
Investigating the Insertion Mechanism of Cell-Penetrating Peptide Penetratin into Cell Membranes: Implications for Targeted Drug Delivery



Abstract
:1. Introduction
2. Results
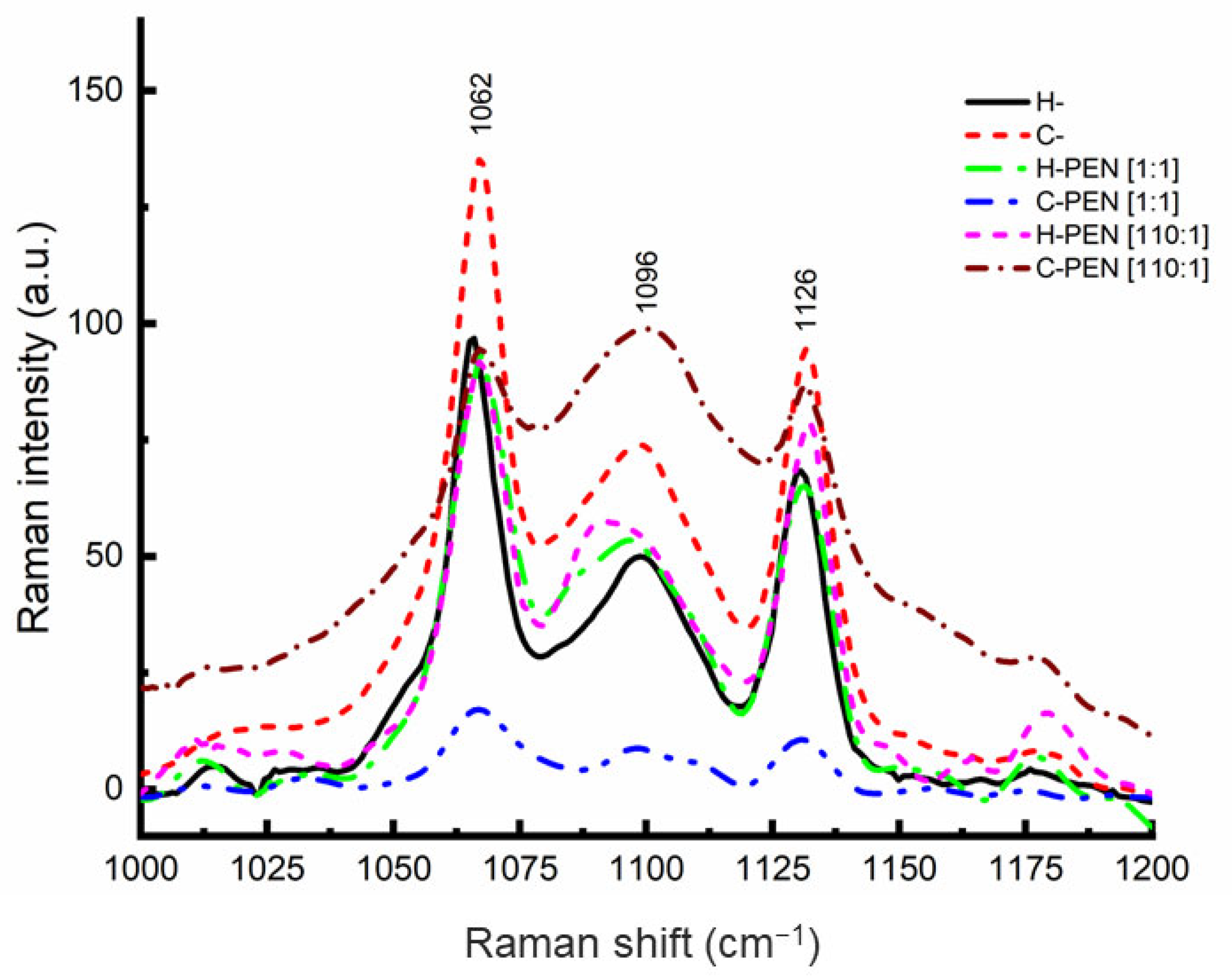
2.1. Characterization of PEN–Vesicle Complexes by Raman Spectroscopy
2.2. Characterization of PEN–Vesicle Complexes by DLS and Zeta Potential
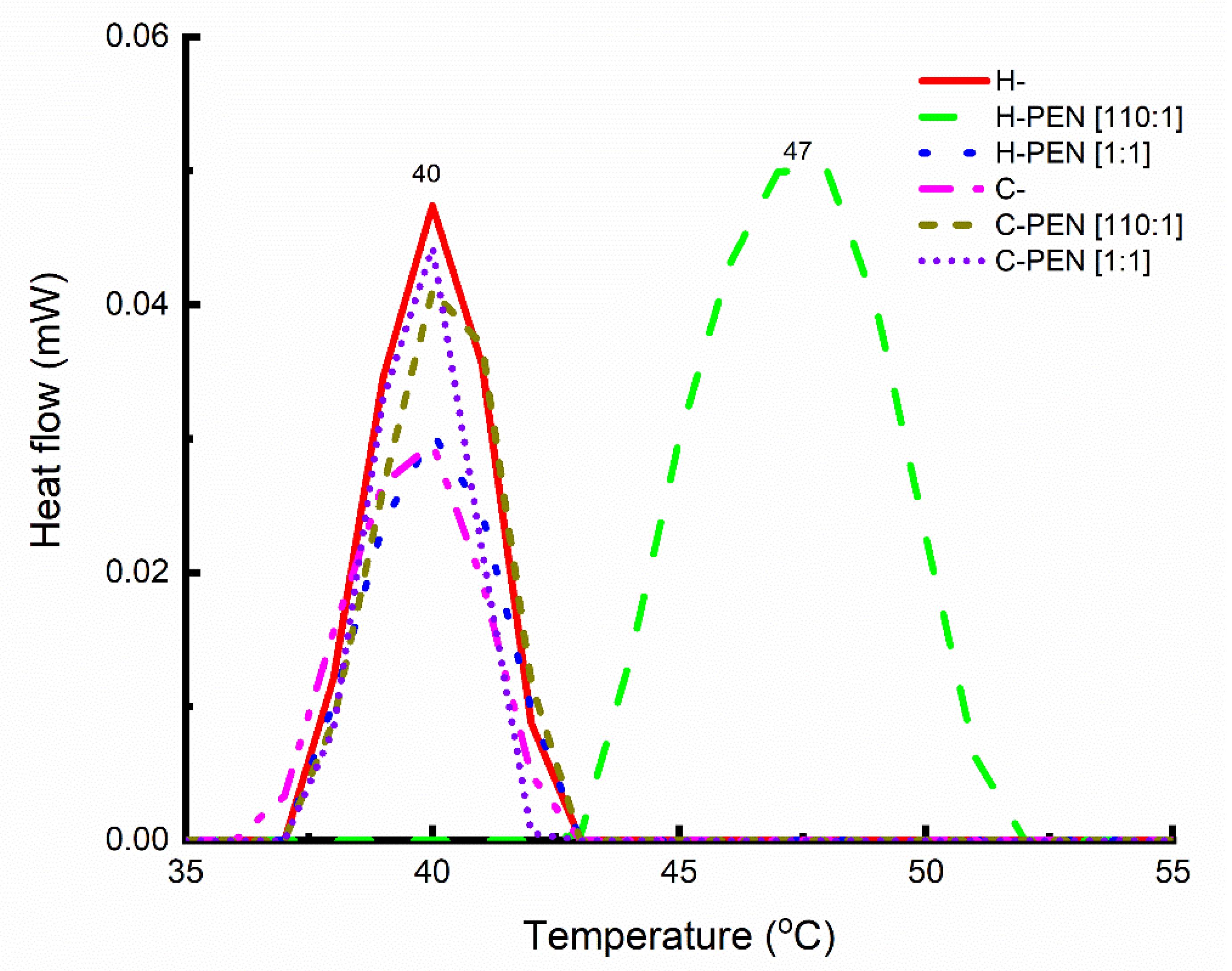
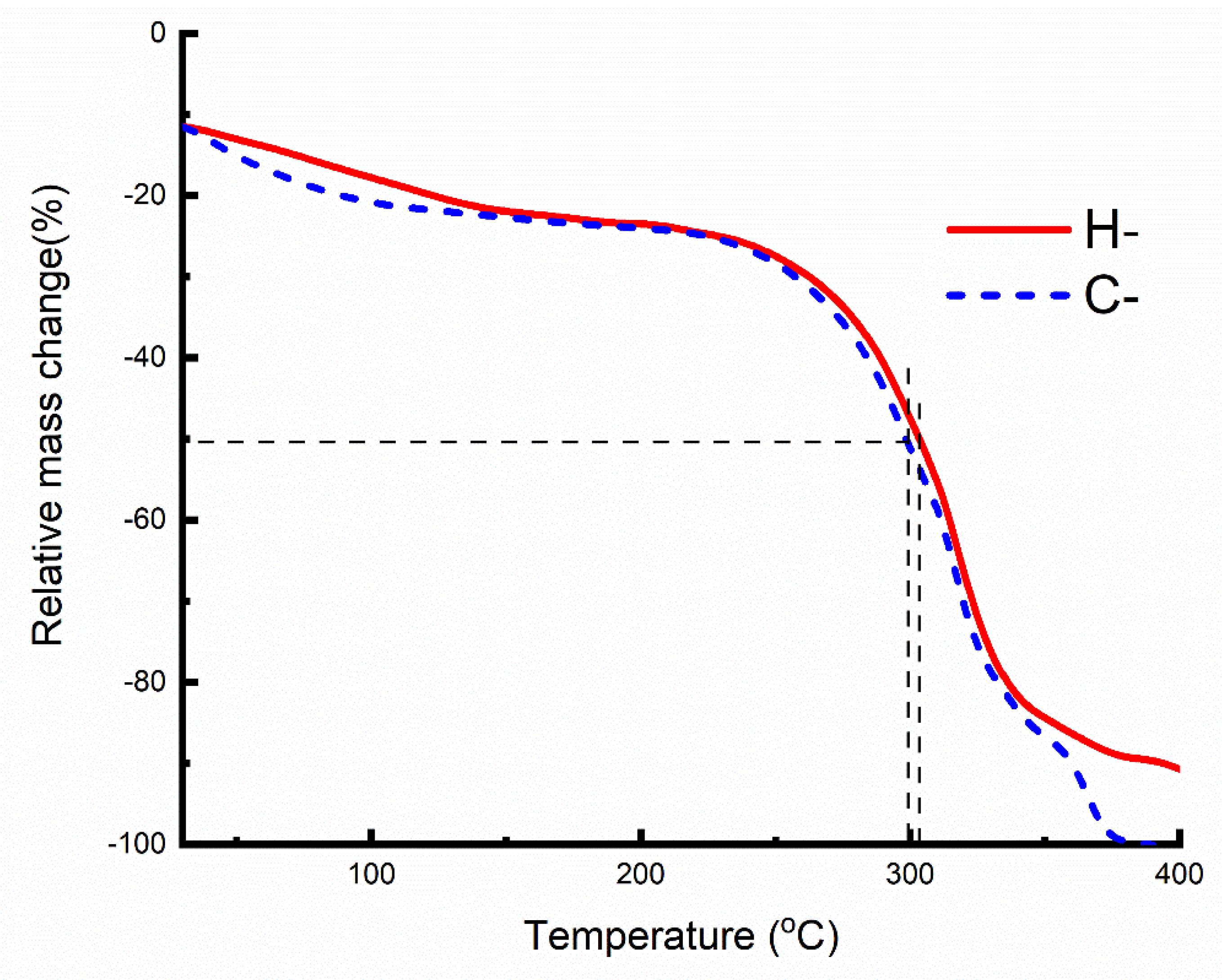
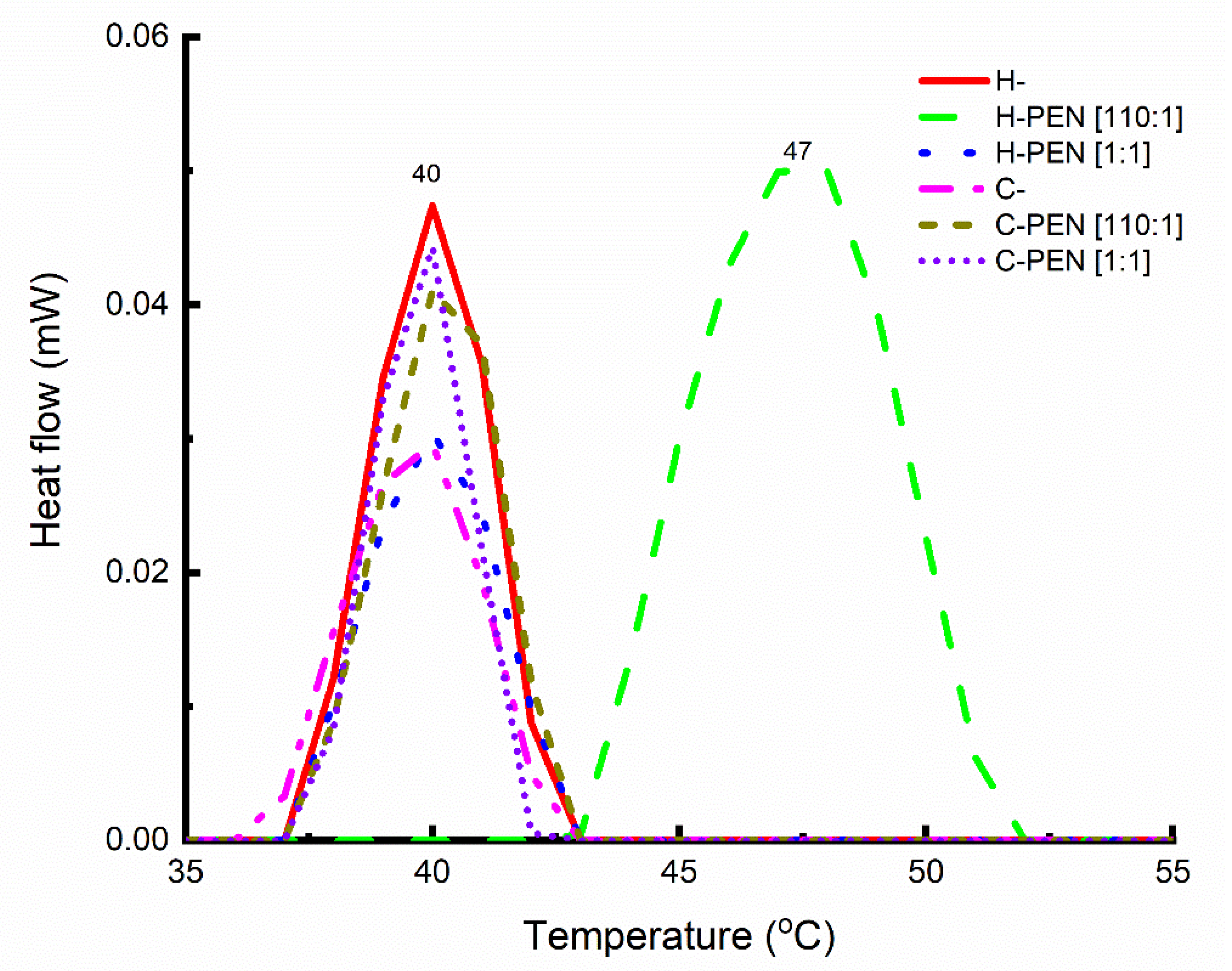
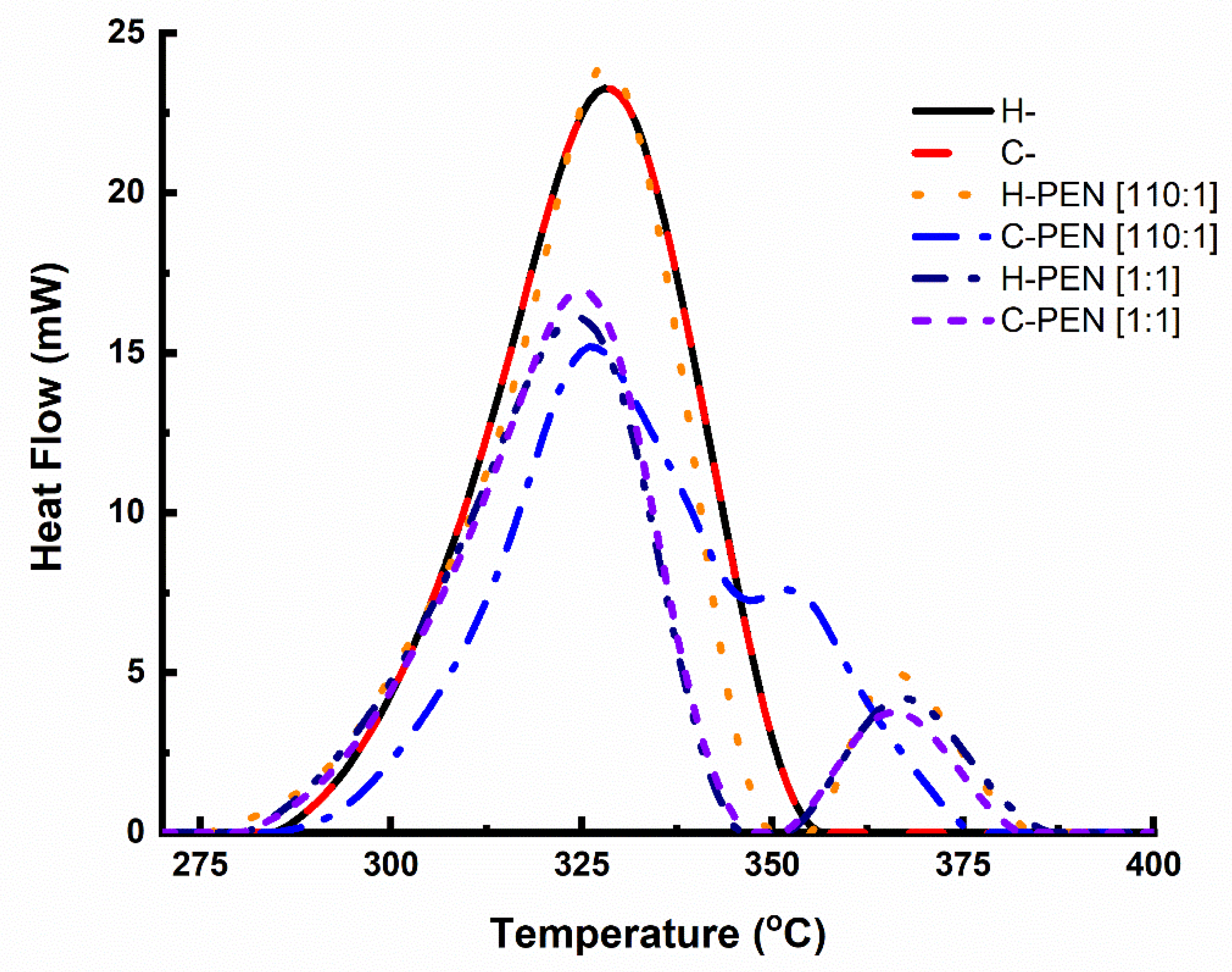
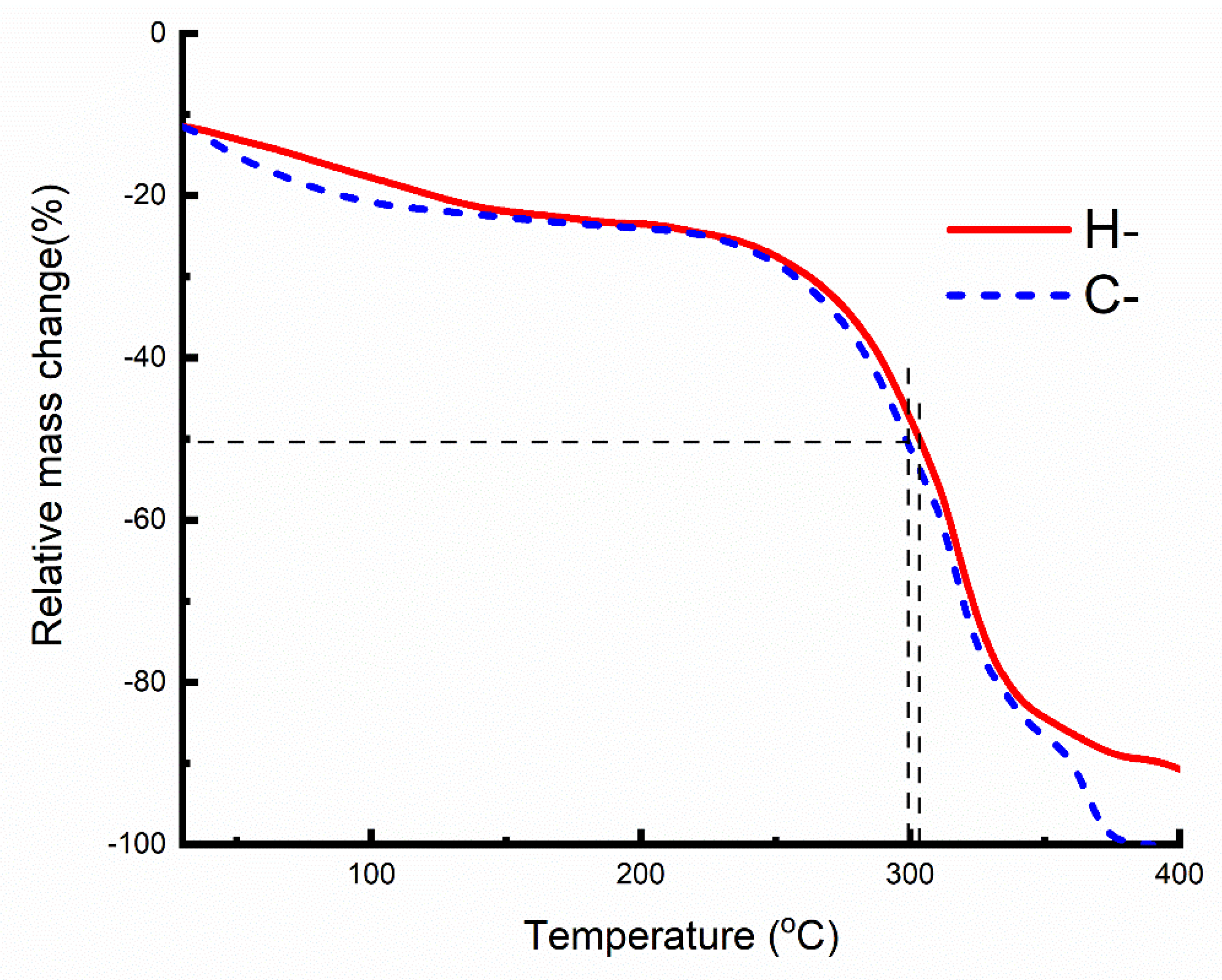
2.3. Thermal Analysis
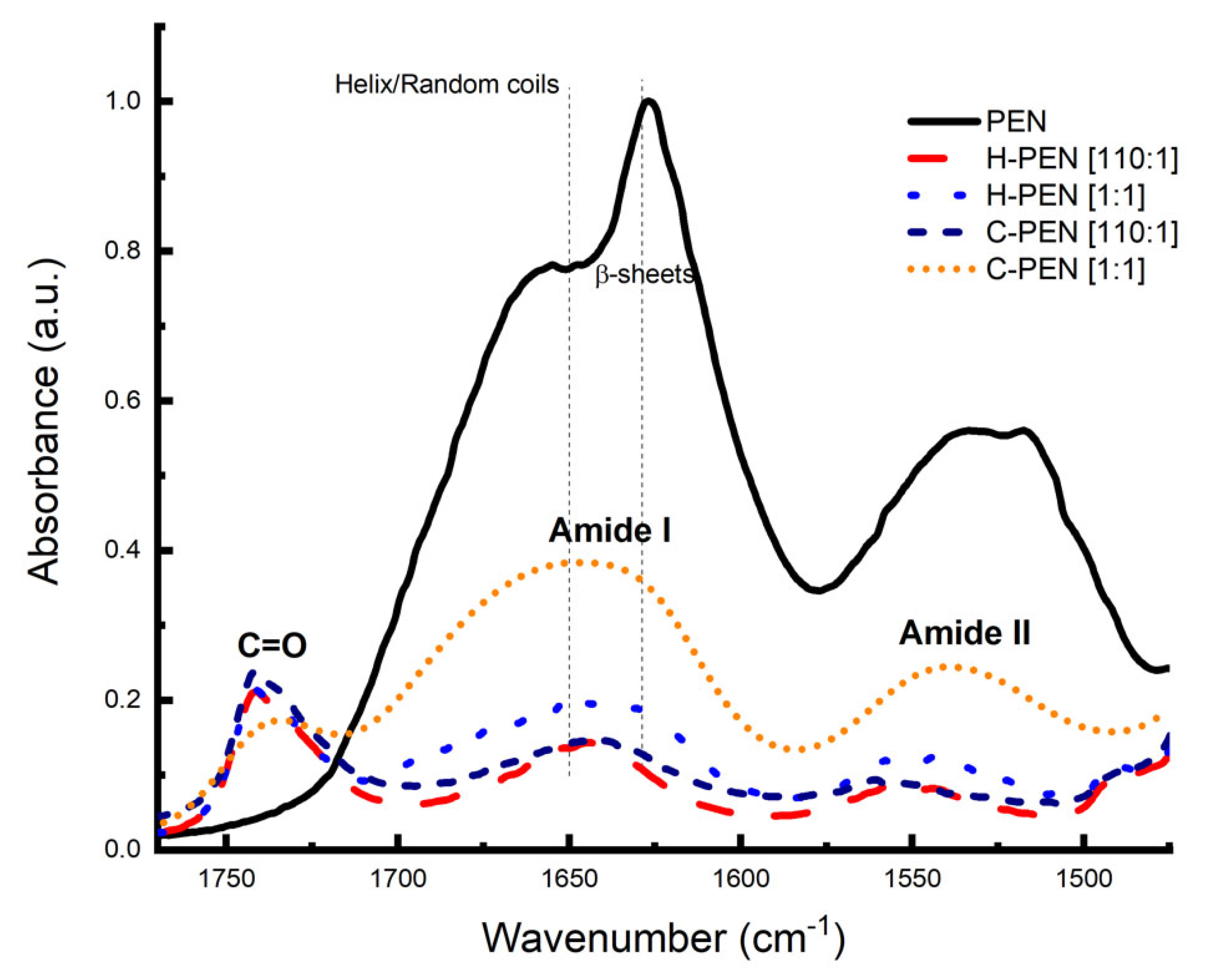
2.4. Structural Bond Analysis
3. Discussion
3.1. Finding A Specific Lipid-to-Peptide Molar Ratio Triggering Direct Insertion of PEN into Cells
3.2. Mechanism of Interaction between C- Model and PEN at the Lipid-to Peptide Molar Ratio of 110
3.3. Mechanism of Interaction between H- and PEN at the Lipid-to Peptide Molar Ratio of 110
4. Materials and Methods
4.1. Materials
4.2. Preparation of Non-Cancer and Tumor Cell Membrane Mimetic Vesicles
4.3. Particle Size and Zeta Potential Measurements
4.4. Raman Spectroscopy
4.5. Infrared Spectroscopy
4.6. Thermogravimetric Analysis (TGA) and Differential Scanning Calorimetry (DSC)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Habault, J.; Poyet, J.-L. Recent Advances in Cell Penetrating Peptide-Based Anticancer Therapies. Molecules 2019, 24, 927. [Google Scholar] [CrossRef] [PubMed]
- Regberg, J.; Srimanee, A.; Langel, Ü. Applications of Cell-Penetrating Peptides for Tumor Targeting and Future Cancer Therapies. Pharmaceuticals 2012, 5, 991–1007. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.T.; Sayers, E.J. Cell entry of cell penetrating peptides: Tales of tails wagging dogs. J. Control. Release 2012, 161, 582–591. [Google Scholar] [CrossRef] [PubMed]
- González-Cruz, A.O.; Hernández-Juárez, J.; Ramírez-Cabrera, M.A.; Balderas-Rentería, I.; Arredondo-Espinoza, E. Peptide-based drug-delivery systems: A new hope for improving cancer therapy. J. Drug Deliv. Sci. Technol. 2022, 72, 103362. [Google Scholar] [CrossRef]
- Stiltner, J.; McCandless, K.; Zahid, M. Cell-Penetrating Peptides: Applications in Tumor Diagnosis and Therapeutics. Pharmaceutics 2021, 13, 890. [Google Scholar] [CrossRef]
