
SARS-CoV-2 Spike-Derived Peptides Presented by HLA Molecules

 ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Protein Expression, Refold, and Purification
2.2. Differential Scanning Fluorimetry
2.3. Peptide Conservation within SARS-CoV-2 Isolates
2.4. Crystallisation and Structural Determination
3. Results
3.1. The Spike-Derived Peptides Were Able to Form a Stable Complex with Their Respective HLA Molecules
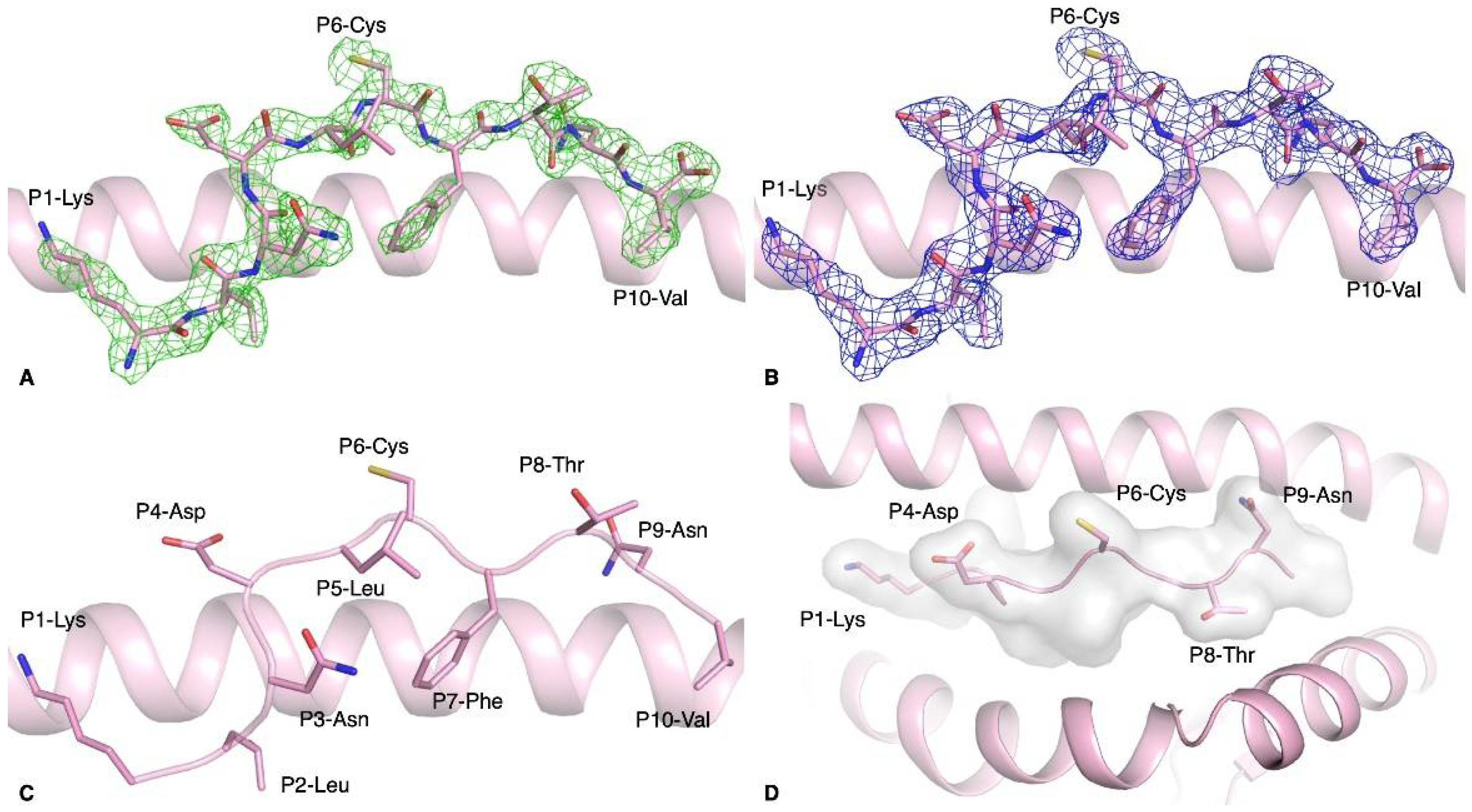
3.2. Structure of HLA-A*02:01-S386–395 Reveals a Flat Peptide Conformation
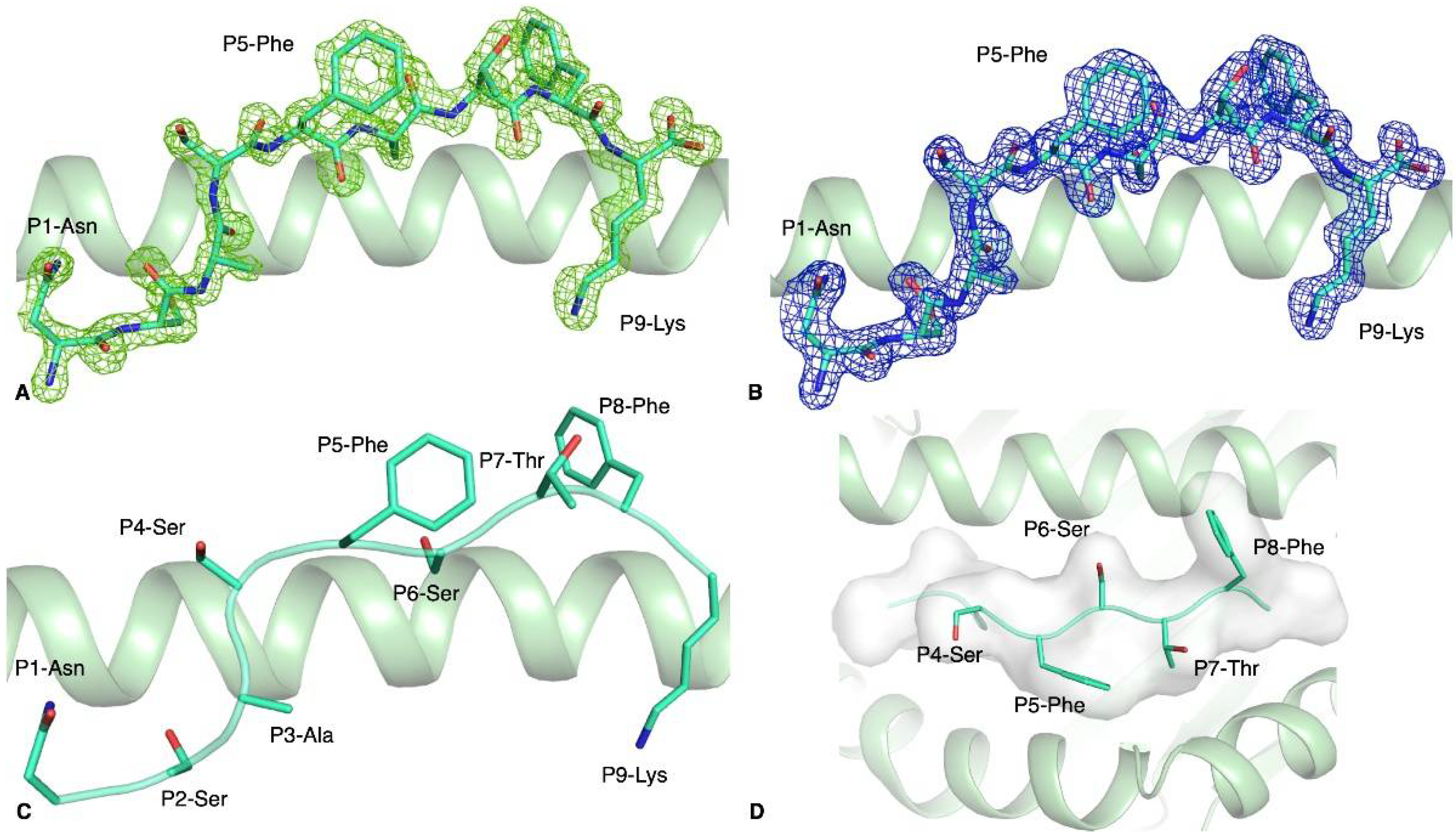
3.3. HLA-A*11:01-S370–378 Presents Aromatic Residues to T Cells
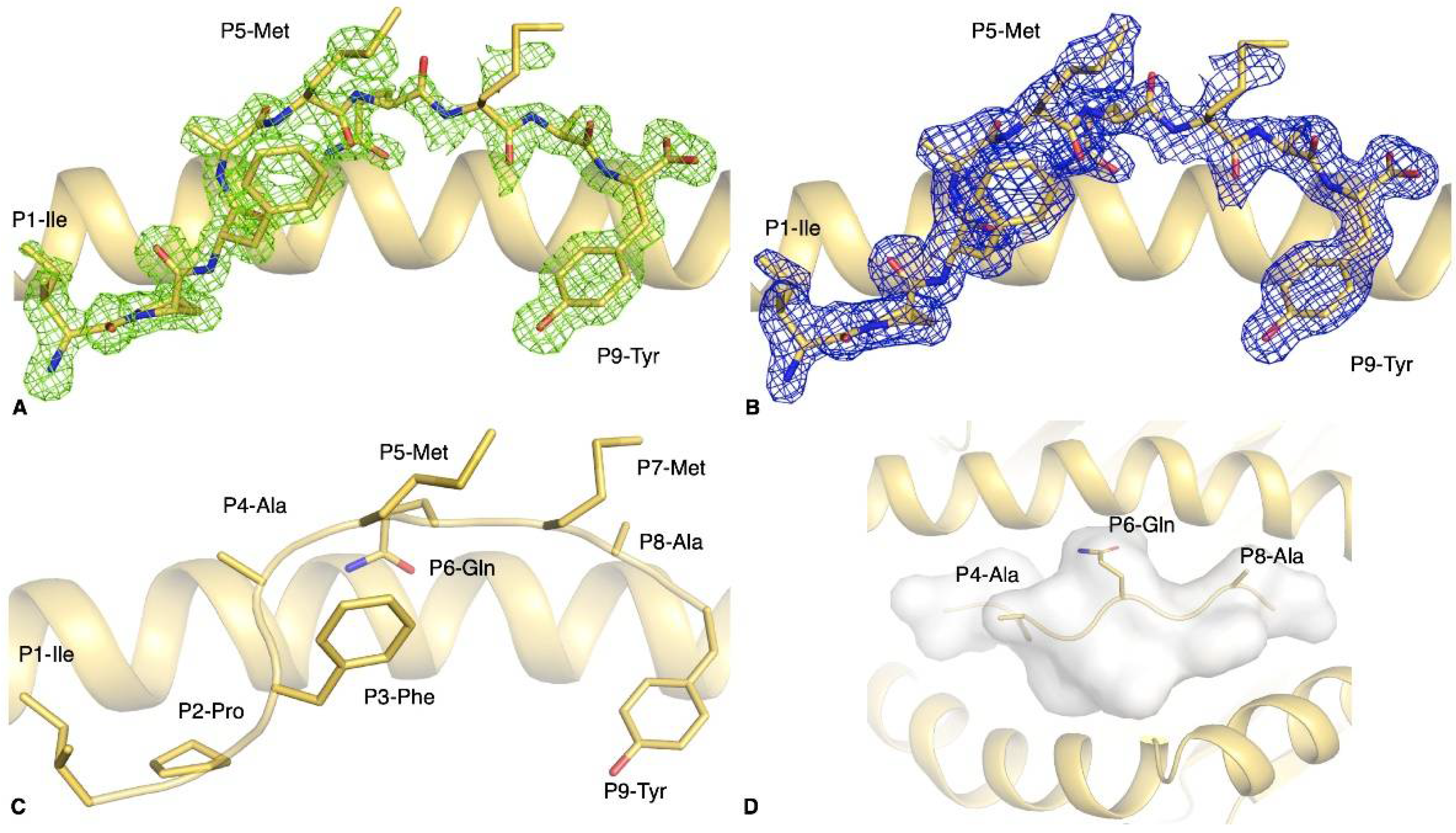
3.4. The S896–904 Peptide Adopted a Flat Conformation in the Cleft of HLA-B*35:01
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zuo, J.; Dowell, A.C.; Pearce, H.; Verma, K.; Long, H.M.; Begum, J.; Aiano, F.; Amin-Chowdhury, Z.; Hallis, B.; Stapley, L.; et al. Robust SARS-CoV-2-specific T cell immunity is maintained at 6 months following pxrimary infection. Nat. Immunol. 2021, 22, 620–626. [Google Scholar] [CrossRef]
