
| Ochrobacetrum sp., Microbacterium sp., Enterobacter sp., Enterobacter cloacae | Drought | Poncirus trifoliate | The organisms help to solubilize phosphorus and are involved in ACC deaminase activity, N-fixation, siderophore and IAA production, and P-solubilization | [146] |
| Bacillus thuringiensis | Drought | Zea mays | Enhancing water and nutrient absorption, osmotic balance | [147] |
| Bacillus subtilis | Drought | Triticum aestivum L. | Augmentation of the antioxidant defense system and sugar production. Stimulation of stress-associated gene expression, synthesis of plant hormone | [148] |
| Rhizophagus clarus | Drought | Glycine max | Augmenting water and nutrient assimilation, osmotic balance | [149] |
| Pseudomonas putida | Drought | Cicer arietinum | The microbe helps in the expression of miRNAs | [150] |
| Achromobacter sp., Variovorax paradoxus, Pseudomonas sp., Ochrobactrum anthropic | Drought | Triticum aestivum and Eleusine coracana | The organisms increased the ACC deaminase activity, foliar nutrient concentrations, and enzymatic/nonenzymatic machinery antioxidants | [151] |
| Streptomyces laurentii, Penicillium sp. | Drought | Sorghum bicolor | The bacteria helped to increase the production of siderophores, ammonia, IAA, and hydrogen cyanide. They also help increase the solubilization of zinc and phosphorus | [152] |
| Pseudomonas sp. | Drought | Eleusine coracana | The microbe enhanced the nutrient concentrations and the antioxidant properties of the plants | [153] |
| Pseudomonas azotoformans | Drought | Triticum aestivum | The organism enhances ACC deaminase activity, EPS and IAA production, expression of biofilm genes, and P-solubilization | [154] |
| Pseudomonas sp., Variovorax paradoxus | Drought | Triticum aestivum | These organisms promote the upregulation of helicases and aquaporin and increase ACC deaminase activity and antioxidant properties | [155] |
| Bacillus cereus, Pseudomonas sp. | Drought | Glycine max | The microbe promotes the production of ammonia, the solubilization of phosphorus, and ACC deaminase activity | [156] |
| Arthrobacter arilaitensis, Streptomyces pseudovenezuelae | Drought | Zea mays | The organisms act by solubilizing phosphorus, producing ammonia, IAA, and siderophore through ACC deaminase activity | [157] |
| Rhizobium sp. | Drought | Glycine max | Production of ACC deaminase and drought-resistant genes | [122] |
| Alcaligenes faecalis, Bacillus cereus, Alcaligenes faecalis | Heavy metals (lead, zinc, copper, and cadmium) | Sorghum bicolor | The organisms promote the production of siderophores, proline, EPS, biosurfactants, and salicylic acid | [158] |
| Azotobacter chroococcum | Heavy metals (copper and lead) | Zea mays | The organism facilitates heavy metal chelation | [159] |
| Pseudomonas, Burkholderia | Heavy metals | Lolium perenne | Plant-growth promotion, stress reduction, and metal sequestration, promoting traits under stress | [160] |
| Bacillus | Heavy metal (chromium) | Brassica nigra | Improving plant tolerance to osmotic stress and microbial genes enhancing plant growth | [161] |
| Bacillus | Heavy metals (cadmium and lead) | Solanum nigrum | Production of plant growth hormones, notable genes involved in the removal of pollutants through various pathways | [162] |
| Alcaligenes faecalis, Bacillus Amyloliquefaciens | Heavy metal (lead) | Mentha piperita | Improvement in growth attributes, nutrient concentration, and mitigation of Pb toxicity in mint | [163] |
| Bacillus | Heavy metal (cadmium) | Triticum aestivum L. | Augmentation of the antioxidant defense system and sugar production. Stimulation of stress-associated gene expression and synthesis of plant hormones | [164] |
| Bacillus sp., Enterobacter sp., Pseudomonas fluorescens | Heat | Zea mays | Thermal stress, protein production, antioxidant synthesis | [165] |
| Bacillus safensis | Heat | Solanum lycopersicum | Secretion of thermal regulating genes and antioxidant biosynthesis | [77] |
| Bacillus safensis, Pseudomonas fluorescens | Heavy metals (zinc and lead), temperature, drought | Helianthus annus | Heavy metal binding and elimination of toxins | [166] |
5.1. Tolerance to Drought
5.2. Tolerance to Salinity
5.3. Tolerance to Metal Toxicity and Heat Stress
5.4. Flooding
6. Impacts of Biotechnologically Engineered Soil Microbiomes on Plants
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pérez-Escamilla, R. Food security and the 2015–2030 sustainable development goals: From human to planetary health. Curr. Dev. Nutr. 2017, 1, e000513. [Google Scholar] [CrossRef] [PubMed]
- Akanmu, A.O.; Babalola, O.O.; Venturi, V.; Ayilara, M.S.; Adeleke, B.S.; Amoo, A.E.; Sobowale, A.A.; Fadiji, A.E.; Glick, B.R. Plant disease management: Leveraging on the plant-microbe-soil interface in the biorational use of organic amendments. Front. Plant Sci. 2021, 12, 1590. [Google Scholar] [CrossRef]
- Varzakas, T.; Smaoui, S. Global food security and sustainability issues: The road to 2030 from nutrition and sustainable healthy diets to Food systems change. Foods 2024, 13, 306. [Google Scholar] [CrossRef] [PubMed]
- Lokko, Y.; Heijde, M.; Schebesta, K.; Scholtès, P.; Van Montagu, M.; Giacca, M. Biotechnology and the bioeconomy—Towards inclusive and sustainable industrial development. New Biotechnol. 2018, 40, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Murugesan, P.; Krishanan, A.; Ramasamy, P.; Chandrasekaran, K.; Rajendran, J.; Paramasivam, S.; Chinnaiyan, U.; Panneerselvam, C.; Aziz, A.T.; Alasmari, A. Biorational methods for effective pest control management in stored products for agricultural sustainability. Entomol. Res. 2024, 54, e12697. [Google Scholar] [CrossRef]
- Wakweya, R.B. Challenges and prospects of adopting climate-smart agricultural practices and technologies: Implications for food security. J. Agric. Food Res. 2023, 14, 100698. [Google Scholar] [CrossRef]
- Ayilara, M.S.; Adeleke, B.S.; Babalola, O.O. Bioprospecting and challenges of plant microbiome research for sustainable agriculture, a review on soybean endophytic bacteria. Microb. Ecol. 2022, 85, 1113–1135. [Google Scholar] [CrossRef] [PubMed]
- Gomes, T.; Pereira, J.A.; Benhadi, J.; Lino-Neto, T.; Baptista, P. Endophytic and epiphytic phyllosphere fungal communities are shaped by different environmental factors in a Mediterranean ecosystem. Microb. Ecol. 2018, 76, 668–679. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Glick, B.R. Root Exudate metabolites alter food crops microbiomes, impacting plant biocontrol and growth. Crops 2024, 4, 43–54. [Google Scholar] [CrossRef]
- Gupta, A.; Singh, S.K.; Singh, M.K.; Singh, V.K.; Modi, A.; Singh, P.K.; Kumar, A. Chapter 7—Plant growth–promoting rhizobacteria and their functional role in salinity stress management. In Abatement of Environmental Pollutants; Singh, P., Kumar, A., Borthakur, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 151–160. [Google Scholar]
- Verzeaux, J.; Hirel, B.; Dubois, F.; Lea, P.J.; Tétu, T. Agricultural practices to improve nitrogen use efficiency through the use of arbuscular mycorrhizae: Basic and agronomic aspects. Plant Sci. 2017, 264, 48–56. [Google Scholar] [CrossRef]
- Mrabet, R. Sustainable agriculture for food and nutritional security. In Sustainable Agriculture and the Environment; Elsevier: Amsterdam, The Netherlands, 2023; pp. 25–90. [Google Scholar]
- Turrini, A.; Sbrana, C.; Giovannetti, M. Belowground environmental effects of transgenic crops: A soil microbial perspective. Res. Microbiol. 2015, 166, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, P.; Xu, M.; Ahamad, L.; Chaudhary, A.; Kumar, G.; Adeleke, B.S.; Verma, K.K.; Hu, D.-M.; Širić, I.; Kumar, P. Application of synthetic consortia for improvement of soil fertility, pollution remediation, and agricultural productivity: A review. Agronomy 2023, 13, 643. [Google Scholar] [CrossRef]