- Liu, C.; Tai, L.; Zhang, W.; Wei, G.; Pan, W.; Lu, W. Penetratin, a Potentially Powerful Absorption Enhancer for Noninvasive Intraocular Drug Delivery. Mol. Pharm. 2014, 11, 1218–1227. [Google Scholar] [CrossRef]
- Jiao, C.-Y.; Delaroche, D.; Burlina, F.; Alves, I.D.; Chassaing, G.; Sagan, S. Translocation and Endocytosis for Cell-penetrating Peptide Internalization. J. Biol. Chem. 2009, 284, 33957–33965. [Google Scholar] [CrossRef]
- Zamotaiev, O.M.; Postupalenko, V.Y.; Shvadchak, V.V.; Pivovarenko, V.G.; Klymchenko, A.S.; Mély, Y. Monitoring penetratin interactions with lipid membranes and cell internalization using a new hydration-sensitive fluorescent probe. Org. Biomol. Chem. 2014, 12, 7036–7044. [Google Scholar] [CrossRef]
- Thorén, P.E.G.; Persson, D.; Isakson, P.; Goksör, M.; Önfelt, A.; Nordén, B. Uptake of analogs of penetratin, Tat(48–60) and oligoarginine in live cells. Biochem. Biophys. Res. Commun. 2003, 307, 100–107. [Google Scholar] [CrossRef]
- Miwa, A.; Kamiya, K. Control of Enzyme Reaction Initiation inside Giant Unilamellar Vesicles by the Cell-Penetrating Peptide-Mediated Translocation of Cargo Proteins. ACS Synth. Biol. 2022, 11, 3836–3846. [Google Scholar] [CrossRef]
- Nigatu, A.S.; Vupputuri, S.; Flynn, N.; Ramsey, J.D. Effects of cell-penetrating peptides on transduction efficiency of PEGylated adenovirus. Biomed. Pharmacother. 2015, 71, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Yin, T.; Xie, W.; Sun, J.; Yang, L.; Liu, J. Penetratin Peptide-Functionalized Gold Nanostars: Enhanced BBB Permeability and NIR Photothermal Treatment of Alzheimer’s Disease Using Ultralow Irradiance. ACS Appl. Mater. Interfaces 2016, 8, 19291–19302. [Google Scholar] [CrossRef]
- Wang, W.; Liu, W.; Xu, S.; Dong, X.; Sun, Y. Design of Multifunctional Agent Based on Basified Serum Albumin for Efficient In Vivo β-Amyloid Inhibition and Imaging. ACS Appl. Bio Mater. 2020, 3, 3365–3377. [Google Scholar] [CrossRef] [PubMed]
- Gehan, P.; Kulifaj, S.; Soule, P.; Bodin, J.B.; Amoura, M.; Walrant, A.; Sagan, S.; Thiam, A.R.; Ngo, K.; Vivier, V.; et al. Penetratin translocation mechanism through asymmetric droplet interface bilayers. Biochim. Biophys. Acta BBA Biomembr. 2020, 1862, 183415. [Google Scholar] [CrossRef] [PubMed]
