
Neurite Growth and Electrical Activity in PC-12 Cells: Effects of H3 Receptor-Inspired Electromagnetic Fields and Inherent Schumann Frequencies

{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
1.1. Electromagnetic Fields
1.2. Histamine-H3-Receptor and Wakefulness
1.3. Schumann Resonances and Their Effect on Biological Systems
2. Materials and Methods
2.1. PC-12 Cell Maintenance
2.2. Electromagnetic Field (EMF) Design
2.3. Exposure Protocol
2.4. Data Collection
2.5. Data Analysis
3. Results
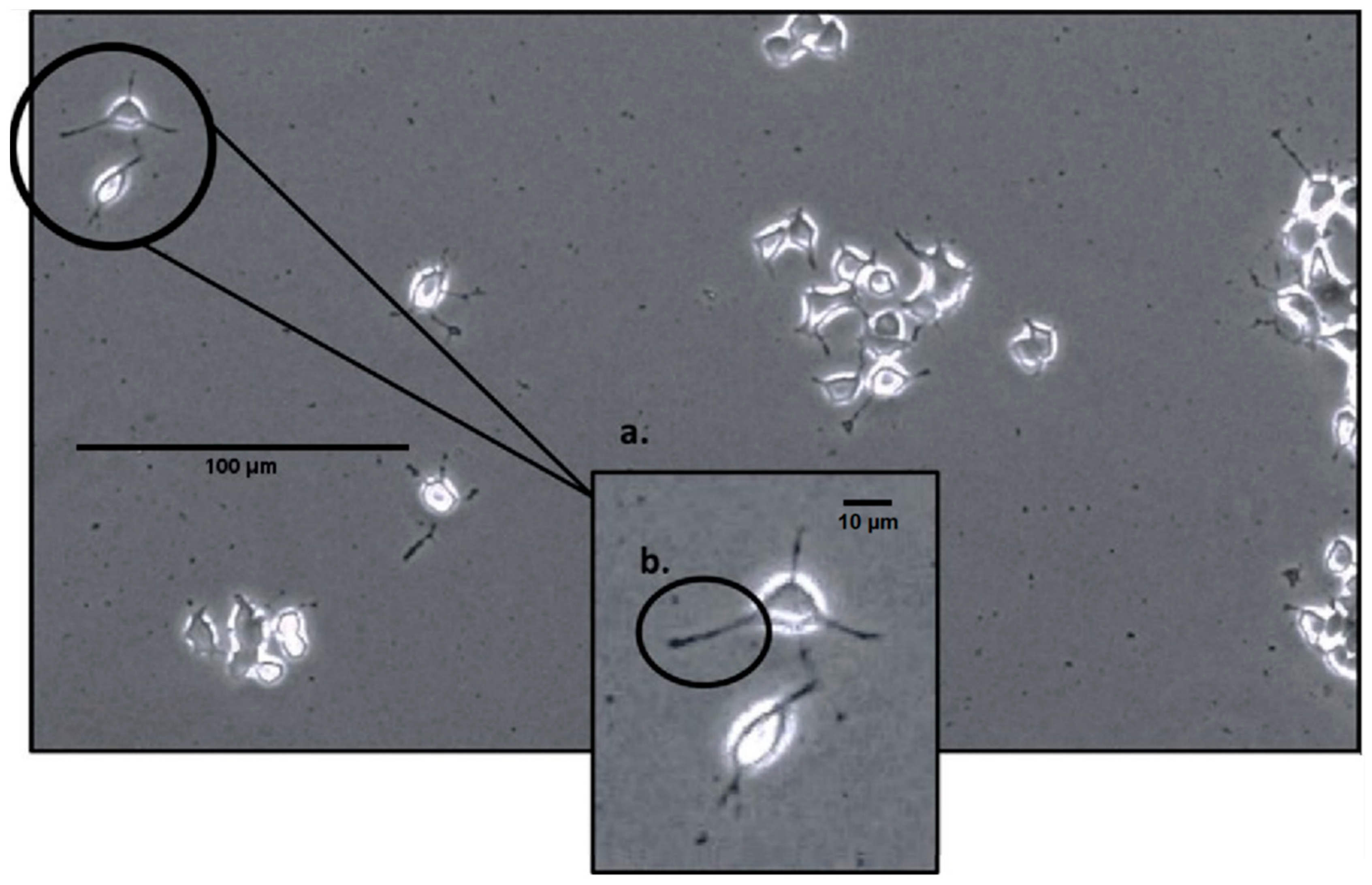
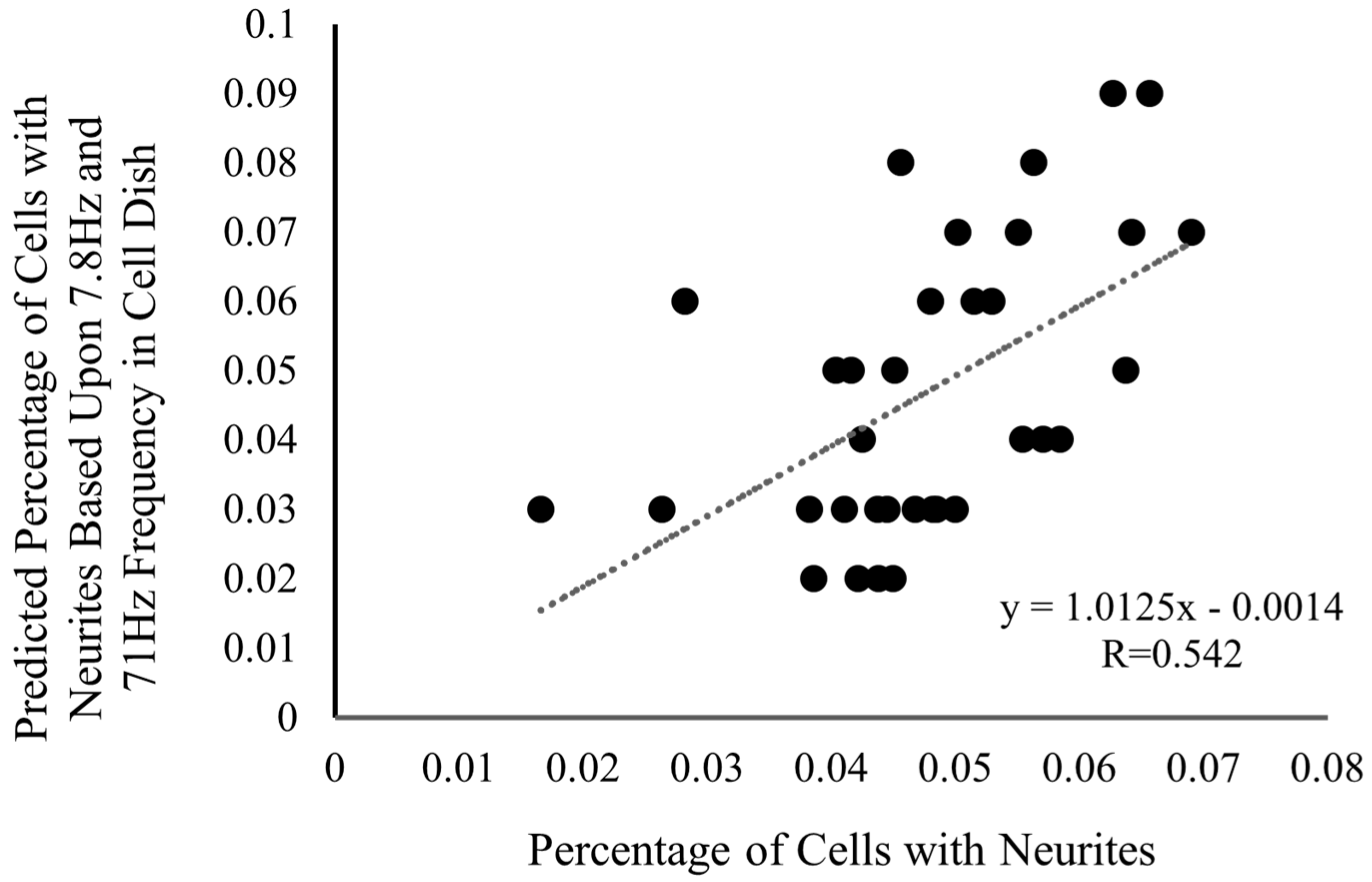
3.1. Neurite Outgrowth and Frequency Analysis
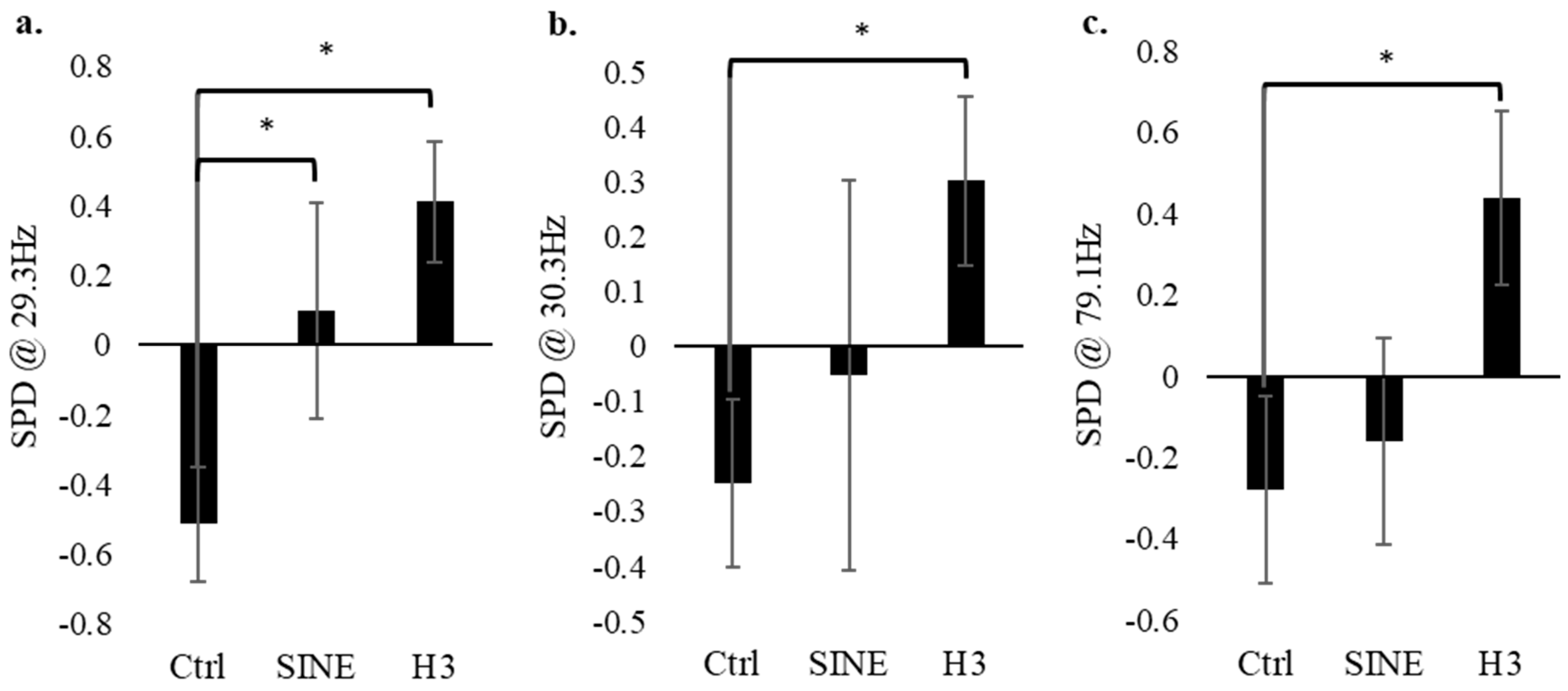
3.2. Electrophysiology of PC-12 Cells–Control vs. SINE vs. H3
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Adey, W.R. Biological effects of electromagnetic fields. J. Cell. Biochem. 1993, 51, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Kovacic, P.; Somanathan, R. Electromagnetic fields: Mechanism, cell signaling, other bioprocesses, toxicity, radicals, antioxidants and beneficial effects. J. Recept. Signal Transduct. 2010, 30, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Li, X.; Bai, L.; Uchida, K.; Bai, W.; Wu, B.; Xu, W.; Zhu, H.; Huang, H. Effects of low frequency electromagnetic field on proliferation of human epidermal stem cells: An in vitro study. Bioelectromagnetics 2013, 34, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Rain, B.; Plourde-Kelly, A.; Lafrenie, R.L.; Dotta, B.T. Induction of apoptosis in B16-BL6 Melanoma Cells Following Exposure to Electromagnetic Fields modelled after Intercellular Calcium Waves. FEBS Open Biol. 2023. [Google Scholar] [CrossRef]
- Ravindran, R.; Branigan, K.S.; Lefebvre, L.M.; Dotta, B.T. Effects of Patterned Electromagnetic Fields and Light-Emitting Diodes on Cancer Cells: Impact on Cell Density and Biophoton Emission When Applied Individually vs. Simultaneously. Appl. Biosci. 2023, 2, 542–549. [Google Scholar] [CrossRef]