- Meena, V.S.; Meena, S.K.; Verma, J.P.; Kumar, A.; Aeron, A.; Mishra, P.K.; Bisht, J.K.; Pattanayak, A.; Naveed, M.; Dotaniya, M.L. Plant beneficial rhizospheric microorganism (PBRM) strategies to improve nutrients use efficiency: A review. Ecol. Eng. 2017, 107, 8–32. [Google Scholar] [CrossRef]
- Gupta, A.; Gopal, M.; Thomas, G.V.; Manikandan, V.; Gajewski, J.; Thomas, G.; Seshagiri, S.; Schuster, S.C.; Rajesh, P.; Gupta, R. Whole genome sequencing and analysis of plant growth promoting bacteria isolated from the rhizosphere of plantation crops coconut, cocoa and arecanut. PLoS ONE 2014, 9, e104259. [Google Scholar] [CrossRef]
- Ercole, T.G.; Kava, V.M.; Aluizio, R.; Pauletti, V.; Hungria, M.; Galli-Terasawa, L.V. Co-inoculation of Bacillus velezensis and Stenotrophomonas maltophilia strains improves growth and salinity tolerance in maize (Zea mays L.). Rhizosphere 2023, 27, 100752. [Google Scholar] [CrossRef]
- Newcombe, G.; Marlin, M.; Barge, E.; Heitmann, S.; Ridout, M.; Busby, P.E. Plant seeds commonly host Bacillus spp., potential antagonists of phytopathogens. Microb. Ecol. 2022, 85, 1356–1366. [Google Scholar] [CrossRef]
- Swarnalakshmi, K.; Yadav, V.; Tyagi, D.; Dhar, D.W.; Kannepalli, A.; Kumar, S. Significance of plant growth promoting rhizobacteria in grain legumes: Growth promotion and crop production. Plants 2020, 9, 1596. [Google Scholar] [CrossRef]
- Khan, N.; Mehmood, A. Revisiting climate change impacts on plant growth and its mitigation with plant growth promoting rhizobacteria. South Afr. J. Bot. 2023, 160, 586–601. [Google Scholar] [CrossRef]
- Lapsansky, E.R.; Milroy, A.M.; Andales, M.J.; Vivanco, J.M. Soil memory as a potential mechanism for encouraging sustainable plant health and productivity. Curr. Opin. Biotechnol. 2016, 38, 137–142. [Google Scholar] [CrossRef]
- Kaur, M.; Karnwal, A. Screening of endophytic bacteria from stress-tolerating plants for abiotic stress tolerance and plant growth-promoting properties: Identification of potential strains for bioremediation and crop enhancement. J. Agric. Food Res. 2023, 14, 100723. [Google Scholar] [CrossRef]
- Filgueiras, L.; Silva, R.; Almeida, I.; Vidal, M.; Baldani, J.I.; Meneses, C.H.S.G. Gluconacetobacter diazotrophicus mitigates drought stress in Oryza sativa L. Plant Soil 2020, 451, 57–73. [Google Scholar] [CrossRef]
- Hossain, M.A.; Hossain, M.S.; Akter, M. Challenges faced by plant growth-promoting bacteria in field-level applications and suggestions to overcome the barriers. Physiol. Mol. Plant Pathol. 2023, 126, 102029. [Google Scholar]
- Shi, S.; Nuccio, E.E.; Shi, Z.J.; He, Z.; Zhou, J.; Firestone, M.K. The interconnected rhizosphere: High network complexity dominates rhizosphere assemblages. Ecol. Lett. 2016, 19, 926–936. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Dippold, M.A.; Kuzyakov, Y.; Razavi, B.S. Spatial pattern of enzyme activities depends on root exudate composition. Soil Biol. Biochem. 2019, 133, 83–93. [Google Scholar] [CrossRef]
- Adeleke, B.S.; Babalola, O.O.; Glick, B.R. Plant growth-promoting root-colonizing bacterial endophytes. Rhizosphere 2021, 20, 100433. [Google Scholar] [CrossRef]
- Debnath, S.; Chakraborty, S.; Langthasa, M.; Choure, K.; Agnihotri, V.; Srivastava, A.; Rai, P.K.; Tilwari, A.; Maheshwari, D.; Pandey, P. Non-rhizobial nodule endophytes improve nodulation, change root exudation pattern and promote the growth of lentil, for prospective application in fallow soil. Front. Plant Sci. 2023, 14, 1152875. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wu, X.; Lin, Z.; Teng, D.; Zhao, Y.; Chen, S.; Hu, X. Screening of cadmium resistant bacteria and their growth promotion of Sorghum bicolor (L.) Moench under cadmium stress. Ecotoxicol. Environ. Saf. 2024, 272, 116012. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, F.X.; Hernández, A.G.; Glick, B.R.; Rossi, M.J. Plant growth-promoting activities and genomic analysis of the stress-resistant Bacillus megaterium STB1, a bacterium of agricultural and biotechnological interest. Biotechnol. Rep. 2020, 25, e00406. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.-M.; Asaf, S.; Khan, A.L.; Khan, A.; Mun, B.-G.; Khan, M.A.; Gul, H.; Lee, I.-J. Complete genome sequence of Pseudomonas psychrotolerans CS51, a plant growth-promoting bacterium, under heavy metal stress conditions. Microorganisms 2020, 8, 382. [Google Scholar] [CrossRef]
- Mwajita, M.R.; Murage, H.; Tani, A.; Kahangi, E.M. Evaluation of rhizosphere, rhizoplane and phyllosphere bacteria and fungi isolated from rice in Kenya for plant growth promoters. SpringerPlus 2013, 2, 606. [Google Scholar] [CrossRef]
- Vives-Peris, V.; de Ollas, C.; Gómez-Cadenas, A.; Pérez-Clemente, R.M. Root exudates: From plant to rhizosphere and beyond. Plant Cell Rep. 2020, 39, 3–17. [Google Scholar] [CrossRef]
- Odelade, K.A.; Babalola, O.O. Bacteria, fungi and archaea domains in rhizospheric soil and their effects in enhancing agricultural productivity. Int. J. Environ. Res. Public Health 2019, 16, 3873. [Google Scholar] [CrossRef] [PubMed]
- Zhalnina, K.; Louie, K.B.; Hao, Z.; Mansoori, N.; da Rocha, U.N.; Shi, S.; Cho, H.; Karaoz, U.; Loqué, D.; Bowen, B.P. Dynamic root exudate chemistry and microbial substrate preferences drive patterns in rhizosphere microbial community assembly. Nat. Microbiol. 2018, 3, 470–480. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.; Pieterse, C.M.; Bakker, P.A.; Berendsen, R.L. Beneficial microbes going underground of root immunity. Plant Cell Environ. 2019, 42, 2860–2870. [Google Scholar] [CrossRef] [PubMed]
- Babalola, O.O.; Adeleke, B.S.; Ayangbenro, A.S. Whole genome sequencing of sunflower root-associated Bacillus cereus. Evol. Bioinform. 2021, 17, 11769343211038948. [Google Scholar] [CrossRef] [PubMed]
- Ngoma, L.; Esau, B.; Babalola, O.O. Isolation and characterization of beneficial indigenous endophytic bacteria for plant growth promoting activity in Molelwane Farm, Mafikeng, South Africa. Afr. J. Biotechnol. 2013, 12, 4104–4114. [Google Scholar]
- Chen, Y.; Ding, Q.; Chao, Y.; Wei, X.; Wang, S.; Qiu, R. Structural development and assembly patterns of the root-associated microbiomes during phytoremediation. Sci. Total Environ. 2018, 644, 1591–1601. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Lang, D.; Wang, J.; Zhang, W.; Zhang, X. Plant-beneficial Streptomyces dioscori SF1 potential biocontrol and plant growth promotion in saline soil within the arid and semi-arid areas. Environ. Sci. Pollut. Res. 2023, 30, 70194–70212. [Google Scholar] [CrossRef] [PubMed]
- Fasusi, O.A.; Amoo, A.E.; Babalola, O.O. Propagation and characterization of viable arbuscular mycorrhizal fungal spores within maize plant (Zea mays L.). J. Sci. Food Agric. 2021, 101, 5834–5841. [Google Scholar] [CrossRef] [PubMed]
- Sritongon, N.; Boonlue, S.; Mongkolthanaruk, W.; Jogloy, S.; Riddech, N. The combination of multiple plant growth promotion and hydrolytic enzyme producing rhizobacteria and their effect on Jerusalem artichoke growth improvement. Sci. Rep. 2023, 13, 5917. [Google Scholar] [CrossRef]
- Hu, L.; Robert, C.A.; Cadot, S.; Zhang, X.; Ye, M.; Li, B.; Manzo, D.; Chervet, N.; Steinger, T.; Van Der Heijden, M.G. Root exudate metabolites drive plant-soil feedbacks on growth and defense by shaping the rhizosphere microbiota. Nat. Commun. 2018, 9, 2738. [Google Scholar] [CrossRef]
