Abstract
Physical roots, exemplifications and consequences of periodic and aperiodic ordering (represented by Fibonacci series) in biological systems are discussed. The physical and biological roots and role of symmetry and asymmetry appearing in biological patterns are addressed. A generalization of the Curie–Neumann principle as applied to biological objects is presented, briefly summarized as: “asymmetry is what creates a biological phenomenon”. The “top-down” and “bottom-up” approaches to the explanation of symmetry in organisms are presented and discussed in detail. The “top-down” approach implies that the symmetry of the biological structure follows the symmetry of the media in which this structure is functioning; the “bottom-up” approach, in turn, accepts that the symmetry of biological structures emerges from the symmetry of molecules constituting the structure. A diversity of mathematical measures applicable for quantification of order in biological patterns is introduced. The continuous, Shannon and Voronoi measures of symmetry/ordering and their application to biological objects are addressed. The fine structure of the notion of “order” is discussed. Informational/algorithmic roots of order inherent in the biological systems are considered. Ordered/symmetrical patterns provide an economy of biological information, necessary for the algorithmic description of a biological entity. The application of the Landauer principle bridging physics and theory of information to the biological systems is discussed.
1. Introduction
Biological objects demonstrate remarkably repeatable patterns, governed by simple mathematical laws and regularities. Biological systems frequently exhibit symmetry and regularity on various spatial scales, starting from the genomic level and biomolecules and extending to the entire organism [1,2,3,4,5,6]. A nearly universal observation, which was reported recently, states the subunits in protein assemblies are arranged in symmetric ways (the subunits themselves may be not symmetrical) [7]. It was hypothesized that the beautiful symmetries of biomolecules may reflect basic principles about the energy landscape in biology, just as symmetry relations do in particle physics [8]. Symmetry is inherent to the bodies of practically all animals (with rare exceptions). Animals are characterized by some kind of overall body symmetry, and these are of only a few types: translational, radial, biradial and bilateral symmetry [9]. Symmetry, in turn, represents a kind of ordering in physical and biological systems (“ordering” is understood as the process of putting elements of a biological pattern in a particular order) [10,11]. When we address the symmetry of ordered patterns, we usually restrict ourselves to mainly considering periodic order [12]. At the same time, order without periodicity has emerged to properly describe an increasing number of complex systems, and in particular biological ones [12]. This kind of ordering was referred to as aperiodic ordering [12]. An outstanding example of aperiodic ordering is supplied by the Fibonacci numbers, or Fibonacci series [13,14,15,16]. Fibonacci and Lucas series appear in biological patterns [17]. Perhaps the most striking example of such samples is supplied by phyllotaxis, which is the arrangement of leaves on a plant stem [13,14,15,16,17]. These patterns are surprisingly regular, so regular in fact that a physicist can compare their order to that of crystals. However, ordering in biological systems is usually not perfect, and quantitative measures of the deviation from the perfect ordering, which were introduced recently, will be discussed below in detail. It should be mentioned that non-ordered, asymmetrical biological systems exist. It is generally agreed that sponges are completely asymmetrical (see Figure 1); however, this thesis may be debated and will be addressed below in detail.

Figure 1.
Calcareous sponges are depicted. Sponges are usually regarded as non-ordered biological objects.
Moreover, it was suggested that breaking symmetry is a prevalent process in biology [3,18]. However, to be broken, symmetry in biological patterns must first appear, and it appears on different levels of organization of biological systems [1,2,4,18]. First of all, let us define rigorously the notion of "symmetry". Symmetry in the text is understood as invariance of the biological object under some mathematical transformations, which may be: translation, reflection, rotation or scaling (including fractal scaling).
A reasonable question is as follows: what is the biological reasoning for periodic and aperiodic ordering in biological systems? In other words, why does nature prefer ordered patterns? The possible answers to this fundamental question may be classified as follows: (i) The appearance of symmetry and other sample patterns is due to the external physical constraints implied on the biological system [5,19,20,21]. This hypothesis accepts that just physical effects, which in many cases act as proximate, direct, tissue-shaping factors during ontogenesis, are also the ultimate causes, (in other words) the indirect factors that provide a selective advantage, of animal or plant symmetry, from organs to body plan level patterns [19,20,21]. These physical constraints, in turn, may be responsible for the time evolution of biological species. In particular, it was suggested that the last common ancestor of metazoans (multicellular animals) was probably cylindrically symmetrical, with a concomitant axial polarity generated by early embryonic signaling. Changes in symmetry type occurred repeatedly with time, with respect to environmental parameters as the main constraining forces, providing an evolutionary change in the macroscopic symmetry of metazoans [21]. (ii) The second idea explaining the abundance of the symmetric patterns in biology implies that the symmetry of biological systems stems from the symmetry of molecules themselves and potentials describing interactions between molecules [22,23]. It was demonstrated that the symmetry of these potentials governs the symmetry of biological systems, such as actin, tubulin and the ubiquitous icosahedral shell structures of viral capsids [22,23]. (iii) The third approach relates the appearance of mathematical ordering in biological systems to pure survival reasons. For example, periodic cicadas emerge from their underground homes to mate every 13 or 17 years, and 13 and 17 are primes (this kind of temporal ordering also represents aperiodic ordering) [24,25,26,27,28,29]. The philosophy of the evolutionary-based explanation of the mathematical ordering is that if cicadas have 12-year cycles, all the predators with 2-, 3-, 4-, and 6-year cycles will eat them; in other words, the cicadas with prime number cycles will have a higher probability of survival. Rigorously speaking, cicadas will leave more offspring if their cycles are described by primes [28,29]. Let us quote [29]: “a prey with a 12-year cycle will meet-every time it appears-properly synchronized predators appearing every 1, 2, 3, 4, 6 or 12 years, whereas a mutant with a 13-year period has the advantage of being subject to fewer predators”. A second explanation, proposed by Cox, Carlton and Yoshimura, concerns the avoidance not of predators but of hybridization with similar subspecies (consider that these explanations are not mutually exclusive) [25,26,27]. Genuine reasoning for the prime-shaped life cycle of cicadas remains debatable, and the discussion of this reasoning gave rise to the deep philosophical discussion of the nature and roots of the notion of “explanation” of natural/biological phenomena in [30]. Survival/reproductive reasons were also involved in the explanation of symmetry appearing in the color of zebra finches, shown in Figure 2.

Figure 2.
Zebra finch is depicted. Symmetrically banded males and females are preferred by an individual of the opposite sex.
It was demonstrated that symmetrically banded males produced more offspring that survived past the period of parental care than males in either of the asymmetric treatments. This appeared to be the effect of female choice processes and female-based parental investment and not male intra-sexual dominance. Thus, it was shown that symmetrically manipulated males gain reproductive advantages in controlled laboratory conditions, which further supports recent theories indicating the evolutionary importance of symmetry in signaling-trait design [31]. Moreover, it was demonstrated that symmetric patterns are attractive not only to females, and it was found that males associated more with symmetrical than asymmetrical females, indicating a preference for symmetry [32]. (iv) Finally, we discuss the more hypothetical relation of the appearance of symmetrical patterns in biological systems to informational reasoning. It was suggested that symmetric biological structures and patterns preferentially arise not only due to natural selection but also because they require less specific information to encode and are therefore much more likely to appear as phenotypic variation through random mutations [33]. This novel concept, which is well tailored to the general informational paradigm of exact sciences and was criticized recently in [34], will be discussed below in relation to the Curie–Minnigerode–Neumann and Landauer principles [35,36,37]. In our paper, we focus on the physical and informational reasoning of symmetry/ordering inherent in biological systems. We will also demonstrate that the notion of ordering has a “fine structure” and hardly could be quantified unequivocally with a single mathematical parameter.
2. Physical Reasoning for the Abundant Symmetry of Biological Patterns
2.1. The “Top-Down” Approach to the Explanation of Symmetry in Organisms: Symmetry Is Dictated by the Properties of Media in Which the Organisms Act
Physical reasoning involved in the explanation of symmetry of biological objects may be classified according to the following main approaches:
- (i)
- The “top-down” approach, implying that the symmetry of the biological structure follows the symmetry of the media in which this structure is functioning;
- (ii)
- The “bottom-up” approach, assuming that the symmetry of biological structures emerges from the symmetry of molecules constituting the structure.