- Poh, C.M.; Carissimo, G.; Wang, B.; Amrun, S.N.; Lee, C.Y.; Chee, R.S.; Fong, S.-W.; Yeo, N.K.-W.; Lee, W.-H.; Torres-Ruesta, A. Two linear epitopes on the SARS-CoV-2 spike protein that elicit neutralising antibodies in COVID-19 patients. Nat. Commun. 2020, 11, 2806. [Google Scholar] [CrossRef]
- Shomuradova, A.S.; Vagida, M.S.; Sheetikov, S.A.; Zornikova, K.V.; Kiryukhin, D.; Titov, A.; Peshkova, I.O.; Khmelevskaya, A.; Dianov, D.V.; Malasheva, M. SARS-CoV-2 Epitopes Are Recognized by a Public and Diverse Repertoire of Human T Cell Receptors. Immunity 2020, 53, 1245. [Google Scholar] [CrossRef] [PubMed]
- Canete, P.F.; Vinuesa, C.G. COVID-19 Makes B Cells Forget, but T Cells Remember. Cell 2020, 183, 13. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, F.R.; McLaren, J.E. T Cell Immunity to Bacterial Pathogens: Mechanisms of Immune Control and Bacterial Evasion. Int. J. Mol. Sci. 2020, 21, 6144. [Google Scholar] [CrossRef] [PubMed]
- Szeto, C.; Lobos, C.A.; Nguyen, A.T.; Gras, S. TCR Recognition of Peptide-MHC-I: Rule Makers and Breakers. Int. J. Mol. Sci. 2020, 22, 68. [Google Scholar] [CrossRef]
- Szeto, C.; Chatzileontiadou, D.S.M.; Nguyen, A.T.; Sloane, H.; Lobos, C.A.; Jayasinghe, D.; Halim, H.; Smith, C.; Riboldi-Tunnicliffe, A.; Grant, E.J. The Presentation of SARS-CoV-2 Peptides by the Common HLA-A*02:01 Molecule. iScience 2020, 24, 102096. [Google Scholar] [CrossRef]