- Tsunoda, T.; van Dam, N.M. Root chemical traits and their roles in belowground biotic interactions. Pedobiologia 2017, 65, 58–67. [Google Scholar] [CrossRef]
- Postma, J.A.; Schurr, U.; Fiorani, F. Dynamic root growth and architecture responses to limiting nutrient availability: Linking physiological models and experimentation. Biotechnol. Adv. 2014, 32, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, M.; Sandhya, S.; Prasad, M.N.V.; Freitas, H. Perspectives of plant-associated microbes in heavy metal phytoremediation. Biotechnol. Adv. 2012, 30, 1562–1574. [Google Scholar] [CrossRef] [PubMed]
- Flandroy, L.; Poutahidis, T.; Berg, G.; Clarke, G.; Dao, M.-C.; Decaestecker, E.; Furman, E.; Haahtela, T.; Massart, S.; Plovier, H.; et al. The impact of human activities and lifestyles on the interlinked microbiota and health of humans and of ecosystems. Sci. Total Environ. 2018, 627, 1018–1038. [Google Scholar] [CrossRef] [PubMed]
- Nannipieri, P.; Ascher-Jenull, J.; Ceccherini, M.T.; Pietramellara, G.; Renella, G.; Schloter, M. Beyond microbial diversity for predicting soil functions: A mini review. Pedosphere 2020, 30, 5–17. [Google Scholar] [CrossRef]
- Sasse, J.; Martinoia, E.; Northen, T. Feed your friends: Do plant exudates shape the root microbiome? Trends Plant Sci. 2018, 23, 25–41. [Google Scholar] [CrossRef] [PubMed]
- Rolfe, S.A.; Griffiths, J.; Ton, J. Crying out for help with root exudates: Adaptive mechanisms by which stressed plants assemble health-promoting soil microbiomes. Curr. Opin. Microbiol. 2019, 49, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K. Plant methyl salicylate induces defense responses in the rhizobacterium Bacillus subtilis. Environ. Microbiol. 2015, 17, 1365–1376. [Google Scholar] [CrossRef] [PubMed]
- Korenblum, E.; Dong, Y.; Szymanski, J.; Panda, S.; Jozwiak, A.; Massalha, H.; Meir, S.; Rogachev, I.; Aharoni, A. Rhizosphere microbiome mediates systemic root metabolite exudation by root-to-root signaling. Proc. Natl. Acad. Sci. USA 2020, 117, 3874–3883. [Google Scholar] [CrossRef]
- Camaille, M.; Fabre, N.; Clément, C.; Ait Barka, E. Advances in wheat physiology in response to drought and the role of plant growth promoting rhizobacteria to trigger drought tolerance. Microorganisms 2021, 9, 687. [Google Scholar] [CrossRef]
- Xu, Y.; Chen, Z.; Li, X.; Tan, J.; Liu, F.; Wu, J. Mycorrhizal fungi alter root exudation to cultivate a beneficial microbiome for plant growth. Funct. Ecol. 2023, 37, 664–675. [Google Scholar] [CrossRef]
- Wu, S.; Wu, K.; Shi, L.; Sun, X.; Tan, Q.; Hu, C. Recruitment of specific microbes through exudates affects cadmium activation and accumulation in Brassica napus. J. Hazard. Mater. 2023, 442, 130066. [Google Scholar] [CrossRef] [PubMed]
- Paul, E.A. The nature and dynamics of soil organic matter: Plant inputs, microbial transformations, and organic matter stabilization. Soil Biol. Biochem. 2016, 98, 109–126. [Google Scholar] [CrossRef]
- Zhang, R.; Vivanco, J.M.; Shen, Q. The unseen rhizosphere root-soil-microbe interactions for crop production. Curr. Opin. Microbiol. 2017, 37, 8–14. [Google Scholar] [CrossRef]
- Wang, D.; Wang, C.; Chen, Y.; Xie, Z. Developing Plant-Growth-Promoting Rhizobacteria: A Crucial Approach for Achieving Sustainable Agriculture. Agronomy 2023, 13, 1835. [Google Scholar] [CrossRef]
- Nascimento, F.X.; Urón, P.; Glick, B.R.; Giachini, A.; Rossi, M.J. Genomic analysis of the 1-aminocyclopropane-1-carboxylate deaminase-producing Pseudomonas thivervalensis sc5 reveals its multifaceted roles in soil and in beneficial interactions with plants. Front. Microbiol. 2021, 12, 752288. [Google Scholar] [CrossRef]
- Pantigoso, H.A.; Yuan, J.; He, Y.; Guo, Q.; Vollmer, C.; Vivanco, J.M. Role of root exudates on assimilation of phosphorus in young and old Arabidopsis thaliana plants. PLoS ONE 2020, 15, e0234216. [Google Scholar] [CrossRef]
- Rasmann, S.; Turlings, T.C.J. Root signals that mediate mutualistic interactions in the rhizosphere. Curr. Opin. Plant Biol. 2016, 32, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Yang, C.; Mei, X.; Shen, S.; Raza, W.; Shen, Q.; Xu, Y. The effect of organic acids from tomato root exudates on rhizosphere colonization of Bacillus amyloliquefaciens T-5. Appl. Soil Ecol. 2013, 64, 15–22. [Google Scholar] [CrossRef]
- Nadeem, S.M.; Imran, M.; Naveed, M.; Khan, M.Y.; Ahmad, M.; Zahir, Z.A.; Crowley, D.E. Synergistic use of biochar, compost and plant growth-promoting rhizobacteria for enhancing cucumber growth under water deficit conditions. J. Sci. Food Agric. 2017, 97, 5139–5145. [Google Scholar] [CrossRef]
- Rekha, K.; Ramasamy, M.; Usha, B. Root exudation of organic acids as affected by plant growth-promoting rhizobacteria Bacillus subtilis RR4 in rice. J. Crop Improv. 2020, 34, 571–586. [Google Scholar] [CrossRef]
- Ahkami, A.H.; Allen White, R.; Handakumbura, P.P.; Jansson, C. Rhizosphere engineering: Enhancing sustainable plant ecosystem productivity. Rhizosphere 2017, 3, 233–243. [Google Scholar] [CrossRef]
- da Silva, A.V.; da Silva, M.K.; Sampaio, E.B.T.; Ferreira, L.F.R.; Passarini, M.R.Z.; de Oliveira, V.M.; Rosa, L.H.; Duarte, A.W.F. Benefits of plant growth-promoting symbiotic microbes in climate change era. In Microbiome under Changing Climate; Elsevier: Amsterdam, The Netherlands, 2022; pp. 85–113. [Google Scholar]
- Babalola, O.O.; Akinola, S.A.; Ayangbenro, A.S. Shotgun metagenomic survey of maize soil rhizobiome. Microbiol. Res. Announc. 2020, 9, e00860-20. [Google Scholar] [CrossRef]
- Mathenge, C.; Thuita, M.; Masso, C.; Gweyi-Onyango, J.; Vanlauwe, B. Variability of soybean response to rhizobia inoculant, vermicompost, and a legume-specific fertilizer blend in Siaya County of Kenya. Soil Tillage Res. 2019, 194, 104290. [Google Scholar] [CrossRef] [PubMed]
- Elsheikh, E.A.; El-Keblawy, A.; Mosa, K.A.; Okoh, A.I.; Saadoun, I. Role of endophytes and rhizosphere microbes in promoting the invasion of exotic plants in arid and semi-arid areas: A review. Sustainability 2021, 13, 13081. [Google Scholar] [CrossRef]
- Ayilara, M.S.; Adeleke, B.S.; Adebajo, M.T.; Akinola, S.A.; Fayose, C.A.; Adeyemi, U.T.; Gbadegesin, L.A.; Omole, R.K.; Johnson, R.M.; Edhemuino, M. Remediation by enhanced natural attenuation; an environment-friendly remediation approach. Front. Environ. Sci. 2023, 11, 1182586. [Google Scholar] [CrossRef]
- Gamalero, E.; Bona, E.; Todeschini, V.; Lingua, G. Saline and arid soils: Impact on bacteria, plants, and their interaction. Biology 2020, 9, 116. [Google Scholar] [CrossRef]
- Akinola, S.A.; Adeyemo, R.O.; Adebayo, I.A. Overview and Emergence of Nanobiotechnology in Plants. In Diverse Applications of Nanotechnology in the Biological Sciences; Apple Academic Press: Williston, VT, USA, 2022; pp. 173–197. [Google Scholar]
- Mainka, T.; Weirathmüller, D.; Herwig, C.; Pflügl, S. Potential applications of halophilic microorganisms for biological treatment of industrial process brines contaminated with aromatics. J. Ind. Microbiol. Biotechnol. 2021, 48, kuab015. [Google Scholar] [CrossRef]
- Gupta, A.; Joia, J.; Sood, A.; Sood, R.; Sidhu, C.; Kaur, G. Microbes as potential tool for remediation of heavy metals: A review. Microb. Biochem. Technol. 2016, 8, 364–372. [Google Scholar] [CrossRef]
- Ayilara, M.S.; Akinola, S.A.; Adebajo, M.T. Benefits and Drawbacks of Microbial Inoculant in Terms of Human Health and the Environment. In Microbial Inoculants: Applications for Sustainable Agriculture; Springer: Berlin/Heidelberg, Germany, 2024; pp. 411–435. [Google Scholar]
- Oyebamiji, Y.O.; Nahar, S.; Shamsudin, N.A.; Salisu, M.A.; Akinola, S.A.; Adebayo, I.A.; Arsad, H. The role of arbuscular mycorrhizal fungi in alleviating heat and drought stress in vegetables. Egypt. J. Bot. 2024, 64, 497–505. [Google Scholar] [CrossRef]