Let us start with the “top-down” pure physical reasoning which may be responsible for the symmetry abundant in the plant and animal realms. The two main symmetries that can be observed in the animal body plan are radial and bilateral, as illustrated in Figure 3 [21,38,39].
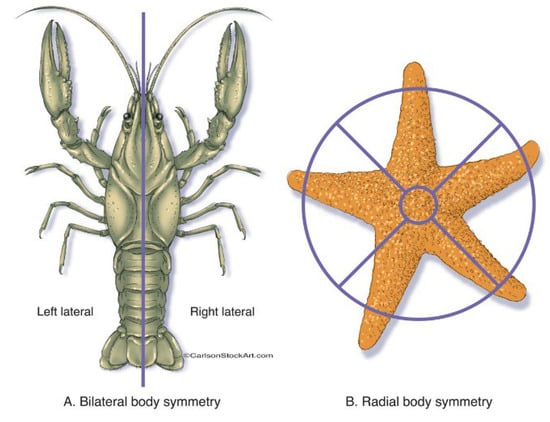
Figure 3.
Bilateral (A) and radial symmetry (B) in biological objects.
Pure physical “demystification” of animal symmetry was suggested in [19,20]. Let us quote [19]: “symmetry is a response in the geometry of the ‘living matter’ to physical forces”. As an example of such a pathway of thinking, consider the directed locomotion of sea inhabitants. The most comprehensive idea which explains how directed locomotion is favored by bilateral symmetry comes from a theoretical paper [18], which argued that bilateral symmetry is favorable for maneuverable locomotion in the macroscopic world (in which inertial forces dominate over viscous forces, i.e., in the high Reynolds numbers’ realm) because bilateral is the only type of symmetry which is streamlined in only one direction while being non-streamlined in others [18]. The symmetry of biological entities which inhabit water and air becomes more clear when the ideas developed in [40] are taken into account. It was concluded in [40] that a large proportion of fish species have developed compressed, elliptical body cross-sectional shapes. This shape is justified from the hydrodynamic point of view, as the basic issue of minimizing drag for a given volume would result in a body of revolution [40]. Thus, this adaptation must have had very significant advantages to have reappeared so many times. One such advantage is clear from the use of the body as a propulsor, as the sideways oscillations of a vertically compressed body will produce much larger forces, but this also reduces rolling, which would, in turn, induce yawing moments due to the difference in angle of attack on the pectorals and the angle of attack on the caudal and dorso-anal fins produced by rolling. Thus, vertical compression is a stabilizing development [40,41].
We propose the following development of the suggested pure “physical explanation” of symmetry inherent for fishes, birds and flying insects. The observed symmetry is to a great extent explained by the isotropy of physical properties of gases and liquids in which fishes and birds act. Gases and some liquids are essentially isotropic media. Gases are isotopic due to the random motion of molecules constituting the gaseous phase (also called the molecular chaos hypothesis), implying that the velocities of colliding molecules are uncorrelated and independent of position [42,43,44,45]. The hypothesis of molecular chaos also implies the equipartition theorem serving as a basis for classical thermodynamics and statistical physics, which states that in thermal equilibrium, energy is shared equally among all of its various forms [46,47,48,49,50]. The equipartition theorem, in turn, is grounded on two main assumptions: (1) the classical version of the canonical probability distribution is applicable and adequate; (2) the classical expression for the total energy of the particle splits additively into two parts: one part depends quadratically on a single variable (say x), and the other (denoted is entirely independent of that variable It is easily seen that the equipartition of energy holds whatever the value of the constant a is; moreover, the equipartition theorem holds for a broad class of potentials , as discussed in [51]. Thus, symmetry of bodies of birds and flying insects possibly emerges from isotropy of air accompanied by equipartition of energy, which is true for air molecules. This “physically grounded” approach may be stretched even further: actually, conservation laws appearing in physics arise from the fundamental symmetries of space-time; thus, symmetry recognized in physical systems reflects these fundamental symmetries; hence, it is plausible to suggest that the symmetry of biological systems also arises from the fundamental space-time symmetries.
The situation with liquids is much more complicated; indeed, liquids may be isotropic and non-isotropic (liquid crystals) [52]. However, the most important biological liquid is water, which is considered isotropic in biophysical models which deal with water and water inhabitants. This assumption, as a matter of fact, is far from trivial. It was demonstrated that the state of water in the body constituents of living organisms and in the vicinity of biological macromolecules differs significantly from the state of water in solutions of simple molecules and in pure water [53,54]. Biological macromolecules induce a characteristic water structure in their close vicinity due to weak macromolecular-water interactions [53,54]. Considering water as an isotropic medium is justified on the macroscopic level of treatment of biological systems. On the other hand, aqueous solutions of DNA govern their chirality, i.e., the lack of mirror symmetry of the solutions, which already brings us to the “bottom-up” approach to the symmetry of biological objects, implying that the symmetry of biological structures emerges from the symmetry of molecules constituting the structure [55,56,57,58,59,60].
2.2. “Bottom-Up” Approach to the Symmetry of Biological Systems, Mathematical Measures of Order in Biological Patterns and the Curie-Neumann Principle
2.2.1. Mathematical Measures of Symmetry and Ordering in Biological Patterns
As we already mentioned in the previous sections, DNA constitutes the symmetry properties (chirality) of aqueous solutions [55,56,57,58,59]. In addition, the tobacco mosaic virus governs the lyotropic liquid crystalline behavior of its viral ensemble [60]. Thus, we come to the “bottom-up” approach to the symmetry of biological structures and patterns, accepting that the symmetry of large-scale, macroscopic biological structures emerges from the symmetry of molecules constituting the structure. High-symmetry assemblies of proteins, resembling those inherent to the geometries of the Platonic solids, were discussed in 7. The symmetry of biological macromolecules in comparison with the symmetry of nuclei, atoms and elementary particles was deeply analyzed in 8. Let us quote from 8: “The role of symmetry in the physics of atoms, nuclei, and elementary particles is different from its role in the biological world. In microphysics, it has been fruitful to postulate symmetry as being fundamental, thereby severely constraining the form of the underlying laws. Furthermore, the consequences of symmetry for dynamics and stability are profound in the subatomic world. General treatment of these symmetry effects, based on group theory, allows a nearly complete classification of states and transitions. Indeed, even the deviations from exact symmetry have their own quantifiable consequences. In the biological world, symmetry exists too, but it often appears to be an accident. While accidents still can have significant consequences, rationalizable on a case-by-case basis, there has been no general unifying theory of symmetry in biology. Exact symmetry in biology would even seem to be antithetical to the notions of complexity, variety, and metamorphosis that are central to the idea of life, as we know it. Nevertheless, as in microphysics, life requires stability and sameness as well as change For a particular protein, biomolecular symmetry will not be exact unless it is the result of gene duplication. It is a quantitative issue how much the forces leading to minimal frustration, partially determined by symmetry considerations, are dominant over the aspects of the landscape arising from randomness”.