- Pesce, M.; Patruno, A.; Speranza, L.; Reale, M. Extremely low frequency electromagnetic field and wound healing: Implication of cytokines as biological mediators. Eur. Cytokine Netw. 2013, 24, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Caliogna, L.; Medetti, M.; Bina, V.; Brancato, A.M.; Castelli, A.; Jannelli, E.; Ivone, A.; Gastaldi, G.; Annunziata, S.; Mosconi, M.; et al. Pulsed electromagnetic fields in bone healing: Molecular pathways and clinical applications. Int. J. Mol. Sci. 2021, 22, 7403. [Google Scholar] [CrossRef] [PubMed]
- Richter-Landsberg, C.; Jastorff, B. The role of cAMP in nerve growth factor-promoted neurite outgrowth in PC-12 cells. J. Cell Biol. 1986, 102, 821–829. [Google Scholar] [CrossRef]
- Carniello, T.N.; Lafrenie, R.M.; Dotta, B.T. The Influence of Burst-Firing EMF on Forskolin-Induced Pheochromocytoma (PC-12) Plasma Membrane Extensions. NeuroSci 2021, 2, 383–399. [Google Scholar] [CrossRef]
- Lachapelle, H.M. Neuroprotective Effects of Fatty Acid Amide Hydrolase Inhibitor (URB597) and Low-Frequency EMF against Paraquat-Induced Neurotoxicity: Relevance to Neurodegenerative Disease. Ph.D. Thesis, Laurentian University of Sudbury, Sudbury, ON, Canada, 2022. [Google Scholar]
- Plourde-Kelly, A. Protein-Patterned EMF as Enzymatic Moderators: Evidence for a Non-Invasive Method of Cellular Manipulation. Ph.D. Thesis, Laurentian University of Sudbury, Sudbury, ON, Canada, 2022. [Google Scholar]
- Naeser, M.A.; Martin, P.I.; Treglia, E.; Ho, M.; Kaplan, E.; Bashir, S.; Hamilton, R.; Coslett, H.B.; Pascual-Leone, A. Research with rTMS in the treatment of aphasia. Restor. Neurol. Neurosci. 2010, 28, 511–529. [Google Scholar] [CrossRef]
- Wang, H.N.; Wang, L.; Zhang, R.G.; Chen, Y.C.; Liu, L.; Gao, F.; Nie, H.; Hou, W.-G.; Peng, Z.-W.; Tan, Q. Anti-depressive mechanism of repetitive transcranial magnetic stimulation in rat: The role of the endocannabinoid system. J. Psychiatr. Res. 2014, 51, 79–87. [Google Scholar] [CrossRef]