- Binder, H.; Lindblom, G. A Molecular View on the Interaction of the Trojan Peptide Penetratin with the Polar Interface of Lipid Bilayers. Biophys. J. 2004, 87, 332–343. [Google Scholar] [CrossRef]
- Joanne, P.; Galanth, C.; Goasdoué, N.; Nicolas, P.; Sagan, S.; Lavielle, S.; Chassaing, G.; El Amri, C.; Alves, I.D. Lipid reorganization induced by membrane-active peptides probed using differential scanning calorimetry. Biochim. Biophys. Acta BBA Biomembr. 2009, 1788, 1772–1781. [Google Scholar] [CrossRef]
- Peetla, C.; Stine, A.; Labhasetwar, V. Biophysical interactions with model lipid membranes: Applications in drug discovery and drug delivery. Mol. Pharm. 2009, 6, 1264–1276. [Google Scholar] [CrossRef]
- Kay, J.G.; Grinstein, S. Sensing Phosphatidylserine in Cellular Membranes. Sensors 2011, 11, 1744–1755. [Google Scholar] [CrossRef]
- Riedl, S.; Rinner, B.; Asslaber, M.; Schaider, H.; Walzer, S.; Novak, A.; Lohner, K.; Zweytick, D. In search of a novel target—Phosphatidylserine exposed by non-apoptotic tumor cells and metastases of malignancies with poor treatment efficacy. Biochim. Biophys. Acta BBA Biomembr. 2011, 1808, 2638–2645. [Google Scholar] [CrossRef]
- Alves, I.D.; Jiao, C.-Y.; Aubry, S.; Aussedat, B.; Burlina, F.; Chassaing, G.; Sagan, S. Cell biology meets biophysics to unveil the different mechanisms of penetratin internalization in cells. Biochim. Biophys. Acta 2010, 1798, 2231–2239. [Google Scholar] [CrossRef]
- Carrier, D.; Pézolet, M. Raman spectroscopic study of the interaction of poly-L-lysine with dipalmitoylphosphatidylglycerol bilayers. Biophys. J. 1984, 46, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Wodlej, C.; Riedl, S.; Rinner, B.; Leber, R.; Drechsler, C.; Voelker, D.R.; Choi, J.-Y.; Lohner, K.; Zweytick, D. Interaction of two antitumor peptides with membrane lipids—Influence of phosphatidylserine and cholesterol on specificity for melanoma cells. PLoS ONE 2019, 14, e0211187. [Google Scholar] [CrossRef] [PubMed]
- Dimova, R.; Aranda, S.; Bezlyepkina, N.; Nikolov, V.; Riske, K.A.; Lipowsky, R. A practical guide to giant vesicles. Probing the membrane nanoregime via optical microscopy. J. Phys. Condens. Matter. 2006, 18, S1151–S1176. [Google Scholar] [CrossRef]