Indeed, the exact, perfect symmetry (in the rigorous mathematical sense) is rare in the realm of biology. It is sparse on different levels of biological structures, and even on the molecular level, it is not abundant. Exact symmetry may be inherent for “averaged” biological species; however, the rigorous mathematical definition of “average” becomes non-trivial for biological entities, which evolve with time. Now we come to the novel ideas that appeared in the theory of symmetry in the last decades. Until now, it was usually latently accepted that symmetry changes abruptly or intermittently, i.e., symmetry is an exact feature of the pattern. In other words, symmetry is usually viewed as a binary feature, where an object is considered as either perfectly symmetric or asymmetric. Despite this, a fundamentally new approach to quantifying symmetry as a continuous measure of a pattern was suggested and tested by Avnir, Zabrodsky and co-workers [61,62,63,64,65]. The notion of the “continuous measure of symmetry” was introduced and developed [61,62,63,64,65]. The continuous measure of symmetry is a fundamentally new approach to quantifying symmetry [61,62,63,64,65]. The continuous measure of symmetry is quantified by the sum of minimum squared distances that are required to move the points of an original shape in order to obtain a symmetrical shape [61,62,63,64,65]. The continuous measure broke the binary “yes–no” paradigm traditionally used for the analysis of the symmetry of patterns [61,62,63,64,65]. This approach was successfully exploited for quantification of the symmetry of electronic wave functions [66] and quasicrystals [67]. The continuous measure of symmetry was recently applied to the analysis of the symmetry of proteins, as well as plant leaves and other structures [68,69]. It was revealed that symmetry deviations of proteins are by far higher in solution, compared to the crystalline state [69]. However, only the first results in the field of the continuous analysis of the symmetry of biological objects were reported, and it seems that the mathematical apparatus of the continuous measure of symmetry has enormous potential in mathematical biology.
It should be emphasized that the continuous measure of symmetry is not a single mathematical value enabling the quantification of order in biological patterns. The alternative measure of order in the biological patterns emerges from the Voronoi tessellation (or Voronoi diagram). A Voronoi tessellation of an infinite plane is a partitioning of the plane into regions based on the distance to a specified discrete set of points (called seeds or nuclei and shown with black squares in Figure 4). For each seed, there is a corresponding region, consisting of all points closer to that seed than to any other [70,71,72,73,74,75,76,77].
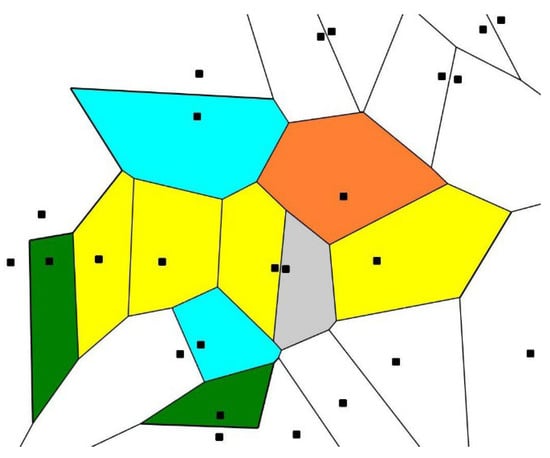
Figure 4.
Voronoi diagram (tessellation) emerging from the initial 25 randomly placed points, also called seeds (represented with black squares); green polygons are quadrangles; yellow polygons are pentagons; the grey polygon is a hexagon; blue polygons are heptagons; the orange polygon is an octagon. Are the seeds ordered? The answer to this question is supplied by calculation of the Voronoi (Shannon) entropy of the pattern.
The Voronoi (Shannon) entropy of a given set of points located in a plane is given by:
where is the probability of finding an n-sided Voronoi polygon within a given Voronoi tessellation and i is the total number of polygon types with different numbers of edges [24,25,26]. The summation in Equation (1) is performed from i = 3 (the smallest possible polygon—a triangle) to the largest coordination number of the polygon; e.g., for an octagon, the largest value of i is 8. Consider the Voronoi diagram built of pentagons only; in such a diagram, and consequently . It is reasonable to relate to such a diagram the maximal degree of order; in contrast, the Voronoi entropy of the random pattern of seeds was established as . Seeds may be displaced continuously from their initial locations, and the Voronoi entropy, in turn, will be changed continuously. Voronoi tessellations were already successfully applied for the analysis of ordering in biological systems, namely proteins [73], cell dynamics [74] and ordering of GABAA receptors in hippocampal inhibitory synapses [75].
Thus, we already have two alternative continuous measures of order in biological systems, namely the continuous measure of symmetry [68,69] and the Voronoi Entropy (and this is not an exhaustive list) [73,74,75,76,77]. Thus, a reasonable question immediately arises: are these measures correlated? It was recently demonstrated that the continuous measure of symmetry and the Voronoi entropy of the pattern are not necessarily correlated; moreover, the anti-correlation of these values was registered [67,78]. Alternative measures of ordering in biological patterns, such as Minkowski functionals, were also introduced [79,80]. A Shannon (informational) measure of symmetry was also suggested recently [81]. Consider a 2D pattern built of 1D and/or 2D shapes or lines, demonstrating a number of symmetry elements (rotational symmetry, centers of symmetry, axes of symmetry, etc.), denoted , , where k is a number of non-identical symmetry operations [81]. Elements form the symmetry group of the shape G (which should be clearly distinguished from the symmetry group of the entire pattern). Thus, the informational measure of the symmetry of the pattern is defined in a Shannon-like form (compare with Equation (2)) as:
where is the probability of appearance of the symmetry operation within the shapes (lines) constituting the pattern, defined as:
where = is the total number of symmetry elements (operations) appearing in the 1D or 2D shapes, recognized in a given pattern, and is the number of the same symmetry elements (operations) calculated for a given pattern. The normalization condition given by Equation (4) is applied:
The informational (Shannon) measure of symmetry, defined by Equation (2), is applicable for mixed patterns built of curves and shapes, and thus it is suitable for the analysis of biological patterns (such as those depicted in Figure 5). We necessarily conclude that the notion of “ordering in biology patterns” has a fine structure, and the correlation between various mathematical measures of order remains an open problem and should be elucidated in future research.
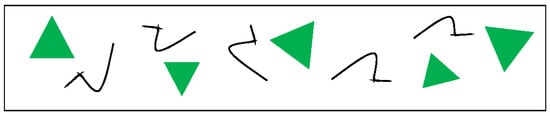
Figure 5.
Mixed pattern built of p = 5 irregular non-symmetric curves and p = 5 equilateral triangles. The symmetry group of the equilateral triangle is the dihedral symmetry group containing 3 p symmetry axes and 3 p rotations (including the rotation, denoted ). One more operation comes from the irregular curves; thus, we have in total 7 p symmetry operations in this pattern. The IMS is easily calculated according to Equation (2): . For details, see [81].
2.2.2. Bottom-Up Approach to the Symmetry of Biological Systems and the Curie–Neumann Principle
Obviously, the properties of biomolecules influence the properties of macroscopic biological objects. However, what is the interrelation between the symmetry of biomolecules and the eventual properties of macroscopic structures? In the realm of crystallography, this interrelation is constituted by the Curie–Minnigerode–Neumann principle, which in its simplest form was formulated by Minnigerode as “the group of the structure of a crystal is contained in the group of each of its physical properties”, or in other words, all elements of symmetry of a crystal are at the same time elements of symmetry of its macroscopic properties [82,83]. This statement may be mathematically expressed as follows:
which is understood as follows: for a physical property to be allowed to exist within an object, it is a necessary but not sufficient condition that the group of the symmetry operations of the object be at least a subgroup of the group of the symmetry operations of the physical property , or alternatively, the tensor describing a physical property (the so-called “property tensor”) has to be invariant against all symmetry operations of the object (crystal), or alternatively, the symmetry elements of the causes must be found in their effects, but the converse is not true; that is, the effects can be more symmetric than the causes [82,83]. However, Curie already clearly understood the role of asymmetry in constituting the physical phenomena. The phenomenon of piezoelectricity emerges from the symmetry breaking in certain crystals [84]. This idea was aphoristically formulated by Curie as follows: “asymmetry is what creates a phenomenon”. And it seems that this approach may be extended to biology; however, there is a long way ahead in order to understand the role of asymmetry in constituting biological phenomena.
The paradoxical reduction in crystal symmetry at the textural level, without a concomitant reduction in the intrinsic symmetry of the crystal structure, was observed for calcitic sponge (Clathrina conrorta and Kebira ufroides) spicules (see Section 1). The effect, opposing the Curie–Minnigerode–Neumann principle, was related to the addition of foreign macromolecules to a growing crystal, occurring at surfaces [85].