- Rutherford, G.; Lithgow, B.; Moussavi, Z. Short and long-term effects of rTMS treatment on Alzheimer’s disease at different stages: A pilot study. J. Exp. Neurosci. 2015, 9, JEN-S24004. [Google Scholar] [CrossRef]
- Lefaucheur, J.P.; André-Obadia, N.; Antal, A.; Ayache, S.S.; Baeken, C.; Benninger, D.H.; Cantello, R.M.; Cincotta, M.; de Carvalho, M.; De Ridder, D.; et al. Evidence-based guidelines on the therapeutic use of repetitive transcranial magnetic stimulation (rTMS). Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2014, 125, 2150–2206. [Google Scholar] [CrossRef]
- Hoogendam, J.M.; Ramakers, G.M.; Di Lazzaro, V. Physiology of repetitive transcranial magnetic stimulation of the human brain. Brain Stimul. 2010, 3, 95–118. [Google Scholar] [CrossRef] [PubMed]
- Priori, A.; Hallett, M.; Rothwell, J.C. Repetitive transcranial magnetic stimulation or transcranial direct current stimulation? Brain Stimul. 2009, 2, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, V.; Thakkar, M.M.; Strecker, R.E.; Basheer, R.; McCarley, R.W. Wakefulness-inducing effects of histamine in the basal forebrain of freely moving rats. Behav. Brain Res. 2004, 152, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Chu, M.; Huang, Z.L.; Qu, W.M.; Eguchi, N.; Yao, M.H.; Urade, Y. Extracellular histamine level in the frontal cortex is positively correlated with the amount of wakefulness in rats. Neurosci. Res. 2004, 49, 417–420. [Google Scholar] [CrossRef] [PubMed]
- Thakkar, M.M. Histamine in the regulation of wakefulness. Sleep Med. Rev. 2011, 15, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Bonaventure, P.; Letavic, M.; Dugovic, C.; Wilson, S.; Aluisio, L.; Pudiak, C.; Lord, B.; Mazur, C.; Kamme, F.; Nishino, S.; et al. Histamine H3 receptor antagonists: From target identification to drug leads. Pharmacology 2007, 73, 1084–1096. [Google Scholar] [CrossRef]