- Liu, Q.; Bi, C.; Li, J.; Liu, X.; Peng, R.; Jin, C.; Sun, Y.; Lyu, Y.; Liu, H.; Wang, H.; et al. Generating Giant Membrane Vesicles from Live Cells with Preserved Cellular Properties. Research 2019, 2019, 6523970. [Google Scholar] [CrossRef]
- Persson, D.; Thorén, P.E.G.; Esbjörner, E.K.; Goksör, M.; Lincoln, P.; Nordén, B. Vesicle size-dependent translocation of penetratin analogs across lipid membranes. Biochim. Biophys. Acta BBA Biomembr. 2004, 1665, 142–155. [Google Scholar] [CrossRef] [PubMed]
- Kitt, J.P.; Bryce, D.A.; Minteer, S.D.; Harris, J.M. Raman Spectroscopy Reveals Selective Interactions of Cytochrome c with Cardiolipin That Correlate with Membrane Permeability. J. Am. Chem. Soc. 2017, 139, 3851–3860. [Google Scholar] [CrossRef] [PubMed]
- Bryce, D.A.; Kitt, J.P.; Harris, J.M. Raman Microscopy Investigation of GLP-1 Peptide Association with Supported Phospholipid Bilayers. Langmuir 2021, 37, 14265–14274. [Google Scholar] [CrossRef]
- Czamara, K.; Majzner, K.; Pacia, M.Z.; Kochan, K.; Kaczor, A.A.; Baranska, M. Raman spectroscopy of lipids: A review. J. Raman Spectrosc. 2015, 46, 4–20. [Google Scholar] [CrossRef]
- Pu, C.; Tang, W. Affinity and selectivity of anchovy antibacterial peptide for Staphylococcus aureus cell membrane lipid and its application in whole milk. Food Control. 2017, 72, 153–163. [Google Scholar] [CrossRef]
- Díaz-Caballero, M.; Navarro, S.; Ventura, S. Soluble Assemblies in the Fibrillation Pathway of Prion-Inspired Artificial Functional Amyloids are Highly Cytotoxic. Biomacromolecules 2020, 21, 2334–2345. [Google Scholar] [CrossRef]
- Krafft, C.; Neudert, L.; Simat, T.; Salzer, R. Near infrared Raman spectra of human brain lipids. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2005, 61, 1529–1535. [Google Scholar] [CrossRef]
- Maherani, B.; Arab-Tehrany, E.; Rogalska, E.; Korchowiec, B.; Kheirolomoom, A.; Linder, M. Vibrational, calorimetric, and molecular conformational study on calcein interaction with model lipid membrane. J. Nanoparticle Res. 2013, 15, 1792. [Google Scholar] [CrossRef]
- Domingues, M.M.; Santiago, P.S.; Castanho, M.A.R.B.; Santos, N.C. What can light scattering spectroscopy do for membrane-active peptide studies? J. Pept. Sci. Off. Publ. Eur. Pept. Soc. 2008, 14, 394–400. [Google Scholar] [CrossRef] [PubMed]