- Mukhtar, T.; Ali, F.; Rafique, M.; Ali, J.; Afridi, M.S.; Smith, D.; Mehmood, S.; Amna; Souleimanov, A.; Jellani, G. Biochemical characterization and potential of Bacillus safensis strain scal1 to mitigate heat stress in Solanum lycopersicum L. J. Plant Growth Regul. 2023, 42, 523–538. [Google Scholar] [CrossRef]
- Mukhtar, S.; Mirza, B.S.; Mehnaz, S.; Mirza, M.S.; Mclean, J.; Malik, K.A. Impact of soil salinity on the microbial structure of halophyte rhizosphere microbiome. World J. Microbiol. Biotechnol. 2018, 34, 136. [Google Scholar] [CrossRef] [PubMed]
- Akinola, S.A.; Ayangbenro, A.S.; Babalola, O.O. Pedological factors as drivers of archaeal and fungal communities in maize rhizosphere: A shotgun metagenomic sequencing approach. SN Appl. Sci. 2023, 5, 351. [Google Scholar] [CrossRef]
- Ladau, J.; Shi, Y.; Jing, X.; He, J.-S.; Chen, L.; Lin, X.; Fierer, N.; Gilbert, J.A.; Pollard, K.S.; Chu, H. Climate change will lead to pronounced shifts in the diversity of soil microbial communities. bioRxiv 2018. [Google Scholar] [CrossRef]
- Mahmud, K.; Missaoui, A.; Lee, K.; Ghimire, B.; Presley, H.W.; Makaju, S. Rhizosphere microbiome manipulation for sustainable crop production. Curr. Plant Biol. 2021, 27, 100210. [Google Scholar] [CrossRef]
- Mukhtar, S.; Mehnaz, S.; Mirza, M.S.; Malik, K.A. Isolation and characterization of bacteria associated with the rhizosphere of halophytes (Salsola stocksii and Atriplex amnicola) for production of hydrolytic enzymes. Braz. J. Microbiol. 2019, 50, 85–97. [Google Scholar] [CrossRef]
- Ayilara, M.S.; Adeleke, B.S.; Akinola, S.A.; Fayose, C.A.; Adeyemi, U.T.; Gbadegesin, L.A.; Omole, R.K.; Johnson, R.M.; Uthman, Q.O.; Babalola, O.O. Biopesticides as a promising alternative to synthetic pesticides: A case for microbial pesticides, phytopesticides, and nanobiopesticides. Front. Microbiol. 2023, 14, 1040901. [Google Scholar] [CrossRef] [PubMed]
- Akinola, S.A.; Babalola, O.O. The importance of adverse soil microbiomes in the light of omics: Implications for food safety. Plant Soil Environ. 2020, 66, 421–430. [Google Scholar] [CrossRef]
- Dardanelli, M.S.; de Cordoba, F.J.F.; Espuny, M.R.; Carvajal, M.A.R.; Díaz, M.E.S.; Serrano, A.M.G.; Okon, Y.; Megías, M. Effect of Azospirillum brasilense coinoculated with Rhizobium on Phaseolus vulgaris flavonoids and Nod factor production under salt stress. Soil Biol. Biochem. 2008, 40, 2713–2721. [Google Scholar] [CrossRef]
- Del Amor, F.M.; Cuadra-Crespo, P. Plant growth-promoting bacteria as a tool to improve salinity tolerance in sweet pepper. Funct. Plant Biol. 2011, 39, 82–90. [Google Scholar] [CrossRef]
- Ait Barka, E.; Nowak, J.; Clément, C. Enhancement of chilling resistance of inoculated grapevine plantlets with a plant growth-promoting rhizobacterium, Burkholderia phytofirmans strain PsJN. Appl. Environ. Microbiol. 2006, 72, 7246–7252. [Google Scholar] [CrossRef]
- Marulanda, A.; Azcón, R.; Chaumont, F.; Ruiz-Lozano, J.M.; Aroca, R. Regulation of plasma membrane aquaporins by inoculation with a Bacillus megaterium strain in maize (Zea mays L.) plants under unstressed and salt-stressed conditions. Planta 2010, 232, 533–543. [Google Scholar] [CrossRef] [PubMed]
- Vaishnav, A.; Kumari, S.; Jain, S.; Varma, A.; Tuteja, N.; Choudhary, D.K. PGPR-mediated expression of salt tolerance gene in soybean through volatiles under sodium nitroprusside. J. Basic Microbiol. 2016, 56, 1274–1288. [Google Scholar] [CrossRef] [PubMed]
- Singh, S. A review on possible elicitor molecules of cyanobacteria: Their role in improving plant growth and providing tolerance against biotic or abiotic stress. J. Appl. Microbiol. 2014, 117, 1221–1244. [Google Scholar] [CrossRef]
- Jha, Y.; Sablok, G.; Subbarao, N.; Sudhakar, R.; Fazil, M.T.; Subramanian, R.; Squartini, A.; Kumar, S. Bacterial-induced expression of RAB18 protein in Orzya sativa salinity stress and insights into molecular interaction with GTP ligand. J. Mol. Recognit. Interdiscip. J. 2014, 27, 521–527. [Google Scholar] [CrossRef]
- Naveed, M.; Mitter, B.; Reichenauer, T.G.; Wieczorek, K.; Sessitsch, A. Increased drought stress resilience of maize through endophytic colonization by Burkholderia phytofirmans PsJN and Enterobacter sp. FD17. Environ. Exp. Bot. 2014, 97, 30–39. [Google Scholar] [CrossRef]
- Cho, S.M.; Kang, B.R.; Han, S.H.; Anderson, A.J.; Park, J.-Y.; Lee, Y.-H.; Cho, B.H.; Yang, K.-Y.; Ryu, C.-M.; Kim, Y.C. 2R, 3R-butanediol, a bacterial volatile produced by Pseudomonas chlororaphis O6, is involved in induction of systemic tolerance to drought in Arabidopsis thaliana. Mol. Plant-Microbe Interact. 2008, 21, 1067–1075. [Google Scholar] [CrossRef]
- Lu, X.; Jin, C.; Yang, J.; Liu, Q.; Wu, S.; Li, D.; Guan, Y.; Cai, Y. Prenatal and lactational lead exposure enhanced oxidative stress and altered apoptosis status in offspring rats’ hippocampus. Biol. Trace Elem. Res. 2013, 151, 75–84. [Google Scholar] [CrossRef]
- Islam, F.; Yasmeen, T.; Ali, Q.; Ali, S.; Arif, M.S.; Hussain, S.; Rizvi, H. Influence of Pseudomonas aeruginosa as PGPR on oxidative stress tolerance in wheat under Zn stress. Ecotoxicol. Environ. Saf. 2014, 104, 285–293. [Google Scholar] [CrossRef]
- Bhattacharyya, P.N.; Jha, D.K. Plant growth-promoting rhizobacteria (PGPR): Emergence in agriculture. World J. Microbiol. Biotechnol. 2012, 28, 1327–1350. [Google Scholar] [CrossRef]
- Giordano, W.; Hirsch, A.M. The expression of MaEXP1, a Melilotus alba expansin gene, is upregulated during the sweetclover-Sinorhizobium meliloti interaction. Mol. Plant-Microbe Interact. 2004, 17, 613–622. [Google Scholar] [CrossRef]
- AbuQamar, S.F.; El-Saadony, M.T.; Saad, A.M.; Desoky, E.S.M.; Elrys, A.S.; Abd El-Mageed, T.A.; Semida, W.M.; Abdelkhalik, A.; Mosa, W.F.; Al Kafaas, S.S.; et al. Halotolerant plant growth-promoting rhizobacteria improve soil fertility and plant salinity tolerance for sustainable agriculture—A review. Plant Stress 2024, 12, 100482. [Google Scholar] [CrossRef]
- Akinola, S.A.; Babalola, O.O. The fungal and archaeal community within plant rhizosphere: A review on their contribution to crop safety. J. Plant Nutr. 2021, 44, 600–618. [Google Scholar] [CrossRef]
- Muhammad, M.; Waheed, A.; Wahab, A.; Majeed, M.; Nazim, M.; Liu, Y.-H.; Li, L.; Li, W.-J. Soil salinity and drought tolerance: An evaluation of plant growth, productivity, microbial diversity, and amelioration strategies. Plant Stress 2023, 11, 100319. [Google Scholar] [CrossRef]
- Singh, P.; Singh, R.K.; Guo, D.-J.; Sharma, A.; Singh, R.N.; Li, D.-P.; Malviya, M.K.; Song, X.-P.; Lakshmanan, P.; Yang, L.-T. Whole genome analysis of sugarcane root-associated endophyte Pseudomonas aeruginosa B18—A plant growth-promoting bacterium with antagonistic potential against Sporisorium scitamineum. Front. Microbiol. 2021, 12, 104. [Google Scholar] [CrossRef] [PubMed]
- Gan, H.M.; McGroty, S.E.; Chew, T.H.; Chan, K.G.; Buckley, L.J.; Savka, M.A.; Hudson, A.O. Whole-genome sequence of Enterobacter sp. strain SST3, an endophyte isolated from Jamaican sugarcane (Saccharum sp.) stalk tissue. J. Bacteriol. 2012, 194, 5981–5982. [Google Scholar] [CrossRef]
- Viscardi, S.; Ventorino, V.; Duran, P.; Maggio, A.; De Pascale, S.; Mora, M.; Pepe, O. Assessment of plant growth promoting activities and abiotic stress tolerance of Azotobacter chroococcum strains for a potential use in sustainable agriculture. J. Soil Sci. Plant Nutr. 2016, 16, 848–863. [Google Scholar] [CrossRef]