- Huang, Z.L.; Mochizuki, T.; Qu, W.M.; Hong, Z.Y.; Watanabe, T.; Urade, Y.; Hayaishi, O. Altered sleep–wake characteristics and lack of arousal response to H3 receptor antagonist in histamine H1 receptor knockout mice. Proc. Natl. Acad. Sci. USA 2006, 103, 4687–4692. [Google Scholar] [CrossRef]
- Leurs, R.; Blandina, P.; Tedford, C.; Timmerman, H. Therapeutic potential of histamine H3 receptor agonists and antagonists. Trends Pharmacol. Sci. 1998, 19, 177–184. [Google Scholar] [CrossRef]
- Parmentier, R.; Anaclet, C.; Guhennec, C.; Brousseau, E.; Bricout, D.; Giboulot, T.; Bozyczko-Coyne, D.; Spiegel, K.; Ohtsu, H.; Williams, M.; et al. The brain H3-receptor as a novel therapeutic target for vigilance and sleep–wake disorders. Biochem. Pharmacol. 2007, 73, 1157–1171. [Google Scholar] [CrossRef]
- Lin, J.S.; Dauvilliers, Y.; Arnulf, I.; Bastuji, H.; Anaclet, C.; Parmentier, R.; Kocher, L.; Yanagisawa, M.; Lehert, P.; Ligneau, X.; et al. An inverse agonist of the histamine H3 receptor improves wakefulness in narcolepsy: Studies in orexin−/− mice and patients. Neurobiol. Dis. 2008, 30, 74–83. [Google Scholar] [CrossRef]
- Barbier, A.J.; Berridge, C.; Dugovic, C.; Laposky, A.D.; Wilson, S.J.; Boggs, J.; Aluisio, L.; Lord, B.; Mazur, C.; Pudiak, C.M.; et al. Acute wake-promoting actions of JNJ-5207852, a novel, diamine-based H3 antagonist. Br. J. Pharmacol. 2004, 143, 649–661. [Google Scholar] [CrossRef]
- Passani, M.B.; Lin, J.S.; Hancock, A.; Crochet, S.; Blandina, P. The histamine H3 receptor as a novel therapeutic target for cognitive and sleep disorders. Trends Pharmacol. Sci. 2004, 25, 618–625. [Google Scholar] [CrossRef] [PubMed]
- Mach, Q.H.; Persinger, M.A. Behavioral changes with brief exposures to weak magnetic fields patterned to stimulate long-term potentiation. Brain Res. 2009, 1261, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Hu, W.-W.; Fan, Y.-Y.; Dai, H.-B.; Fu, Q.-L.; Wei, E.-Q.; Luo, J.-H.; Chen, Z. Carnosine protects against NMDA-induced neurotoxicity in differentiated rat PC-12 cells through carnosine-histidine-histamine pathway and H1/H3 receptors. Biochem. Pharmacol. 2007, 73, 709–717. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.; Dai, H.; He, P.; Hu, W.; Fan, Y.; Zhang, W.; Chen, Z. The H3 receptor antagonist clobenpropit protects against Abeta42-induced neurotoxicity in differentiated rat PC-12 cells. Die Pharm. 2010, 65, 257–260. [Google Scholar]