- Dandurand, J.; Samouillan, V.; Lacoste-Ferre, M.H.; Lacabanne, C.; Bochicchio, B.; Pepe, A. Conformational and thermal characterization of a synthetic peptidic fragment inspired from human tropoelastin: Signature of the amyloid fibers. Pathol. Biol. 2014, 62, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.L.; Nguyen, T.H.; Nguyen, D.H. Development and In Vitro Evaluation of Liposomes Using Soy Lecithin to Encapsulate Paclitaxel. Int. J. Biomater. 2017, 2017, 8234712. [Google Scholar] [CrossRef] [PubMed]
- Karmakar, S.; Maity, P.; Halder, A. Charge-Driven Interaction of Antimicrobial Peptide NK-2 with Phospholipid Membranes. ACS Omega 2017, 2, 8859–8867. [Google Scholar] [CrossRef] [PubMed]
- Almarwani, B.; Phambu, E.N.; Alexander, C.; Nguyen, H.A.T.; Phambu, N.; Sunda-Meya, A. Vesicles mimicking normal and cancer cell membranes exhibit differential responses to the cell-penetrating peptide Pep-1. Biochim. Biophys. Acta Biomembr. 2018, 1860, 1394–1402. [Google Scholar] [CrossRef] [PubMed]
- Phambu, N.; Almarwani, B.; Garcia, A.M.; Hamza, N.S.; Muhsen, A.; Baidoo, J.E.; Sunda-Meya, A. Chain length effect on the structure and stability of antimicrobial peptides of the (RW)(n) series. Biophys. Chem. 2017, 227, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Phambu, N.; Almarwani, B.; Alwadai, A.; Phambu, E.N.; Faciane, N.; Marion, C.; Sunda-Meya, A. Calorimetric and Spectroscopic Studies of the Effects of the Cell Penetrating Peptide Pep-1 and the Antimicrobial Peptide Combi-2 on Vesicles Mimicking Escherichia coli Membrane. Langmuir 2017, 33, 12908–12915. [Google Scholar] [CrossRef] [PubMed]
- Willumeit, R.; Kumpugdee, M.; Funari, S.S.; Lohner, K.; Navas, B.P.; Brandenburg, K.; Linser, S.; Andrä, J. Structural rearrangement of model membranes by the peptide antibiotic NK-2. Biochim. Biophys. Acta BBA Biomembr. 2005, 1669, 125–134. [Google Scholar] [CrossRef]
- Roeters, S.J.; Iyer, A.; Pletikapić, G.; Kogan, V.; Subramaniam, V.; Woutersen, S. Evidence for Intramolecular Antiparallel Beta-Sheet Structure in Alpha-Synuclein Fibrils from a Combination of Two-Dimensional Infrared Spectroscopy and Atomic Force Microscopy. Sci. Rep. 2017, 7, 41051. [Google Scholar] [CrossRef]