- Yarra, R. The wheat NHX gene family: Potential role in improving salinity stress tolerance of plants. Plant Gene 2019, 18, 100178. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.; Anee, T.I.; Parvin, K.; Nahar, K.; Mahmud, J.A.; Fujita, M. Regulation of ascorbate-glutathione pathway in mitigating oxidative damage in plants under abiotic stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.-J. The role of the plant antioxidant system in drought tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef]
- Ouledali, S.; Ennajeh, M.; Ferrandino, A.; Khemira, H.; Schubert, A.; Secchi, F. Influence of arbuscular mycorrhizal fungi inoculation on the control of stomata functioning by abscisic acid (ABA) in drought-stressed olive plants. South Afr. J. Bot. 2019, 121, 152–158. [Google Scholar] [CrossRef]
- Rizvi, A.; Khan, M.S. Biotoxic impact of heavy metals on growth, oxidative stress and morphological changes in root structure of wheat (Triticum aestivum L.) and stress alleviation by Pseudomonas aeruginosa strain CPSB1. Chemosphere 2017, 185, 942–952. [Google Scholar] [CrossRef] [PubMed]
- Balestrini, R.; Lumini, E. Focus on mycorrhizal symbioses. Appl. Soil Ecol. 2018, 123, 299–304. [Google Scholar] [CrossRef]
- Kapoor, R.; Evelin, H.; Mathur, P.; Giri, B. Arbuscular mycorrhiza: Approaches for abiotic stress tolerance in crop plants for sustainable agriculture. In Plant Acclimation to Environmental Stress; Springer: Berlin/Heidelberg, Germany, 2012; pp. 359–401. [Google Scholar]
- Arshad, M.; Saleem, M.; Hussain, S. Perspectives of bacterial ACC deaminase in phytoremediation. Trends Biotechnol. 2007, 25, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Camerini, S.; Senatore, B.; Lonardo, E.; Imperlini, E.; Bianco, C.; Moschetti, G.; Rotino, G.L.; Campion, B.; Defez, R. Introduction of a novel pathway for IAA biosynthesis to rhizobia alters vetch root nodule development. Arch. Microbiol. 2008, 190, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R. Plant growth-promoting bacteria: Mechanisms and applications. Scientifica 2012, 2012, 963401. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.K.; Ding, Y.Z.; Feng, R.W.; Wang, R.G.; Xu, Y.M.; Chen, C.; Wei, X.L.; Chen, W.M. Burkholderia metalliresistens sp. nov., a multiple metal-resistant and phosphate-solubilising species isolated from heavy metal-polluted soil in Southeast China. Antonie Van Leeuwenhoek 2015, 107, 1591–1598. [Google Scholar] [CrossRef] [PubMed]
- Patten, C.L.; Glick, B.R. Bacterial biosynthesis of indole-3-acetic acid. Can. J. Microbiol. 1996, 42, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Mathew, B.T.; Torky, Y.; Amin, A.; Mourad, A.-H.I.; Ayyash, M.M.; El-Keblawy, A.; Hilal-Alnaqbi, A.; AbuQamar, S.F.; El-Tarabily, K.A. Halotolerant marine rhizosphere-competent actinobacteria promote Salicornia bigelovii growth and seed production using seawater irrigation. Front. Microbiol. 2020, 11, 552. [Google Scholar] [CrossRef]
- Neubauer, U.; Furrer, G.; Schulin, R. Heavy metal sorption on soil minerals affected by the siderophore desferrioxamine B: The role of Fe (III)(hydr) oxides and dissolved Fe (III). Eur. J. Soil Sci. 2002, 53, 45–55. [Google Scholar] [CrossRef]
- El-Tarabily, K.A.; ElBaghdady, K.Z.; AlKhajeh, A.S.; Ayyash, M.M.; Aljneibi, R.S.; El-Keblawy, A.; AbuQamar, S.F. Polyamine-producing actinobacteria enhance biomass production and seed yield in Salicornia bigelovii. Biol. Fertil. Soils 2020, 56, 499–519. [Google Scholar] [CrossRef]
- Ghosh, U.D.; Saha, C.; Maiti, M.; Lahiri, S.; Ghosh, S.; Seal, A.; MitraGhosh, M. Root associated iron oxidizing bacteria increase phosphate nutrition and influence root to shoot partitioning of iron in tolerant plant Typha angustifolia. Plant Soil 2014, 381, 279–295. [Google Scholar] [CrossRef]
- Yadav, J.; Verma, J.P.; Jaiswal, D.K.; Kumar, A. Evaluation of PGPR and different concentration of phosphorus level on plant growth, yield and nutrient content of rice (Oryza sativa). Ecol. Eng. 2014, 62, 123–128. [Google Scholar]
- Ali, S.; Glick, B.R. Plant–bacterial interactions in management of plant growth under abiotic stresses. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2019; pp. 21–45. [Google Scholar]
- Igiehon, N.O.; Babalola, O.O.; Aremu, B.R. Genomic insights into plant growth promoting rhizobia capable of enhancing soybean germination under drought stress. BMC Microbiol. 2019, 19, 159. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, P.; Parveen, H.; Gangola, S.; Kumar, G.; Bhatt, P.; Chaudhary, A. Plant growth-promoting rhizobacteria and their application in sustainable crop production. In Microbial Technology for Sustainable Environment; Springer: Singapore, 2021; pp. 217–234. [Google Scholar]
- Fahsi, N.; Mahdi, I.; Mesfioui, A.; Biskri, L.; Allaoui, A. Plant growth-promoting rhizobacteria isolated from the Jujube (Ziziphus lotus) plant enhance wheat growth, Zn uptake, and heavy metal tolerance. Agriculture 2021, 11, 316. [Google Scholar] [CrossRef]
- Zahid, G.; Iftikhar, S.; Shimira, F.; Ahmad, H.M.; Kaçar, Y.A. An overview and recent progress of plant growth regulators (PGRs) in the mitigation of abiotic stresses in fruits: A review. Sci. Hortic. 2023, 309, 111621. [Google Scholar] [CrossRef]
- Ghazi, A.; Atia, E.; Elsakhawy, T. Evaluation of an endophytic plant growth-promoting bacterium, Klebsiella variicola, in mitigation of salt stress in tuberose (Polianthes tuberosa L.). J. Hortic. Sci. Biotechnol. 2021, 96, 770–782. [Google Scholar] [CrossRef]
- Ulrich, K.; Kube, M.; Becker, R.; Schneck, V.; Ulrich, A. Genomic analysis of the endophytic Stenotrophomonas strain 169 reveals features related to plant-growth promotion and stress tolerance. Front. Microbiol. 2021, 12, 687463. [Google Scholar] [CrossRef]
- Lu, L.; Chang, M.; Han, X.; Wang, Q.; Wang, J.; Yang, H.; Guan, Q.; Dai, S. Beneficial effects of endophytic Pantoea ananatis with ability to promote rice growth under saline stress. J. Appl. Microbiol. 2021, 131, 1919–1931. [Google Scholar] [CrossRef]
- Annapurna, K.; Govindasamy, V.; Ajit, V.; Choudhary, D. Mitigation of drought stress in wheat crop by drought tolerant endophytic bacterial isolates. Vegetos. 2019, 32, 486–493. [Google Scholar]
- Gamalero, E.; Favale, N.; Bona, E.; Novello, G.; Cesaro, P.; Massa, N.; Glick, B.R.; Orozco-Mosqueda, M.d.C.; Berta, G.; Lingua, G. Screening of bacterial endophytes able to promote plant growth and increase salinity tolerance. Appl. Sci. 2020, 10, 5767. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Al-Ghamdi, A.A.; Ali, H.M.; Alayafi, A.A. Azospirillum lipoferum FK1 confers improved salt tolerance in chickpea (Cicer arietinum L.) by modulating osmolytes, antioxidant machinery and stress-related genes expression. Environ. Exp. Bot. 2019, 159, 55–65. [Google Scholar] [CrossRef]
- Tahir, M.; Ahmad, I.; Shahid, M.; Shah, G.M.; Farooq, A.B.U.; Akram, M.; Tabassum, S.A.; Naeem, M.A.; Khalid, U.; Ahmad, S. Regulation of antioxidant production, ion uptake and productivity in potato (Solanum tuberosum L.) plant inoculated with growth promoting salt tolerant Bacillus strains. Ecotoxicol. Environ. Saf. 2019, 178, 33–42. [Google Scholar] [CrossRef]
- Afridi, M.S.; Mahmood, T.; Salam, A.; Mukhtar, T.; Mehmood, S.; Ali, J.; Khatoon, Z.; Bibi, M.; Javed, M.T.; Sultan, T. Induction of tolerance to salinity in wheat genotypes by plant growth promoting endophytes: Involvement of ACC deaminase and antioxidant enzymes. Plant Physiol. Biochem. 2019, 139, 569–577. [Google Scholar] [CrossRef]
- Vimal, S.R.; Patel, V.K.; Singh, J.S. Plant growth promoting Curtobacterium albidum strain SRV4: An agriculturally important microbe to alleviate salinity stress in paddy plants. Ecol. Indic. 2019, 105, 553–562. [Google Scholar] [CrossRef]