- Jarjour, N.N.; Jameson, S.C. T Cell Memory: Understanding COVID-19. Immunity 2020, 54, 14–18. [Google Scholar] [CrossRef]
- Koutsakos, M.; Illing, P.T.; Nguyen, T.H.O.; Mifsud, N.A.; Crawford, J.C.; Rizzetto, S.; Eltahla, A.A.; Clemens, E.B.; Sant, S.; Chua, B.Y. Human CD8(+) T cell cross-reactivity across influenza A, B and C viruses. Nat. Immunol. 2019, 20, 613. [Google Scholar] [CrossRef]
- Agerer, B.; Koblischke, M.; Gudipati, V.; Montano-Gutierrez, L.F.; Smyth, M.; Popa, A.; Genger, J.-W.; Endler, L.; Florian, D.M.; Mühlgrabner, V.; et al. SARS-CoV-2 mutations in MHC-I-restricted epitopes evade CD8(+) T cell responses. Sci. Immunol. 2021, 6, 57. [Google Scholar] [CrossRef] [PubMed]
- Aragão, D.; Aishima, J.; Cherukuvada, H.; Clarken, R.; Clift, M.; Cowieson, N.P.; Ericsson, D.J.; Gee, C.L.; Macedo, S.; Mudie, N. MX2: A high-flux undulator microfocus beamline serving both the chemical and macromolecular crystallography communities at the Australian Synchrotron. J. Synchrotron Radiat. 2018, 25 Pt 3, 885. [Google Scholar] [CrossRef]