- Chen, S.W.; Drakulic, S.; Deas, E.; Ouberai, M.; Aprile, F.A.; Arranz, R.; Ness, S.; Roodveldt, C.; Guilliams, T.; De-Genst, E.J.; et al. Structural characterization of toxic oligomers that are kinetically trapped during α-synuclein fibril formation. Proc. Natl. Acad. Sci. USA 2015, 112, E1994–E2003. [Google Scholar] [CrossRef]
- Celej, M.S.; Sarroukh, R.; Goormaghtigh, E.; Fidelio, G.D.; Ruysschaert, J.-M.; Raussens, V. Toxic prefibrillar α-synuclein amyloid oligomers adopt a distinctive antiparallel β-sheet structure. Biochem. J. 2012, 443, 719–726. [Google Scholar] [CrossRef] [PubMed]
- Okada, Y.; Okubo, K.; Ikeda, K.; Yano, Y.; Hoshino, M.; Hayashi, Y.; Kiso, Y.; Itoh-Watanabe, H.; Naito, A.; Matsuzaki, K. Toxic Amyloid Tape: A Novel Mixed Antiparallel/Parallel β-Sheet Structure Formed by Amyloid β-Protein on GM1 Clusters. ACS Chem. Neurosci. 2019, 10, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Sayegh, R.S.R.; de Fátima Correia Batista, I.; de Melo, R.L.; Riske, K.A.; Daffre, S.; Montich, G.; da Silva Junior, P.I. Longipin: An Amyloid Antimicrobial Peptide from the Harvestman Acutisoma longipes (Arachnida: Opiliones) with Preferential Affinity for Anionic Vesicles. PLoS ONE 2016, 11, e0167953. [Google Scholar] [CrossRef] [PubMed]
- Schultz, Z.D.; Levin, I.W. Vibrational Spectroscopy of Biomembranes. Annu. Rev. Anal. Chem. 2011, 4, 343–366. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Bi, Y.; Zhang, H.; Dong, S.; Teng, L.; Lee, R.J.; Yang, Z. Cell-Penetrating Peptides in Diagnosis and Treatment of Human Diseases: From Preclinical Research to Clinical Application. Front. Pharmacol. 2020, 11, 697. [Google Scholar] [CrossRef]
- Alves, I.D.; Correia, I.; Jiao, C.Y.; Sachon, E.; Sagan, S.; Lavielle, S.; Tollin, G.; Chassaing, G. The interaction of cell-penetrating peptides with lipid model systems and subsequent lipid reorganization: Thermodynamic and structural characterization. J. Pept. Sci. 2009, 15, 200–209. [Google Scholar] [CrossRef]
- Li, G.; Huang, Y.; Feng, Q.; Chen, Y. Tryptophan as a Probe to Study the Anticancer Mechanism of Action and Specificity of α-Helical Anticancer Peptides. Molecules 2014, 19, 12224–12241. [Google Scholar] [CrossRef]
- dos Santos Cabrera, M.P.; Baldissera, G.; da Costa Silva-Gonçalves, L.; de Souza, B.M.; Riske, K.A.; Palma, M.S.; Ruggiero, J.R.; Arcisio-Miranda, M. Combining experimental evidence and molecular dynamic simulations to understand the mechanism of action of the antimicrobial octapeptide jelleine-I. Biochemistry 2014, 53, 4857–4868. [Google Scholar] [CrossRef]