- Li, H.; Lei, P.; Pang, X.; Li, S.; Xu, H.; Xu, Z.; Feng, X. Enhanced tolerance to salt stress in canola (Brassica napus L.) seedlings inoculated with the halotolerant Enterobacter cloacae HSNJ4. Appl. Soil Ecol. 2017, 119, 26–34. [Google Scholar] [CrossRef]
- Sarkar, A.; Pramanik, K.; Mitra, S.; Soren, T.; Maiti, T.K. Enhancement of growth and salt tolerance of rice seedlings by ACC deaminase-producing Burkholderia sp. MTCC 12259. J. Plant Physiol. 2018, 231, 434–442. [Google Scholar] [CrossRef]
- Bhattacharyya, D.; Lee, Y.H. A cocktail of volatile compounds emitted from Alcaligenes faecalis JBCS1294 induces salt tolerance in Arabidopsis thaliana by modulating hormonal pathways and ion transporters. J. Plant Physiol. 2017, 214, 64–73. [Google Scholar] [CrossRef]
- Samaddar, S.; Chatterjee, P.; Choudhury, A.R.; Ahmed, S.; Sa, T. Interactions between Pseudomonas spp. and their role in improving the red pepper plant growth under salinity stress. Microbiol. Res. 2019, 219, 66–73. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Alaraidh, I.A.; Alsahli, A.A.; Alamri, S.A.; Ali, H.M.; Alayafi, A.A. Bacillus firmus (SW5) augments salt tolerance in soybean (Glycine max L.) by modulating root system architecture, antioxidant defense systems and stress-responsive genes expression. Plant Physiol. Biochem. 2018, 132, 375–384. [Google Scholar] [CrossRef]
- Hmaeid, N.; Wali, M.; Mahmoud, O.M.-B.; Pueyo, J.J.; Ghnaya, T.; Abdelly, C. Efficient rhizobacteria promote growth and alleviate NaCl-induced stress in the plant species Sulla carnosa. Appl. Soil Ecol. 2019, 133, 104–113. [Google Scholar] [CrossRef]
- Sapre, S.; Gontia-Mishra, I.; Tiwari, S. Klebsiella sp. confers enhanced tolerance to salinity and plant growth promotion in oat seedlings (Avena sativa). Microbiol. Res. 2018, 206, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Abdiev, A.; Khaitov, B.; Toderich, K.; Park, K.W. Growth, nutrient uptake and yield parameters of chickpea (Cicer arietinum L.) enhance by Rhizobium and Azotobacter inoculations in saline soil. J. Plant Nutr. 2019, 42, 2703–2714. [Google Scholar] [CrossRef]
- Khan, M.A.; Hamayun, M.; Asaf, S.; Khan, M.; Yun, B.-W.; Kang, S.-M.; Lee, I.-J. Rhizospheric Bacillus spp. rescues plant growth under salinity stress via regulating gene expression, endogenous hormones, and antioxidant system of Oryza sativa L. Front. Plant Sci. 2021, 12, 665590. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.; Gupta, S. ACC deaminase producing bacteria with multifarious plant growth promoting traits alleviates salinity stress in French bean (Phaseolus vulgaris) plants. Front. Microbiol. 2019, 10, 1506. [Google Scholar]
- Tolba, S.T.; Ibrahim, M.; Amer, E.A.; Ahmed, D.A. First insights into salt tolerance improvement of Stevia by plant growth-promoting Streptomyces species. Arch. Microbiol. 2019, 201, 1295–1306. [Google Scholar] [CrossRef]
- Wahdan, S.F.M.; Tanunchai, B.; Wu, Y.T.; Sansupa, C.; Schädler, M.; Dawoud, T.M.; Buscot, F.; Purahong, W. Deciphering Trifolium pratense L. holobiont reveals a microbiome resilient to future climate changes. Microbiologyopen 2021, 10, e1217. [Google Scholar] [CrossRef]
- Armada, E.; Azcón, R.; López-Castillo, O.M.; Calvo-Polanco, M.; Ruiz-Lozano, J.M. Autochthonous arbuscular mycorrhizal fungi and Bacillus thuringiensis from a degraded Mediterranean area can be used to improve physiological traits and performance of a plant of agronomic interest under drought conditions. Plant Physiol. Biochem. 2015, 90, 64–74. [Google Scholar] [CrossRef]
- Lastochkina, O.; Ivanov, S.; Petrova, S.; Garshina, D.; Lubyanova, A.; Yuldashev, R.; Kuluev, B.; Zaikina, E.; Maslennikova, D.; Allagulova, C. Role of endogenous salicylic acid as a hormonal intermediate in the bacterial endophyte Bacillus subtilis-induced protection of wheat genotypes contrasting in drought susceptibility under dehydration. Plants 2022, 11, 3365. [Google Scholar] [CrossRef]
- Oliveira, T.C.; Cabral, J.S.R.; Santana, L.R.; Tavares, G.G.; Santos, L.D.S.; Paim, T.P.; Müller, C.; Silva, F.G.; Costa, A.C.; Souchie, E.L. The arbuscular mycorrhizal fungus Rhizophagus clarus improves physiological tolerance to drought stress in soybean plants. Sci. Rep. 2022, 12, 9044. [Google Scholar] [CrossRef]
- Jatan, R.; Tiwari, S.; Asif, M.H.; Lata, C. Genome-wide profiling reveals extensive alterations in Pseudomonas putida-mediated miRNAs expression during drought stress in chickpea (Cicer arietinum L.). Environ. Exp. Bot. 2019, 157, 217–227. [Google Scholar] [CrossRef]
- Chandra, D.; Srivastava, R.; Glick, B.R.; Sharma, A.K. Rhizobacteria producing ACC deaminase mitigate water-stress response in finger millet (Eleusine coracana (L.) Gaertn.). 3 Biotech 2020, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Kour, D.; Rana, K.L.; Kaur, T.; Sheikh, I.; Yadav, A.N.; Kumar, V.; Dhaliwal, H.S.; Saxena, A.K. Microbe-mediated alleviation of drought stress and acquisition of phosphorus in great millet (Sorghum bicolour L.) by drought-adaptive and phosphorus-solubilizing microbes. Biocatal. Agric. Biotechnol. 2020, 23, 101501. [Google Scholar] [CrossRef]
- Chandra, D.; Srivastava, R.; Glick, B.R.; Sharma, A.K. Drought-tolerant Pseudomonas spp. improve the growth performance of finger millet (Eleusine coracana (L.) Gaertn.) under non-stressed and drought-stressed conditions. Pedosphere 2018, 28, 227–240. [Google Scholar] [CrossRef]
- Ansari, F.; Jabeen, M.; Ahmad, I. Pseudomonas azotoformans FAP5, a novel biofilm-forming PGPR strain, alleviates drought stress in wheat plant. Int. J. Environ. Sci. Technol. 2021, 18, 3855–3870. [Google Scholar] [CrossRef]
- Chandra, D.; Srivastava, R.; Gupta, V.V.; Franco, C.M.; Sharma, A.K. Evaluation of ACC-deaminase-producing rhizobacteria to alleviate water-stress impacts in wheat (Triticum aestivum L.) plants. Can. J. Microbiol. 2019, 65, 387–403. [Google Scholar] [CrossRef]
- Dubey, A.; Saiyam, D.; Kumar, A.; Hashem, A.; Abd_Allah, E.F.; Khan, M.L. Bacterial root endophytes: Characterization of their competence and plant growth promotion in soybean (Glycine max (L.) Merr.) under drought stress. Int. J. Environ. Res. Public Health 2021, 18, 931. [Google Scholar] [CrossRef] [PubMed]
- Chukwuneme, C.F.; Babalola, O.O.; Kutu, F.R.; Ojuederie, O.B. Characterization of actinomycetes isolates for plant growth promoting traits and their effects on drought tolerance in maize. J. Plant Interact. 2020, 15, 93–105. [Google Scholar] [CrossRef]
- El-Meihy, R.M.; Abou-Aly, H.E.; Youssef, A.M.; Tewfike, T.A.; El-Alkshar, E.A. Efficiency of heavy metals-tolerant plant growth promoting bacteria for alleviating heavy metals toxicity on sorghum. Environ. Exp. Bot. 2019, 162, 295–301. [Google Scholar] [CrossRef]
- Rizvi, A.; Khan, M.S. Heavy metal induced oxidative damage and root morphology alterations of maize (Zea mays L.) plants and stress mitigation by metal tolerant nitrogen fixing Azotobacter chroococcum. Ecotoxicol. Environ. Saf. 2018, 157, 9–20. [Google Scholar] [CrossRef]
- Ke, T.; Guo, G.; Liu, J.; Zhang, C.; Tao, Y.; Wang, P.; Xu, Y.; Chen, L. Improvement of the Cu and Cd phytostabilization efficiency of perennial ryegrass through the inoculation of three metal-resistant PGPR strains. Nviron. Pollut. 2021, 271, 116314. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, N.; Ilyas, N.; Yasmin, H.; Sayyed, R.; Hasnain, Z.; Elsayed, E.A.; El Enshasy, H.A. Role of Bacillus cereus in improving the growth and phytoextractability of Brassica nigra (L.) K. Koch in chromium contaminated soil. Molecules 2021, 26, 1569. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Xu, M.; Wei, Q.; Tang, M.; Guan, L.; Lou, L.; Xu, X.; Hu, Z.; Chen, Y.; Shen, Z. Promotion of growth and phytoextraction of cadmium and lead in Solanum nigrum L. mediated by plant-growth-promoting rhizobacteria. Ecotoxicol. Environ. Saf. 2020, 205, 111333. [Google Scholar] [CrossRef]