3. Informational Reasoning for Symmetry in Biological Systems
3.1. Symmetry and Order in Biological Systems Have Informational/Algorithmic Roots
Alternative reasoning for the abundance of symmetrical patterns in biological systems was suggested in [33]. The authors of [33] noted that it is plausible to assume (by a certain analogy to engineering design) that symmetry may stem from natural selection, as discussed in Section 2.1, in which it was demonstrated that bilateral symmetry of sea inhabitants is favorable for their maneuverable locomotion in water [19]. However, evolution, unlike engineers, cannot plan ahead, and so these symmetrical features must also afford some immediate selective advantage which is hard to reconcile with the breadth of systems where symmetry is observed. It was suggested in [33] that the symmetric structures preferentially arise not only due to natural selection but also because they require less specific information (and consequently less energy according to the Landauer principle [35,36,37]) to encode and are therefore much more likely to appear as phenotypic variation through random mutations. Arguments from algorithmic information theory enabled the formalization of this hypothesis, leading to the prediction that many genotype–phenotype maps are exponentially biased toward phenotypes with low descriptional complexity (preference for symmetry is a special case of the bias toward compressible descriptions [86]). The authors of the aforementioned hypothesis validated the predictions of this idea with biological data, showing that protein complexes, RNA secondary structures and a model gene regulatory network all exhibit the expected exponential bias toward simpler (and more symmetric) phenotypes [33]. The authors of [33] supplied arguments supporting their concept and rooted in the algorithmic information theory, in which it is well accepted that when the space of algorithms is considered, outputs that can be generated by short programs are exponentially more likely to be produced than outputs that can only be generated by long programs. It was demonstrated in [33] that formalism developed in the algorithmic information theory may be successfully applied for the analysis of genotype–phenotype maps. Thus, symmetry appearing in biological systems emerges from the “informational arguments”, providing an economy of biological information necessary for the description of a biological entity [33,86].
The “informational biological paradigm” introduced and developed in [33] was recently criticized in [34], in which the role of symmetry breaking in biological systems was stressed. It was noted that while symmetry may arise more commonly in biological structures with low complexity, there is evolutionary pressure to develop asymmetry in many biological structures with high complexity. The emergence of symmetry cannot be fully understood without considering the emergence of asymmetry as well [34]. Consider, for example, the human brain, one of the most complex and mysterious biological structures [34,87]. While the two halves of the brain look roughly symmetric at first glance, a recent large-scale neuroimaging demonstrated that structural left–right asymmetries are the rule, rather than the exception, for cortical brain areas [87]. Importantly, the human central nervous system is not the only one that shows such striking asymmetries. Breaking symmetry is therefore a crucial step in the development of all nervous systems [34]. This statement is in striking correspondence with the Curie–Minnigerode–Neumann principle, formulated by Curie as follows: “asymmetry is what creates a phenomenon” [82,83], which may reshaped as follows: “asymmetry is what creates a biological phenomenon”.
The explanation of symmetrical patterns abundant in biological systems with arguments rooted in the algorithmic information theory seems deep and promising. We propose stretching this approach to the grounding of other kinds of periodic and aperiodic ordering appearing in biological systems, such as Fibonacci series and Archimedean and Lucas spirals [13,14,15,16,17,88]. Indeed, the Fibonacci series found in phenotypic structures of plants and animals, defined by Equation (6).
and the Archimedean spiral, defined by Equation (7).
represent examples of simple and informationally effective mathematical regularities, which may be specified by short algorithms. The authors of [88] reported a model of the cell division implying asymmetric cell division. In the model, cells divide asymmetrically to generate a mature and an immature cell [88]. The model output on the number of cells generated over time fits specific Fibonacci p-number sequences depending on the maturation time [88]. Thus, the relation of the Fibonacci series to the asymmetry of biological processes became elucidated [88].
3.2. Symmetry and Ordering in Biological Systems and the Landauer Principle: Informational Paradigm of Biology
The idea that symmetry in biological patterns is deeply rooted in the informational basic structure of reality fits with ideas introduced by John Archibald Wheeler, who suggested that fundamentals of physics should be re-built on the informational groundings and assumed that the main notions of physics are deeply rooted in the “bit-based” scientific paradigm [89]. This approach may be very briefly and aphoristically summarized as follows: “all physical things are information-theoretic in origin”, aphoristically reduced to “it from bit” [89]. The idea was developed recently within the highly debated and controversial Landauer principle, suggesting the thermodynamic equivalent of information, establishing the lower theoretical limit of energy consumption of computation [35,36]. It holds that “any logically irreversible manipulation of information, such as the erasure of a bit or the merging of two computation paths, must be accompanied by a corresponding entropy increase in non-information-bearing degrees of freedom of the information-processing apparatus or its environment” [35,36]. In other words, there is a minimum possible amount of energy E required to erase one bit of information, known as the Landauer limit and supplied by Equation (8):
where is the Boltzmann constant and T is the absolute temperature of the heat sink [90,91,92,93]. The Landauer principle was experimentally tested in [94,95]. Extensions of the Landauer principle to the realms of quantum mechanics [96] and general relativity [97] were reported. The Landauer principle applied to mechanical motion demonstrates that dissipation of energy is the key process through which mechanical motion becomes observable [98]. The analysis of the performance of photon detectors (such as eyes) leads to the conclusion that only efficiency is limited by the Landauer energy bounds on information gain and information erasure [99]. Estimation of information contained in molecular motion based on the Landauer principle was performed in [100]. The Landauer principle restricts the informational capacity of biological systems; thus, it is closely related to the abundance of ordering in biological systems; indeed, periodic and aperiodic ordering enable the saving of memory/energy available for the organism. The Landauer principle bridges the informational and thermodynamic paradigms of life, which explains the ability of organisms to maintain low levels of entropy that explain order [101]. The informational paradigm of life enabled the analysis of the SARS-CoV-2 virus using Shannon’s information theory [102]. A relationship between the information entropy of genomes and their mutation dynamics was established. In particular, it was revealed that genomes undergo genetic mutations over time driven by a tendency to reduce their overall information entropy [102]. Let us roughly estimate the informational capacity of living cells with the Landauer principle. Consider that the characteristic spatial range of living cells, namely, , spans the dimensions of a majority of prokaryotic and eukaryotic cells [102]. Thus, the maximal informational capacity of a living cell may be estimated, according to the Landauer principle, with Equation (9); if we speculate that information exchange occurs only via the surface of a living cell we estimate the following:
where and the total and specific interfacial energies of a cell. Assuming yields [103]. (This value should not be confused with the genomic capacity of a cell [104,105].) Thus, we may estimate the informational interfacial capacity of a small micro-scaled cell as bits and compare this with the DNA-based code, which enables the storage of bits of information [106]. Thus, the informational capacity of DNA and cells is restricted; hence, assessing the thermodynamic efficiency of the computations performed by organisms becomes crucial. The authors of [107] posed and addressed the following fundamental question: how close has life come to maximally efficient computation (presumably under the pressure of natural selection)? The suggested answer is summarized as follows: despite inevitable shifts across the architectures of life, the authors revealed a surprising consistency in the efficiency of translation, one of the most universal types of computation carried out in biological systems [107]. The analyses demonstrated that as bacteria become larger, their overall translational efficiency converges on that of a single ribosome [107]. In addition, this efficiency is maintained for unicellular eukaryote and mammalian cells [107]. Astonishingly, this efficiency is only about an order of magnitude larger than the Landauer bound, supplied by Equation (8) (see [107]). It is instructive to compare the efficiency of biological computations to that of the best supercomputers. Actually, the cost of computation in supercomputers is about eight orders of magnitude worse than the Landauer bound given by Equation (8), which is about six orders of magnitude less efficient than biological translation when both are compared to the appropriate Landauer bound [107]. Biology is beating our current engineered computational thermodynamic efficiencies by an astonishing degree [107].
It should be emphasized that symmetry and order (periodic and aperiodic) inherent in biological systems improve the efficiency of biological computation; indeed, when an n-fold symmetry is present, the single computation act governs the location of a number of n “spots” in the biological pattern. An interface between artificially created digital information and information produced by organisms was addressed in 108. It was demonstrated that human-related digital information has reached a similar magnitude to information in the biosphere [108].