- Graber, Z.T.; Thomas, J.; Johnson, E.; Gericke, A.; Kooijman, E.E. Effect of H-Bond Donor Lipids on Phosphatidylinositol-3,4,5-Trisphosphate Ionization and Clustering. Biophys. J. 2018, 114, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Wadhwani, P.; Epand, R.F.; Heidenreich, N.; Burck, J.; Ulrich, A.S.; Epand, R.M. Membrane-Active Peptides and the Clustering of Anionic Lipids. Biophys. J. Vol. 2012, 103, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Mangelschots, J.; Bibian, M.; Gardiner, J.; Waddington, L.; Van Wanseele, Y.; Van Eeckhaut, A.; Acevedo, M.M.D.; Van Mele, B.; Madder, A.; Hoogenboom, R.; et al. Mixed α/β-Peptides as a Class of Short Amphipathic Peptide Hydrogelators with Enhanced Proteolytic Stability. Biomacromolecules 2016, 17, 437–445. [Google Scholar] [CrossRef]
- Vincent, J.S.; Levin, I.W. Interaction of ferricytochrome c with cardiolipin multilayers: A resonance Raman study. J. Am. Chem. Soc. 1986, 108, 3551–3554. [Google Scholar] [CrossRef]
- Klaiss-Luna, M.C.; Jemioła-Rzemińska, M.; Strzałka, K.; Manrique-Moreno, M. Understanding the Biophysical Interaction of LTX-315 with Tumoral Model Membranes. Int. J. Mol. Sci. 2022, 24, 581. [Google Scholar] [CrossRef]
- Blume, A.; Huebner, W.; Messner, G. Fourier transform infrared spectroscopy of 13C:O labeled phospholipids hydrogen bonding to carbonyl groups. Biochemistry 1988, 27, 8239–8249. [Google Scholar] [CrossRef]
- Dyck, M.; Kerth, A.; Blume, A.; Lösche, M. Interaction of the Neurotransmitter, Neuropeptide Y, with Phospholipid Membranes: Infrared Spectroscopic Characterization at the Air/Water Interface†. J. Phys. Chem. B 2006, 110, 22152–22159. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | [Lipid/Peptide] Molar Ratio | Hydrodynamic Radius (nm) | Zeta Potential (mV) | Transition Temperature (°C) | ΔH (J/g) | Raman Intensity Ratio I1098/I1126 | Raman Intensity Ratio I1098/I1062 |
|---|---|---|---|---|---|---|---|
| PEN 10 μM | 34 ± 8 | 11.1 ± 2.3 | |||||
| C- 10 μM | 260 ± 4 | −24.8 ± 2.9 | 39.8 ± 0.6 | 6.59 ± 2.40 | 0.789 | 0.551 | |
| C- PEN | 1 | 399 ± 4 | −0.446 ± 1.2 | 39.9 ± 0.3 | 3.02 ± 1.20 | 0.846 | 0.55 |