- Hashikawa-Hobara, N.; Chan, N.Y.K.; Levi, R. Histamine 3 receptor activation reduces the expression of neuronal angiotensin II type 1 receptors in the heart. J. Pharmacol. Exp. Ther. 2012, 340, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Price, C. ELF electromagnetic waves from lightning: The Schumann resonances. Atmosphere 2016, 7, 116. [Google Scholar] [CrossRef]
- Danho, S.; Schoellhorn, W.; Aclan, M. Innovative technical implementation of the Schumann resonances and its influence on organisms and biological cells. IOP Conf. Ser. Mater. Sci. Eng. 2019, 564, 012081. [Google Scholar] [CrossRef]
- Elhalel, G.; Price, C.; Fixler, D.; Shainberg, A. Cardioprotection from stress conditions by weak magnetic fields in the Schumann Resonance band. Sci. Rep. 2019, 9, 1645. [Google Scholar] [CrossRef] [PubMed]
- Martel, J.; Chang, S.H.; Chevalier, G.; Ojcius, D.M.; Young, J.D. Influence of electromagnetic fields on the circadian rhythm: Implications for human health and disease. Biomed. J. 2023, 46, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Fdez-Arroyabe, P.; Fornieles-Callejon, J.; Santurtun, A.; Szangolies, L.; Donner, R.V. Schumann resonance and cardiovascular hospital admission in the area of Granada, Spain: An event coincidence analysis approach. Sci. Total Environ. 2020, 705, 135813. [Google Scholar] [CrossRef] [PubMed]
- Cherry, N.J. Human intelligence: The brain, an electromagnetic system synchronised by the Schumann resonance signal. Med. Hypotheses 2003, 60, 843–844. [Google Scholar] [CrossRef] [PubMed]
- Bongers, G.; Bakker, R.A.; Leurs, R. Molecular aspects of the histamine H3 receptor. Biochem. Pharmacol. 2007, 73, 1195–1204. [Google Scholar] [CrossRef] [PubMed]