- Zafar-ul-Hye, M.; Tahzeeb-ul-Hassan, M.; Wahid, A.; Danish, S.; Khan, M.J.; Fahad, S.; Brtnicky, M.; Hussain, G.S.; Battaglia, M.L.; Datta, R. Compost mixed fruits and vegetable waste biochar with ACC deaminase rhizobacteria can minimize lead stress in mint plants. Sci. Rep. 2021, 11, 6606. [Google Scholar] [CrossRef] [PubMed]
- Awan, S.A.; Ilyas, N.; Khan, I.; Raza, M.A.; Rehman, A.U.; Rizwan, M.; Rastogi, A.; Tariq, R.; Brestic, M. Bacillus siamensis reduces cadmium accumulation and improves growth and antioxidant defense system in two wheat (Triticum aestivum L.) varieties. Plants 2020, 9, 878. [Google Scholar] [CrossRef] [PubMed]
- Saleem, M.; Nawaz, F.; Hussain, M.B.; Ikram, R.M. Comparative effects of individual and consortia plant growth promoting bacteria on physiological and enzymatic mechanisms to confer drought tolerance in maize (Zea mays L.). J Soil Sci. Plant Nutr. 2021, 21, 3461–3476. [Google Scholar] [CrossRef]
- Mousavi, S.M.; Motesharezadeh, B.; Hosseini, H.M.; Alikhani, H.; Zolfaghari, A.A. Root-induced changes of Zn and Pb dynamics in the rhizosphere of sunflower with different plant growth promoting treatments in a heavily contaminated soil. Ecotoxicol. Environ. Saf. 2018, 147, 206–216. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.N. Agriculturally important microbiomes: Biodiversity and multifarious PGP attributes for amelioration of diverse abiotic stresses in crops for sustainable agriculture. Biomed. J. Sci. Technol. Res. 2017, 1, 861–864. [Google Scholar]
- Etesami, H.; Maheshwari, D.K. Use of plant growth promoting rhizobacteria (PGPRs) with multiple plant growth promoting traits in stress agriculture: Action mechanisms and future prospects. Ecotoxicol. Environ. Saf. 2018, 156, 225–246. [Google Scholar] [CrossRef]
- Ma, Y.; Rajkumar, M.; Moreno, A.; Zhang, C.; Freitas, H. Serpentine endophytic bacterium Pseudomonas azotoformans ASS1 accelerates phytoremediation of soil metals under drought stress. Chemosphere 2017, 185, 75–85. [Google Scholar] [CrossRef]
- Kumar, U.; Kaviraj, M.; Rout, S.; Chakraborty, K.; Swain, P.; Nayak, P.; Nayak, A. Combined application of ascorbic acid and endophytic N-fixing Azotobacter chroococcum Avi2 modulates photosynthetic efficacy, antioxidants and growth-promotion in rice under moisture deficit stress. Microbiol. Res. 2021, 250, 126808. [Google Scholar] [CrossRef] [PubMed]
- Gagné-Bourque, F.; Bertrand, A.; Claessens, A.; Aliferis, K.A.; Jabaji, S. Alleviation of drought stress and metabolic changes in timothy (Phleum pratense L.) colonized with Bacillus subtilis B26. Front. Plant Sci. 2016, 7, 584. [Google Scholar] [CrossRef] [PubMed]
- Cherni, M.; Ferjani, R.; Mapelli, F.; Boudabous, A.; Borin, S.; Ouzari, H.-I. Soil parameters drive the diversity of Citrus sinensis rhizosphere microbiota which exhibits a potential in plant drought stress alleviation. Appl. Soil Ecol. 2019, 135, 182–193. [Google Scholar] [CrossRef]
- Valetti, L.; Iriarte, L.; Fabra, A. Growth promotion of rapeseed (Brassica napus) associated with the inoculation of phosphate solubilizing bacteria. Appl. Soil Ecol. 2018, 132, 1–10. [Google Scholar] [CrossRef]
- Silva, E.R.; Zoz, J.; Oliveira, C.E.S.; Zuffo, A.M.; Steiner, F.; Zoz, T.; Vendruscolo, E.P. Can co-inoculation of Bradyrhizobium and Azospirillum alleviate adverse effects of drought stress on soybean (Glycine max L. Merrill.)? Arch. Microbiol. 2019, 201, 325–335. [Google Scholar] [CrossRef]
- Arora, S.; Jha, P.N. Drought-tolerant Enterobacter bugandensis WRS7 induces systemic tolerance in Triticum aestivum L. (wheat) under drought conditions. J. Plant Growth Regul. 2023, 42, 7715–7730. [Google Scholar] [CrossRef]
- Kour, D.; Khan, S.S.; Kour, H.; Kaur, T.; Devi, R.; Rai, A.K.; Yadav, A.N. ACC deaminase producing phytomicrobiomes for amelioration of abiotic stresses in plants for agricultural sustainability. J. Plant Growth Regul. 2023, 43, 963–985. [Google Scholar] [CrossRef]
- Abdelkrim, S.; Abid, G.; Chaieb, O.; Taamalli, W.; Mannai, K.; Louati, F.; Jebara, M.; Jebara, S.H. Plant growth promoting rhizobacteria modulates the antioxidant defense and the expression of stress-responsive genes providing Pb accumulation and tolerance of grass pea. Environ. Sci. Pollut. Res. 2023, 30, 10789–10802. [Google Scholar] [CrossRef]
- Noori, F.; Etesami, H.; Noori, S.; Forouzan, E.; Jouzani, G.S.; Malboobi, M.A. Whole genome sequence of Pantoea agglomerans ANP8, a salinity and drought stress–resistant bacterium isolated from alfalfa (Medicago sativa L.) root nodules. Biotechnol. Rep. 2021, 29, e00600. [Google Scholar] [CrossRef]
- Lastochkina, O.; Aliniaeifard, S.; Garshina, D.; Garipova, S.; Pusenkova, L.; Allagulova, C.; Fedorova, K.; Baymiev, A.; Koryakov, I.; Sobhani, M. Seed priming with endophytic Bacillus subtilis strain-specifically improves growth of Phaseolus vulgaris plants under normal and salinity conditions and exerts anti-stress effect through induced lignin deposition in roots and decreased oxidative and osmotic damages. J. Plant Physiol. 2021, 263, 153462. [Google Scholar]
- Nohwar, N.; Khandare, R.V.; Desai, N.S. Isolation and characterization of salinity tolerant nitrogen fixing bacteria from Sesbania sesban (L) root nodules. Biocataly. Agric. Biotechnol. 2019, 21, 101325. [Google Scholar] [CrossRef]
- Khan, M.; Asaf, S.; Khan, A.; Adhikari, A.; Jan, R.; Ali, S.; Imran, M.; Kim, K.M.; Lee, I.J. Plant growth-promoting endophytic bacteria augment growth and salinity tolerance in rice plants. Plant Biol. 2020, 22, 850–862. [Google Scholar] [CrossRef]
- Bhatta, D.; Adhikari, A.; Kang, S.-M.; Kwon, E.-H.; Jan, R.; Kim, K.-M.; Lee, I.-J. Hormones and the antioxidant transduction pathway and gene expression, mediated by Serratia marcescens DB1, lessen the lethality of heavy metals (As, Ni, and Cr) in Oryza sativa L. Ecotoxicol. Environ. Saf. 2023, 263, 115377. [Google Scholar] [CrossRef]
- Sapkota, S.; Harris-Shultz, K.R.; Strickland, T.C.; Anderson, W.F. Identification of cultured and diazotrophic bacterial endophytes in warm-season grasses. PhytoFrontiers 2023, 3, 411–419. [Google Scholar] [CrossRef]
- Pérez-García, A.; Romero, D.; de Vicente, A. Plant protection and growth stimulation by microorganisms: Biotechnological applications of Bacilli in agriculture. Curr. Opin. Biotechnol. 2011, 22, 187–193. [Google Scholar] [CrossRef]
- Lal, S.; Ratna, S.; Said, O.B.; Kumar, R. Biosurfactant and exopolysaccharide-assisted rhizobacterial technique for the remediation of heavy metal contaminated soil: An advancement in metal phytoremediation technology. Environ. Technol. Innov. 2018, 10, 243–263. [Google Scholar] [CrossRef]
- Zhu, Y.; Xu, F.; Liu, Q.; Chen, M.; Liu, X.; Wang, Y.; Sun, Y.; Zhang, L. Nanomaterials and plants: Positive effects, toxicity and the remediation of metal and metalloid pollution in soil. Sci. Total Environ. 2019, 662, 414–421. [Google Scholar] [CrossRef]
- Kaushal, M.; Wani, S.P. Plant-growth-promoting rhizobacteria: Drought stress alleviators to ameliorate crop production in drylands. Ann. Microbiol. 2016, 66, 35–42. [Google Scholar] [CrossRef]
- Oyedoh, O.P.; Yang, W.; Dhanasekaran, D.; Santoyo, G.; Glick, B.R.; Babalola, O.O. Rare rhizo-Actinomycetes: A new source of agroactive metabolites. Biotechnol. Adv. 2023, 67, 108205. [Google Scholar] [CrossRef] [PubMed]