| C- PEN | 5 | 222 ± 14 | −13.3 ± 1.3 | 40.2 ± 0.2 | 10.89 ± 2.20 | 0.645 | 0.513 |
| C- PEN | 10 | 304 ± 9 | −15.3 ± 2.7 | 39.1 ± 0.3 | 18.31 ± 1.90 | 0.671 | 0.472 |
| C- PEN | 50 | 241 ± 7 | −22.4 ± 3.1 | 40.0 ± 0.4 | 28.1 ± 3.10 | 0.864 | 0.691 |
| C- PEN | 110 | 258 ± 2 | −16.8 ± 2.4 | 40.5 ± 0.2 | 11.66 ± 1.40 | 1.136 | 1.042 |
| Name | [Lipid/Peptide] Molar Ratio | Hydrodynamic Radius (nm) | Zeta Potential (mV) | Transition Temperature (°C) | ΔH | Raman Intensity Ratio I1098/I1126 | Raman Intensity Ratio I1098/I1062 |
|---|---|---|---|---|---|---|---|
| PEN 10 μM | 34 ± 8 | 11.1 ± 2.3 | |||||
| H- 10 μM | 395 ± 4 | −0.75 ± 1.78 | 40.1 ± 0.5 | 5.79 ± 1.80 | |||
| H- PEN | 1 | 342 ± 2 | 6.17 ± 0.40 | 40.1 ± 0.4 | 5.04 ± 0.80 | 0.729 | 0.526 |
| H- PEN | 5 | 593 ± 3 | 3.23 ± 1.10 | 40.2 ± 0.2 | 3.85 ± 1.10 | 0.814 | 0.588 |
| H- PEN | 10 | 503 ± 11 | 3.64 ± 0.80 | 40.0 ± 0.2 | 6.49 ± 0.90 | 0.783 | 0.557 |
| H- PEN | 50 | 487 ± 17 | 2.23 ± 1.50 | 40.2 ± 0.6 | 3.84 ± 1.20 | 0.741 | 0.571 |
| H- PEN | 110 | 563 ± 4 | 1.313 ± 0.637 | 47.5 ± 0.5 | 5.93 ± 0.40 | 0.753 | 0.617 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almarwani, B.; Hamada, Y.Z.; Phambu, N.; Sunda-Meya, A. Investigating the Insertion Mechanism of Cell-Penetrating Peptide Penetratin into Cell Membranes: Implications for Targeted Drug Delivery. Biophysica 2023, 3, 620-635. https://doi.org/10.3390/biophysica3040042
Almarwani B, Hamada YZ, Phambu N, Sunda-Meya A. Investigating the Insertion Mechanism of Cell-Penetrating Peptide Penetratin into Cell Membranes: Implications for Targeted Drug Delivery. Biophysica. 2023; 3(4):620-635. https://doi.org/10.3390/biophysica3040042
Chicago/Turabian StyleAlmarwani, Bashiyar, Yahia Z. Hamada, Nsoki Phambu, and Anderson Sunda-Meya. 2023. "Investigating the Insertion Mechanism of Cell-Penetrating Peptide Penetratin into Cell Membranes: Implications for Targeted Drug Delivery" Biophysica 3, no. 4: 620-635. https://doi.org/10.3390/biophysica3040042
APA StyleAlmarwani, B., Hamada, Y. Z., Phambu, N., & Sunda-Meya, A. (2023). Investigating the Insertion Mechanism of Cell-Penetrating Peptide Penetratin into Cell Membranes: Implications for Targeted Drug Delivery. Biophysica, 3(4), 620-635. https://doi.org/10.3390/biophysica3040042