- Bhowmik, M.; Khanam, R.; Vohora, D. Histamine H3 receptor antagonists in relation to epilepsy and neurodegeneration: A systemic consideration of recent progress and perspectives. Br. J. Pharmacol. 2012, 167, 1398–1414. [Google Scholar] [CrossRef]
- Chen, J.; Liu, C.; Lovenberg, T.W. Molecular and pharmacological characterization of the mouse histamine H3 receptor. Eur. J. Pharmacol. 2003, 467, 57–65. [Google Scholar] [CrossRef]
- Cosic, I. Macromolecular bioactivity: Is it resonant interaction between macromolecules?-theory and applications. IEEE Trans. Biomed. Eng. 1994, 41, 1101–1114. [Google Scholar] [CrossRef]
- Ross, M.L.; Koren, S.A.; Persinger, M.A. Physiologically patterned weak magnetic fields applied over left frontal lobe increase acceptance of false statements as true. Electromagn. Biol. Med. 2008, 27, 365–371. [Google Scholar] [CrossRef]
- Zhuang, Y.; Ge, Q.; Li, Q.; Xu, L.; Geng, X.; Wang, R.; He, J. Combined behavioral and EEG evidence for the 70 Hz frequency selection of short-term spinal cord stimulation in disorders of consciousness. CNS Neurosci. Ther. 2023. [Google Scholar] [CrossRef] [PubMed]
- Kanno, T.; Morita, I.; Yamaguchi, S.; Yokoyama, T.; Kamei, Y.; Anil, S.M.; Karagiozov, K.L. Dorsal column stimulation in persistent vegetative state. Neuromodul. Technol. Neural Interface 2009, 12, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Mulligan, B.P. Biological Effects of Actual and Simulated Geomagnetic Fields. Master’s Thesis, Library and Archives Canada = Bibliothèque et Archives Canada, Ottawa, ON, Canada, 2012. [Google Scholar]
- Rouleau, N.; Dotta, B.T. Electromagnetic fields as structure-function zeitgebers in biological systems: Environmental orchestrations of morphogenesis and consciousness. Front. Integr. Neurosci. 2014, 8, 84. [Google Scholar] [CrossRef] [PubMed]
- Plourde-Kelly, A.D.; Dotta, B.; Koren, S. p38 MAPK patterned EMF affects PC-12 neurite outgrowth after 2 days of treatment. Brain Stimul. 2021, 14, 1694. [Google Scholar] [CrossRef]