- Asif, M.; Ahmad, R.; Pervez, A.; Al Farraj, D.A.; Elshikh, M.S.; Shahzad, M.; Irshad, U.; Abbasi, A.M. Combination of melatonin and plant growth promoting rhizobacteria improved the growth of Spinacia oleracea L. under the arsenic and cadmium stresses. Physiol. Mol. Plant Pathol. 2023, 127, 102097. [Google Scholar] [CrossRef]
- Wu, J.; Kamal, N.; Hao, H.; Qian, C.; Liu, Z.; Shao, Y.; Zhong, X.; Xu, B. Endophytic Bacillus megaterium BM18-2 mutated for cadmium accumulation and improving plant growth in Hybrid Pennisetum. Biotechnol. Rep. 2019, 24, e00374. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Imtiaz, M.; Nawaz, M.S.; Mubeen, F.; Sarwar, Y.; Hayat, M.; Asif, M.; Naqvi, R.Z.; Ahmad, M.; Imran, A. Thermotolerant PGPR consortium B3P modulates physio-biochemical and molecular machinery for enhanced heat tolerance in maize during early vegetative growth. Ann. Microbiol. 2023, 73, 34. [Google Scholar] [CrossRef]
- Adeleke, B.S.; Ayilara, M.S.; Akinola, S.A.; Babalola, O.O. Biocontrol mechanisms of endophytic fungi. Egypt. J. Biol. Pest Control 2022, 32, 46. [Google Scholar] [CrossRef]
- Tewari, S.; Arora, N. Soybean production under flooding stress and its mitigation using plant growth-promoting microbes. In Environmental Stresses in Soybean Production; Elsevier: Amsterdam, The Netherlands, 2016; pp. 23–40. [Google Scholar]
- Philippot, L.; Chenu, C.; Kappler, A.; Rillig, M.C.; Fierer, N. The interplay between microbial communities and soil properties. Nat. Rev. Microbiol. 2024, 22, 226–239. [Google Scholar] [CrossRef]
- Wang, C.; Kuzyakov, Y. Rhizosphere engineering for soil carbon sequestration. Trends Plant Sci. 2024, 29, 447–468. [Google Scholar] [CrossRef]
- Adeleke, B.S.; Babalola, O.O. The endosphere microbial communities, a great promise in agriculture. Int. Microbiol. 2021, 24, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, A.; Ahmad, E.; Khan, M.S.; Saif, S.; Rizvi, A. Role of plant growth promoting rhizobacteria in sustainable production of vegetables: Current perspective. Sci. Hortic. 2015, 193, 231–239. [Google Scholar] [CrossRef]
- Gupta, G.; Parihar, S.S.; Ahirwar, N.K.; Snehi, S.K.; Singh, V. Plant growth promoting rhizobacteria (PGPR): Current and future prospects for development of sustainable agriculture. J. Microbiol. Biochem. Technol. 2015, 7, 096–102. [Google Scholar]
- Sudheer, S.; Bai, R.G.; Usmani, Z.; Sharma, M. Insights on engineered microbes in sustainable agriculture: Biotechnological developments and future prospects. Curr. Genom. 2020, 21, 321–333. [Google Scholar] [CrossRef]
- O’Callaghan, M.; Ballard, R.A.; Wright, D. Soil microbial inoculants for sustainable agriculture: Limitations and opportunities. Soil Use Manag. 2022, 38, 1340–1369. [Google Scholar] [CrossRef]
- Adeleke, B.S.; Ayangbenro, A.S.; Babalola, O.O. Genomic assessment of Stenotrophomonas indicatrix for improved sunflower plant. Curr. Genet. 2021, 67, 891–907. [Google Scholar] [CrossRef] [PubMed]
- Jansson, J.K.; McClure, R.; Egbert, R.G. Soil microbiome engineering for sustainability in a changing environment. Nat. Biotechnol. 2023, 41, 1716–1728. [Google Scholar] [CrossRef] [PubMed]
- Moreira, Z.P.M.; Chen, M.Y.; Ortuno, D.L.Y.; Haney, C.H. Engineering plant microbiomes by integrating eco-evolutionary principles into current strategies. Curr. Opinion Plant Biol. 2023, 71, 102316. [Google Scholar] [CrossRef] [PubMed]
- Cho, A.; Joshi, A.; Hur, H.-G.; Lee, J.-H. Nodulation experiment by cross-inoculation of nitrogen-fixing bacteria isolated from root nodules of several leguminous plants. J. Microbiol. Biotechnol. 2024, 34, 570. [Google Scholar] [CrossRef] [PubMed]
- Jiao, H.; Wang, R.; Qin, W.; Yang, J. Screening of rhizosphere nitrogen fixing, phosphorus and potassium solubilizing bacteria of Malus sieversii (Ldb.) Roem. and the effect on apple growth. J. Plant Physiol. 2024, 292, 154142. [Google Scholar] [CrossRef]
- Chauhan, P.; Singh, M.; Sharma, A.; Singh, M.; Chadha, P.; Kaur, A. Halotolerant and plant growth-promoting endophytic fungus Aspergillus terreus CR7 alleviates salt stress and exhibits genoprotective effect in Vigna radiata. Front. Microbiol. 2024, 15, 1336533. [Google Scholar] [CrossRef]
- Zeng, R.; Liu, H.; Hong, Z.; Wang, X.; Cheng, S.; Xu, J.; Dai, Z. Co-inoculation effects of B. licheniformis and P. aeruginosa on soil Cd and As availability and rice accumulation. J. Environ. Manag. 2024, 351, 119739. [Google Scholar] [CrossRef]
- Pandino, G.; Abbate, C.; Scavo, A.; Di Benedetto, D.; Mauromicale, G.; Lombardo, S. Co-inoculation of arbuscular mycorrhizal fungi and plant growth promoting bacteria improves plant growth and yield of globe artichoke (Cynara cardunculus scolymus). Sci. Hortic. 2024, 335, 113365. [Google Scholar] [CrossRef]
- Solórzano-Acosta, R.; Toro, M.; Zúñiga-Dávila, D. Effect of co-inoculation with growth-promoting bacteria and arbuscular Mycorrhizae on growth of Persea americana seedlings infected with Phytophthora cinnamomi. Microorganisms 2024, 12, 721. [Google Scholar] [CrossRef]
- Slimani, A.; Oufdou, K.; Meddich, A. Intercropping and co-inoculation of beneficial microorganisms of soils improve drought tolerance in barley and Alfalfa plants. J. Crop Health 2024, 76, 471–485. [Google Scholar] [CrossRef]
- Agunbiade, V.F.; Fadiji, A.E.; Agbodjato, N.A.; Babalola, O.O. Isolation and characterization of plant-growth-promoting, drought-tolerant rhizobacteria for improved maize productivity. Plants 2024, 13, 1298. [Google Scholar] [CrossRef] [PubMed]
- Geller, A.M.; Levy, A. “What I cannot create, I do not understand”: Elucidating microbe–microbe interactions to facilitate plant microbiome engineering. Curr. Opin. Microbiol. 2023, 72, 102283. [Google Scholar] [CrossRef] [PubMed]
- Owen, D.; Williams, A.P.; Griffith, G.W.; Withers, P.J. Use of commercial bio-inoculants to increase agricultural production through improved phosphrous acquisition. Appl. Soil Ecol. 2015, 86, 41–54. [Google Scholar] [CrossRef]
- Xiao, J.-l.; Sun, J.-G.; Pang, B.; Zhou, X.; Gong, Y.; Jiang, L.; Zhang, L.; Ding, X.; Yin, J. Isolation and screening of stress-resistant endophytic fungus strains from wild and cultivated soybeans in cold region of China. Appl. Microbiol. Biotechnol. 2021, 105, 755–768. [Google Scholar] [CrossRef]
- Adeleke, B.; Ayangbenro, A.; Babalola, O. In vitro screening of sunflower associated endophytic bacteria with Plant growth-promoting traits. Front. Sustain. Food Syst. 2022, 6, 903114. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adeleke, B.S.; Chaudhary, P.; Ayilara, M.S.; Ojo, F.M.; Erinoso, S.M.; Upadhayay, V.K.; Adeyemo, A.I.; Akinola, S.A. Rhizosphere Microbiomes Mediating Abiotic Stress Mitigation for Improved Plant Nutrition. Ecologies 2024, 5, 375-401. https://doi.org/10.3390/ecologies5030024
Adeleke BS, Chaudhary P, Ayilara MS, Ojo FM, Erinoso SM, Upadhayay VK, Adeyemo AI, Akinola SA. Rhizosphere Microbiomes Mediating Abiotic Stress Mitigation for Improved Plant Nutrition. Ecologies. 2024; 5(3):375-401. https://doi.org/10.3390/ecologies5030024
Chicago/Turabian StyleAdeleke, Bartholomew Saanu, Parul Chaudhary, Modupe Stella Ayilara, Funmilola Mabel Ojo, Sakiru Morenikeji Erinoso, Viabhav Kumar Upadhayay, Adeyemi Isaiah Adeyemo, and Saheed Adekunle Akinola. 2024. "Rhizosphere Microbiomes Mediating Abiotic Stress Mitigation for Improved Plant Nutrition" Ecologies 5, no. 3: 375-401. https://doi.org/10.3390/ecologies5030024
APA StyleAdeleke, B. S., Chaudhary, P., Ayilara, M. S., Ojo, F. M., Erinoso, S. M., Upadhayay, V. K., Adeyemo, A. I., & Akinola, S. A. (2024). Rhizosphere Microbiomes Mediating Abiotic Stress Mitigation for Improved Plant Nutrition. Ecologies, 5(3), 375-401. https://doi.org/10.3390/ecologies5030024