Information aspects of order in DNA were discussed in [12]. When the structure of DNA was discovered, it was first assumed that the basic repeating unit of DNA polymer was a tetranucleotide, in which the four different kinds of nucleotides recurred in regular sequence [12]. Thus, DNA was originally viewed as a trivially periodic macromolecule, characterized by translational symmetry, which is unable to store the amount of information required for the governance of cell function [12]. It was Schrödinger who suggested that genetic material should consist of a long sequence of a few repeating elements exhibiting a well-defined order without the recourse of periodic repetition [109]. In this way, the notion of a 1D aperiodic crystal was introduced and the deep connection between symmetry and information was established [12,109]. Again, the correspondence: “more symmetry–less information” takes place.
4. Conclusions
The concept of symmetry became one of the central concepts of mathematical biology, as already occurred in physics [110]. Biological systems demonstrate remarkable symmetry and ordering which may be periodic or aperiodic (represented by the Fibonacci series, describing seed spirals on the sunflower’s face and primes appearing in the lifecycle of cicadas) [1,2,4,12,17,24,25,26,27,28]. Various types of order and symmetry span all biological spatial scales starting from biomolecules [7,8] and reaching entire organisms and the biological patterns created by organisms [9,19,20,21]. The two main symmetries that can be observed in the animal body plan are radial and bilateral, [21,38,39]. The eggs of fucoid brown algae such as Fucus spp. and Silvetia spp. (formerly Pelvetia) demonstrate spherical symmetry [111]. The unfertilized eggs have no cell wall and have a spherical shape, and their cytological components are evenly distributed [111]. Fractal symmetry was registered for sponges [112]. The symmetry may be even regenerated with time, as occurs with damaged jellyfish [113]. The challenging and fundamental question is as follows: what is the physical and biological reasoning of periodic and aperiodic ordering in biological systems? In other words, why does living nature prefer ordered/symmetrical patterns? The possible answers to this fundamental question may be summarized as follows: (i) The appearance of symmetry/ordering is due to the external physical constraints implied on the biological system [5,19,20,21]. Physical factors/constraints follow the basic conservation laws emerging, in turn, from the space-time symmetries [110]. Thus, it may be hypothesized that the symmetry of biological systems also at least partially arises from the fundamental space-time symmetries. (ii) The second hypothesis explaining the abundance of the highly ordered patterns in biology accepts that the symmetry of biological systems emerges from the symmetry of molecules themselves and potentials describing interactions between molecules [22,23]. It was reported recently that roughly 40% of the biomolecules registered in the Protein Data Bank exhibit some type of symmetry, including formal global symmetry, local symmetry or pseudo-symmetry (many proteins are built from asymmetrical subunits; however, the entire protein may be symmetrical) [114]. (iii) The third approach relates the appearance of mathematical ordering in biological systems to the pure survival, evolutionary reasons [24,25,26,27,28,29]. (iv) The most recent hypothesis relates the appearance of symmetrical patterns in biological systems to informational reasoning [33]. It was suggested that symmetric biological structures and patterns preferentially arise not only due to natural selection but also because they require less specific information (and less energy) to encode [33]. We addressed this approach within the general informational scientific paradigm, proposed by John Archibald Wheeler, who suggested that the fundamentals of physics should be re-built on the informational groundings and assumed that the main notions of physics are deeply rooted in the “bit-based” scientific paradigm [89]. The ideas discussed in [33] stretch this approach to the “bit-based” biological paradigm, which may be formulated as follows: economy of information is essential in the constitution of biological patterns. Thus, it becomes understandable why only the simplest mathematical regularities such as Fibonacci series [13,14,15,16,17,89] and symmetrical patterns are abundant in biology; these regularities decrease the algorithmic complexity of biological systems [33]. The informational paradigm of biology is closely related to the Landauer principle, bridging the theory of information to physics and suggesting the thermodynamic equivalent of information, under establishing the lower theoretical limit of energy consumption of physical/biological computation [35,36,37,90,91,92,93,94,95,96,97,98,99,100]. The analyses of the computational efficiency of bacteria demonstrated that as bacteria become larger, their overall translational efficiency converges on that of a single ribosome [106]. Moreover, this efficiency is universal for unicellular eukaryote and mammalian cells, and it is only about an order of magnitude larger than the Landauer bound [35,36,37,90,91,92,93,94,95,96,97,98,99,100,107]. Thus, the Landauer principle briefly formulated as “information is physical” may be re-shaped to “biological information is physical and, strictly speaking, thermodynamically rooted”. Symmetry and ordering essentially increase the computational/informational/energetic effectivity of biological systems [33].
It should be emphasized that the role of symmetry in biological systems is very different from that in physics (such as symmetry of crystals). Symmetry in the biologically rooted patterns is never perfect. Thus, mathematical methods enabling the quantification of symmetry and its deviation from the mathematically perfect symmetrical patterns were suggested [61,62,63,64,65,66,67,68,69]. A number of mathematical measures (Procrustes, Hausdorff or Euclidian distances) may be used for quantification of deviation of a biological pattern from a perfect symmetry [115,116]. It turns out that the notion of “order” has a fine structure and may be quantified mathematically with essentially different mathematical measures such as the continuous measure of symmetry, Voronoi (Shannon) entropy and Minkowski functionals [61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81]. Thus, a unique mathematical measure quantifying “order” in biological systems does not exist. It was demonstrated that the maxima and minima of the Shannon entropy and continuous measure of symmetry are not necessarily correlated; moreover, in certain cases, the maxima of the continuous measure of symmetry may correspond to the minima of the Shannon entropy [78]. The concept of fluctuating asymmetry was introduced for quantification of deviation from perfect symmetry in biological systems [115,116]. Accurate quantification of ordering and symmetry in biological patterns is well expected to be a hot theme of the nearest future mathematical biology investigations. Consider that “physical order”, as quantified by the Boltzmann entropy, and symmetry are in an intimate relation: symmetry usually decreases the Boltzmann entropy of systems [10,11]. Correlation between the Boltzmann entropy [44,45] and various mathematical measures of order should be elucidated.
Moreover, breaking of symmetry and asymmetry are crucial for understanding biological phenomena [34,86,87]. For example, breaking symmetry is a crucial step in the development of all biological systems [34]. Thus, the Curie–Minnigerode–Neumann principle may reshaped for biological systems as follows: “asymmetry is what creates a biological phenomenon” [82,83]. Thus, we conclude that symmetry and order, abundant in biologically rooted patterns, could be hardly related to a single, unique physical or evolutionary reason. It also should be emphasized that the symmetry of biological objects evolved with time, as discussed in [116,117] for the bilateral symmetry of flowers and sea urchins (in conjunction with cephalization). We conclude that a general unifying theory of symmetry in biology does not exist and, perhaps, has no chance to be developed. However, the role of symmetry/asymmetry (in relation to informational reasoning) in understanding biological phenomena is crucial.
Funding
This research received no external funding.
Data Availability Statement
Not applicable.
Acknowledgments
The author is thankful to M. Nosonovsky for extremely fruitful discussions. The author is thankful to Irina Legchenkova for her kind help in preparing this paper. The author is thankful to the anonymous reviewer for the extremely fruitful review of the manuscript.
Conflicts of Interest
The author declares no conflict of interest.