- Lin, J.S.; Sergeeva, O.A.; Haas, H.L. Histamine H3 receptors and sleep-wake regulation. J. Pharmacol. Exp. Ther. 2011, 336, 17–23. [Google Scholar] [CrossRef]
- Atzori, M.; Lau, D.; Tansey, E.P.; Chow, A.; Ozaita, A.; Rudy, B.; McBain, C.J. H2 histamine receptor-phosphorylation of Kv3. 2 modulates interneuron fast spiking. Nat. Neurosci. 2000, 3, 791–798. [Google Scholar] [CrossRef]
- Giordano, G.; Costa, L.G. Primary neurons in culture and neuronal cell lines for in vitro neurotoxicological studies. In In Vitro Neurotoxicology: Methods and Protocols; Humana Press: Totowa, NJ, USA, 2011; pp. 13–27. [Google Scholar]
- Oprea, D.; Sanz, C.G.; Barsan, M.M.; Enache, T.A. PC-12 cell line as a neuronal cell model for biosensing applications. Biosensors 2022, 12, 500. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lefebvre, L.M.; Plourde-Kelly, A.D.; Saroka, K.S.; Dotta, B.T. Neurite Growth and Electrical Activity in PC-12 Cells: Effects of H3 Receptor-Inspired Electromagnetic Fields and Inherent Schumann Frequencies. Biophysica 2024, 4, 74-82. https://doi.org/10.3390/biophysica4010005
Lefebvre LM, Plourde-Kelly AD, Saroka KS, Dotta BT. Neurite Growth and Electrical Activity in PC-12 Cells: Effects of H3 Receptor-Inspired Electromagnetic Fields and Inherent Schumann Frequencies. Biophysica. 2024; 4(1):74-82. https://doi.org/10.3390/biophysica4010005
Chicago/Turabian StyleLefebvre, Landon M., Adam D. Plourde-Kelly, Kevin S. Saroka, and Blake T. Dotta. 2024. "Neurite Growth and Electrical Activity in PC-12 Cells: Effects of H3 Receptor-Inspired Electromagnetic Fields and Inherent Schumann Frequencies" Biophysica 4, no. 1: 74-82. https://doi.org/10.3390/biophysica4010005
APA StyleLefebvre, L. M., Plourde-Kelly, A. D., Saroka, K. S., & Dotta, B. T. (2024). Neurite Growth and Electrical Activity in PC-12 Cells: Effects of H3 Receptor-Inspired Electromagnetic Fields and Inherent Schumann Frequencies. Biophysica, 4(1), 74-82. https://doi.org/10.3390/biophysica4010005