References
- Finnerty, J.H. The origins of axial patterning in the metazoa: How old is bilateral symmetry? Int. J. Dev. Biol. 2003, 47, 523–529. [Google Scholar] [PubMed]
- Finnerty, J.H.; Pang, K.; Burton, P.; Paulson, D.; Martindale, M.Q. Origins of Bilateral Symmetry: Hox and Dpp Expression in a Sea Anemone. Science 2004, 304, 1335. [Google Scholar] [CrossRef] [PubMed]
- Longo, G.; Montévil, M. From Physics to Biology by Extending Criticality and Symmetry Breakings. In Perspectives on Organisms; Lecture Notes in Morphogenesis; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Yonekura, T.; Sugiyama, M. Symmetry and its transition in phyllotaxis. J. Plant. Res. 2021, 134, 417–430. [Google Scholar] [CrossRef] [PubMed]
- Dumais, J. Can mechanics control pattern formation in plants? Curr. Opin. Plant Biol. 2007, 10, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Dengler, N.G. Anisophylly and dorsiventral shoot symmetry. Int. J. Plant Sci. 1999, 160, S67–S80. [Google Scholar] [CrossRef]
- Cannon, K.A.; Ochoa, J.M.; Yeates, T.O. High-symmetry protein assemblies: Patterns and emerging applications. Curr. Opin. Struct. Biol. 2019, 55, 77–84. [Google Scholar] [CrossRef]
- Wolynes, P.G. Symmetry and the energy landscapes of biomolecules. Proc. Natl. Acad. Sci. USA 1996, 93, 14249–14255. [Google Scholar] [CrossRef]
- Hollo, G. A new paradigm for animal symmetry. Interface Focus 2015, 5, 20150032. [Google Scholar] [CrossRef]
- Bormashenko, E. Entropy, Information, and Symmetry: Ordered is Symmetrical. Entropy 2020, 22, 11. [Google Scholar] [CrossRef]
- Bormashenko, E. Entropy, Information, and Symmetry; Ordered is Symmetrical, II: System of Spins in the Magnetic Field. Entropy 2020, 22, 235. [Google Scholar] [CrossRef]
- Macia, E. The role of aperiodic order in science and technology. Rep. Prog. Phys. 2005, 68, 1–45. [Google Scholar] [CrossRef]
- Mitchison, G.J. Phyllotaxis and the Fibonacci series. Science 1977, 196, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Adam, J.A. Mathematics in Nature: Modeling Patterns in the Natural World; Princeton University Press: Princeton, NJ, USA, 2003; Chapter 10; pp. 213–230. [Google Scholar]
- Nowlan, R.A. Rabbits & Patterns. In Masters of Mathematics; SensePublishers: Rotterdam, The Netherlands, 2017; Chapter 10; pp. 161–177. [Google Scholar]
- Posamentier, A.S.; Lehmann, I. The Fabulous Fibonacci Numbers; Prometheus Books: Amherst, NY, USA, 2007; Chapter 2; pp. 59–76. [Google Scholar]
- Swinton, J.; Ochu, E. Novel Fibonacci and non-Fibonacci structure in the sunflower: Results of a citizen science experiment. R. Soc. Open Sci. 2016, 3, 160091. [Google Scholar] [CrossRef]
- Li, R.; Bowerman, B. Symmetry Breaking in Biology. Cold Spring Harb. Perspect. Biol. 2010, 2, a003475. [Google Scholar] [CrossRef] [PubMed]
- Hollo, G. Demystification of animal symmetry: Symmetry is a response to mechanical forces. Biol. Direct. 2017, 12, 11. [Google Scholar] [CrossRef] [PubMed]
- Holló, G.; Novák, M. The manoeuvrability hypothesis to explain the maintenance of bilateral symmetry in animal evolution. Biol Direct. 2012, 7, 22. [Google Scholar] [CrossRef]
- Manuel, M. Early evolution of symmetry and polarity in metazoan body plans. Comptes Rendus Biol. 2009, 332, 184–209. [Google Scholar] [CrossRef]
- Van Workum, K.; Douglas, J.F. Schematic Models of Molecular Self-Organization. Macromol. Symp. 2005, 227, 1–16. [Google Scholar] [CrossRef]
- Van Workum, K.; Douglas, J.F. Symmetry, equivalence, and molecular self-assembly. Phys. Rev. E 2006, 73, 031502. [Google Scholar] [CrossRef]
- Williams, K.S.; Smith, K.G.; Stephen, F.M. Emergence of 13-Yr Periodical Cicadas (Cicadidae: Magicicada): Phenology, Mortality, and Predators Satiation. Ecology 1993, 74, 1143–1152. [Google Scholar] [CrossRef]
- Yoshimura, J. The Evolutionary Origins of Periodical Cicadas During Ice Ages. Am. Nat. 1997, 149, 112–124. [Google Scholar] [CrossRef]
- Cox, R.; Carlton, C.E. Paleoclimatic Influences in the Evolution of Periodical Cicadas. Am. Midl. Nat. 1988, 120, 183–193. [Google Scholar] [CrossRef]
- Cox, R.; Carlton, C.E. A Commentary on Prime Numbers and Life Cycles of Periodical Cicadas. Am. Nat. 1998, 152, 162–164. [Google Scholar] [CrossRef] [PubMed]
- Webb, G.F. The prime number periodical cicada problem. Am. Inst. Math. Sci. 2001, 1, 387–399. [Google Scholar] [CrossRef]
- Goles, E.; Schulz, A.B.; Markus, M. Prime number selection of cycles in a predator-prey model. Complexity 2001, 6, 33–38. [Google Scholar] [CrossRef]
- Baker, A. Are there Genuine Mathematical Explanations of Physical Phenomena? Mind 2005, 114, 223–238. [Google Scholar] [CrossRef]
- Swaddle, J.H. Reproductive success and symmetry in zebra finches. Anim. Behav. 1996, 51, 203–210. [Google Scholar] [CrossRef]
- Hansen, L.T.T.; Amundsen, T.; Forsgren, E. Symmetry: Attractive not only to females. Proc. R. Soc. Lond. B 1999, 266, 1235–1240. [Google Scholar] [CrossRef]
- Johnston, I.G.; Dinglee, K.; Greenbury, S.F.; Camargoh, Q.; Doyei, J.P.K.; Ahnert, S.E.; Louis, A.A. Symmetry and simplicity pontaneously emerge from the algorithmic nature of evolution. Proc. Natl. Acad. Sci. USA 2022, 119, e2113883119. [Google Scholar] [CrossRef]
- Ocklenburg, S.; Mundorf, A. Symmetry and asymmetry in biological structures. Proc. Natl. Acad. Sci. USA 2022, 119, e2204881119. [Google Scholar] [CrossRef]
- Landauer, R. Dissipation and heat generation in the computing process. IBM J. Res. Dev. 1961, 5, 183. [Google Scholar] [CrossRef]
- Landauer, R. Information is physical. Phys. Today 1991, 44, 23–29. [Google Scholar] [CrossRef]
- Bormashenko, E. The Landauer Principle: Re-Formulation of the Second Thermodynamics Law or a Step to Great Unification? Entropy 2019, 21, 918. [Google Scholar] [CrossRef]
- Finnerty, J.H. Did internal transport, rather than directed locomotion, favor the evolution of bilateral symmetry in animals? Bioessays 2005, 27, 1174–1180. [Google Scholar] [CrossRef]
- Toxvaerd, S. The Emergence of the Bilateral Symmetry in Animals: A Review and a New Hypothesis. Symmetry 2021, 13, 261. [Google Scholar] [CrossRef]
- Weihs, D. Stability Versus Maneuverability in Aquatic Locomotion. Integr. Comp. Biol. 2002, 42, 127–134. [Google Scholar] [CrossRef]
- Lighthill, M.J. Aquatic animal propulsion of high hydromechanical efficiency. J. Fluid Mech. 1970, 44, 265–301. [Google Scholar] [CrossRef]
- Landau, L.D.; Lifshitz, E.M. Statistical Physics, 3rd ed.; Course of Theoretical Physics; Elsevier: Oxford, UK, 2011. [Google Scholar]
- Kittel, C.H. Thermal Physics; John and Wiley & Sons: New York, NY, USA, 1969. [Google Scholar]
- Baierlein, R. Thermal Physics; Cambridge University Press: Cambridge, UK, 1999. [Google Scholar]
- Shroeder, D.V. An Introduction to Thermal Physics; Oxford University Press: Oxford, UK, 2021. [Google Scholar]
- van Hemmen, J.L. A Generalized Equipartition Theorem. Phys. Lett. A 1980, 79, 25–28. [Google Scholar] [CrossRef]
- Lawrence, E.; Turner, L.E., Jr. Generalized Classical Equipartition Theorem. Am. J. Phys. 1976, 44, 104–105. [Google Scholar]
- Landsberg, P.T. Generalized Equipartition. Am. J. Phys. 1978, 46, 296. [Google Scholar] [CrossRef]
- Landsberg, P.T. Equipartition for a Relativistic Gas. Am. J. Phys. 1992, 60, 561. [Google Scholar] [CrossRef]
- Lawless, W.N. Energy Equipartition: A Restatement. Am. J. Phys. 1964, 32, 686–687. [Google Scholar] [CrossRef]
- Bormashenko, E.; Gendelman, O. On the applicability of the equipartition theorem. Therm. Sci. 2010, 14, 855–858. [Google Scholar] [CrossRef]
- Stephen, M.J.; Straley, J.P. Physics of liquid crystals. Rev. Mod. Phys. 1974, 46, 617. [Google Scholar] [CrossRef]
- Mathur-De Vré, R. The NMR studies of water in biological systems. Prog. Biophys. Mol. Biol. 1980, 35, 103–134. [Google Scholar] [CrossRef]
- Tait, M.; Franks, F. Water in Biological Systems. Nature 1971, 230, 91–94. [Google Scholar] [CrossRef]
- Tortora, L.; Lavrentovich, O.D. Chiral symmetry breakingby spatial confinement in tactoidal droplets of lyotropic chromonic liquid rystals. Proc. Natl. Acad. Sci. USA 2011, 108, 5163–5168. [Google Scholar] [CrossRef]
- Strzelecka, T.E.; Davidson, M.W.; Rill, R.L. Multiple liquid crystal phases of DNA at high concentrations. Nature 1988, 331, 457–460. [Google Scholar] [CrossRef]
- Dierking, I. Chiral Liquid Crystals: Structures, Phases, Effects. Symmetry 2014, 6, 444–472. [Google Scholar] [CrossRef]
- Dierking, I.; Figueiredo Neto, A.M. Novel Trends in Lyotropic Liquid Crystals. Crystals 2020, 10, 604. [Google Scholar] [CrossRef]
- Bawden, F.C.; Pirie, N.W.; Bernal, J.D.; Fankuchen, I. Liquid Crystalline Substances from Virus-infected Plants. Nature 1936, 138, 1051–1052. [Google Scholar] [CrossRef]
- Zabrodsky, H.; Peleg, S.; Avnir, D. Continuous symmetry measures. J. Am. Chem. Soc. 1992, 114, 7843–7851. [Google Scholar] [CrossRef]
- Zabrodsky, H.; Peleg, S.; Avnir, D. Continuous symmetry measures. 2. Symmetry groups and the tetrahedron. J. Am. Chem. Soc. 1993, 115, 8278–8828. [Google Scholar] [CrossRef]
- Zabrodsky, H.; Avnir, D. Continuous Symmetry Measures. 4. Chirality. J. Am. Chem. Soc. 1995, 117, 462–473. [Google Scholar] [CrossRef]
- Pinsky, M.; Avnir, D. Continuous Symmetry Measures. 5. The Classical Polyhedra. Inorg. Chem. 1998, 37, 5575–5582. [Google Scholar] [CrossRef] [PubMed]
- Zabrodsky, H.; Peleg, S.; Avnir, D. Symmetry as a continuous feature. IEEE Trans. Pattern Anal. Mach. Intell. 1995, 17, 1154–1166. [Google Scholar] [CrossRef]
- Grimme, S. Continuous symmetry measures for electronic wavefunctions. Chem. Phys. Lett. 1998, 297, 15–22. [Google Scholar] [CrossRef]
- Bormashenko, E.; Legchenkova, I.; Frenkel, M.; Shvalb, N.; Shoval, S. Voronoi Entropy vs. Continuous Measure of Symmetry of the Penrose Tiling: Part I. Analysis of the Voronoi Diagrams. Symmetry 2021, 13, 1659. [Google Scholar] [CrossRef]
- Pinsky, M.; Zait, A.; Bonjack, M.; Avnir, D. Continuous symmetry analyses: Cnv and Dn measures of molecules, complexes, and proteins. Comp. Chem. 2013, 34, 2–9. [Google Scholar] [CrossRef]
- Bonjack, M.; Avni, D. The near-symmetry of protein oligomers: NMR-derived structures. Sci. Rep. 2020, 10, 8367. [Google Scholar] [CrossRef]
- Voronoi, G. Nouvelles applications des paramètres continus à la théorie des formes quadratiques. Deuxième mémoire. Recherches sur les paralléloèdres primitifs. Reine Angew. Math. 1908, 134, 198–287. [Google Scholar] [CrossRef]
- Liebling, T.M.; Pournin, L. Voronoi diagrams and Delaunay triangulations: Ubiquitous Siamese Twins. Doc. Math. ISMP 2012, 419, 431. [Google Scholar]
- Suárez-Plasencia, L.; Herrera-Macías, J.A.; Legón-Pérez, C.M.; Socorro-LLanes, R.; Rojas, O.; Sosa-Gómez, G. Analysis of the Number of Sides of Voronoi Polygons in PassPoint. In Computer Science and Health Engineering in Health Services. COMPSE 2020; Marmolejo-Saucedo, J.A., Vasant, P., Litvinchev, I., Rodriguez-Aguilar, R., Martinez-Rios, F., Eds.; Lecture Notes of the Institute for Computer Sciences, Social Informatics and Telecommunications Engineering; Springer: Cham, Switzerland, 2021; Volume 359. [Google Scholar]
- Poupon, A. Voronoi and Voronoi-related tessellations in studies of protein structure and interaction. Curr. Opin. Struct. Biol. 2004, 14, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Bock, M.; Tyagi, A.K.; Kreft, J.U.; Alt, W. Generalized Voronoi Tessellation as a Model of Two-dimensional Cell Tissue Dynamics. Bull. Math. Biol. 2010, 72, 1696–1731. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.T.; Tao, C.L.; Zhang, X. Mesophasic organization of GABAA receptors in hippocampal inhibitory synapses. Nat. Neurosci. 2020, 23, 1589–1596. [Google Scholar] [CrossRef]
- Railam, T.; Ambrulevičius, F.; Penkauskas, T.; Jankune, M.; Meškauskasm, T.; Vanderah, T.; Valincius, G. Clusters of protein pores in phospholipid bilayer membranes can be identified and characterized by electrochemical impedance spectroscopy. Electrochim. Acta 2020, 364, 137179. [Google Scholar] [CrossRef]
- Bormashenko, E.; Frenkel, M.; Vilk, A.; Legchenkova, I.; Fedorets, A.A.; Aktaev, N.; Dombrovsky, L.A.; Nosonovsky, M. Characterization of Self-Assembled 2D Patterns with Voronoi Entropy. Entropy 2018, 20, 956. [Google Scholar] [CrossRef]
- Frenkel, M.; Fedorets, A.A.; Dombrovsky, L.A.; Nosonovsky, M.; Legchenkova, I.; Bormashenko, E. Continuous Symmetry Measure vs Voronoi Entropy of Droplet Clusters. J. Phys. Chem. C 2021, 125, 2431–2436. [Google Scholar] [CrossRef]
- Larkin, T.J.; Canuto, H.C.; Kettunen, M.I.; Booth, T.S.; Hu, D.; Krishnan, A.S.; Bohndiek, S.E.; Neves, A.A.; McLachlan, C.; Hobson, M.P.; et al. Analysis of image heterogeneity using 2D Minkowski functionals detects tumor responses to treatment. Magn. Reson. Med. 2014, 71, 402–410. [Google Scholar] [CrossRef]
- Boehm, H.F.; Link, T.M.; Monetti, R.; Mueller, D.; Rummeny, E.J.; Newitt, D.; Majumdar, S.; Raeth, C. Application of the Minkowski functionals in 3D to high-resolution MR images of trabecular bone: Prediction of the biomechanical strength by nonlinear topological measures. Proc. SPIE 2004, 5370, 172–180. [Google Scholar]
- Bormashenko, E.; Legchenkova, I.; Frenkel, F.; Shvalb, N.; Shoval, S. Shannon (Information) Measures of Symmetry for 1D and 2D Shapes and Patterns. Appl. Sci. 2022, 12, 1127. [Google Scholar] [CrossRef]
- Curie, P. Sur la symetrie. Soc. Mineralog. France Bull. Paris 1884, 7, 418–457. [Google Scholar] [CrossRef]
- Brandmuller, J. An extension of the Neumann-Minnigerode-Curie Principle. Comp. Maths. Appl. 1986, 12, 97–100. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, J.; Yu, J.; Guo, Y.; Sun, Q. Symmetry-breaking induced large piezoelectricity in Janus tellurene materials. Phys. Chem. Chem. Phys. 2019, 21, 1207–1216. [Google Scholar] [CrossRef] [PubMed]
- Aizenberg, J.; Hanson, J.; Koetzle, T.F.; Leiserowitz, L.; Weiner, S.; Addadi, L. Biologically Induced Reduction in Symmetry: A Study of Crystal Texture of Calcitic Sponge Spicules. Chem. Eur. J. 1995, 1, 414–422. [Google Scholar] [CrossRef]
- Dingle, K.; Pérez, G.V.; Louis, A.A. Generic predictions of output probability based on complexities of inputs and outputs. Sci. Rep. 2020, 10, 4415. [Google Scholar] [CrossRef]
- Kong, X.-Z.; Mathias, S.R.; Guadalupe, T.; ENIGMA Laterality Working Group; Glahn, D.C.; Franke, B.; Crivello, F.; Tzourio-Mazoyer, N.; Fisher, S.E.; Thompson, P.M.; et al. Mapping cortical brain asymmetry in 17,141 healthy individuals worldwide via the ENIGMA Consortium. Proc. Natl. Acad. Sci. USA 2018, 115, E5154–E5163. [Google Scholar] [CrossRef]
- Boman, B.M.; Dinh, T.N.; Decker, K.; Emerick, B.; Raymond, C.; Schleiniger, G. Why do Fibonacci numbers appear in patterns of growth in nature? A model for tissue renewal based on asymmetric cell division. Fibonacci Q. 2017, 55, 30–41. [Google Scholar]
- Wheeler, J.A. Information, physics, quantum: The search for links. In Proceedings of the III International Symposium on Foundations of Quantum Mechanics, Tokyo, Japan, 28–31 August 1989. [Google Scholar]
- Bormashenko, E. Informational Reinterpretation of the Mechanics Notions and Laws. Entropy 2020, 22, 631. [Google Scholar] [CrossRef]
- Vopson, M.M. The mass-energy-information equivalence principle. AIP Adv. 2019, 9, 095206. [Google Scholar] [CrossRef]
- Vopson, M.M. Experimental protocol for testing the mass–energy–information equivalence principle. AIP Adv. 2022, 12, 035311. [Google Scholar] [CrossRef]
- Vopson, M.M. The information catastrophe. AIP Adv. 2020, 10, 085014. [Google Scholar] [CrossRef]
- Bérut, A.; Arakelyan, A.; Petrosyan, A.; Ciliberto, S.; Dillenschneider, R.; Lutz, E. Experimental verification of Landauer’s principle linking information and thermodynamics. Nature 2012, 483, 187–189. [Google Scholar] [CrossRef]
- Jun, Y.; Gavrilov, Y.M.; Bechhoefer, J. High-precision test of Landauer’s principle in a feedback trap. Phys. Rev. Lett. 2014, 113, 190601. [Google Scholar] [CrossRef] [PubMed]
- Aydin, A.; Sisman, A.; Kosloff, R. Landauer’s Principle in a Quantum Szilard Engine without Maxwell’s Demon. Entropy 2020, 22, 294. [Google Scholar] [CrossRef] [PubMed]
- Herrera, L. Landauer Principle and General Relativity. Entropy 2020, 22, 340. [Google Scholar] [CrossRef]
- Müller, J.G. Observable and Unobservable Mechanical Motion. Entropy 2020, 22, 737. [Google Scholar] [CrossRef]
- Müller, J.G. Photon Detection as a Process of Information Gain. Entropy 2020, 22, 392. [Google Scholar] [CrossRef]
- Müller, J.G. Information Contained in Molecular Motion. Entropy 2019, 21, 1052. [Google Scholar] [CrossRef]
- Yolles, M.; Frieden, R. Viruses as Living Systems—A Metacybernetic View. Systems 2022, 10, 70. [Google Scholar] [CrossRef]
- Vopson, M.M. A Possible Information Entropic Law of Genetic Mutations. Appl. Sci. 2022, 12, 6912. [Google Scholar] [CrossRef]
- Bormashenko, E.; Voronel, A. Spatial scales of living cells and their energetic and informational capacity. Eur. Biophys. J. 2018, 47, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Vellai, T.; Vida, G. The origin of eukaryotes: The difference between prokaryotic and eukaryotic cells. Proc. R. Soc. B 1999, 266, 1571–1577. [Google Scholar] [CrossRef] [PubMed]
- Lane, N.; Martin, W. The energetics of genome complexity. Nature 2010, 467, 929–934. [Google Scholar] [CrossRef]
- Goldman, N.; Bertone, P.; Chen, A.; Dessimoz, C.; LeProust, E.M.; Sipos, B.; Birney, B. Towards practical, high-capacity, low-maintenance information storage in synthesized DNA. Nature 2013, 494, 77–80. [Google Scholar] [CrossRef]
- Kempes, C.P.; Wolpert, D.; Cohen, Z.; Pérez-Mercader, J. The thermodynamic efficiency of computations made in cells across the range of life. Philos. Trans. R. Soc. A 2017, 375, 20160343. [Google Scholar] [CrossRef]
- Gillings, M.R.; Hilbert, M.; Kemp, D.J. Information in the Biosphere: Biological and Digital Worlds. Trends Biol. Evol. 2016, 31, 180–189. [Google Scholar] [CrossRef]
- Schrödinger, E. What is Life? The Physical Aspects of the Living Cell; Cambridge University Press: New York, NY, USA, 1944. [Google Scholar]
- Weyl, H. Symmetry; Princeton University Press: Princeton, NJ, USA, 1989. [Google Scholar]
- Homblé, F.; Léonetti, M. Emergence of symmetry breaking in fucoid zygotes. Trends Plant. Sci. 2007, 12, 253–259. [Google Scholar] [CrossRef]
- Kaandorp, J.A.; de Kluijver, M.J. Verification of fractal growth models of the sponge Haliclona oculata (Porifera) with transplantation experiments. Mar. Biol. 1992, 113, 133–143. [Google Scholar] [CrossRef]
- Abrams, M.J.; Basinger, T.; Yuan, W.; Guo, C.-L.; Goentoro, L. Self-repairing symmetry in jellyfish through mechanically driven reorganization. Proc. Natl. Acad. Sci. USA 2015, 112, E3365–E3373. [Google Scholar] [CrossRef]
- Duarte, J.M.; Dutta, S.; Goodsell, D.S.; Burley, S.K. Exploring protein symmetry at the RCSB Protein Data Bank. Emerg. Top Life Sci. 2022, 6, 231–243. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.H.; Raz, S.; Hel-Or, H.; Nevo, E. Fluctuating Asymmetry: Methods, Theory, and Applications. Symmetry 2010, 2, 466–540. [Google Scholar] [CrossRef]
- Klingenberg, C.P. Analyzing Fluctuating Asymmetry with Geometric Morphometrics: Concepts, Methods, and Applications. Symmetry 2015, 7, 843–934. [Google Scholar] [CrossRef]
- Hileman, L.C. Bilateral flower symmetry—How, when and why? Curr. Opin. Plant Biol. 2014, 17, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Grabowsky, G.L. Symmetry, locomotion and the evolution of an anterior end: A lesson from sea urchins. Evolution 1994, 48, 1131–1146. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).