- Kabsch, W. XDS. Acta Crystallogr. D Biol. Crystallogr. 2010, 66 Pt 2, 125. [Google Scholar] [CrossRef]
- McCoy, A.J.; Grosse-Kunstleve, R.W.; Adams, P.D.; Winn, M.D.; Storoni, L.C.; Read, R.J. Phaser crystallographic software. J. Appl. Crystallogr. 2007, 40 Pt 4, 658. [Google Scholar] [CrossRef]
- Collaborative Computational Project, Number 4. The CCP4 suite: Programs for protein crystallography. Acta Crystallogr. D Biol. Crystallogr. 1994, 50 Pt 5, 760. [Google Scholar]
- Culshaw, A.; Ladell, K.; Gras, S.; E McLaren, J.; Miners, K.L.; Farenc, C.; Heuvel, H.V.D.; Gostick, E.; Dejnirattisai, W.; Wangteeraprasert, A. Germline bias dictates cross-serotype reactivity in a common dengue-virus-specific CD8(+) T cell response. Nat. Immunol. 2017, 18, 1228. [Google Scholar] [CrossRef] [PubMed]
- Gras, S.; Kedzierski, L.; Valkenburg, S.A.; Laurie, K.; Liu, Y.C.; Denholm, J.T.; Richards, M.J.; Rimmelzwaan, G.F.; Kelso, A.; Doherty, P.C. Cross-reactive CD8+ T-cell immunity between the pandemic H1N1-2009 and H1N1-1918 influenza A viruses. Proc. Natl. Acad. Sci. USA 2010, 107, 12599. [Google Scholar] [CrossRef] [PubMed]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Crystallogr. D Biol. Crystallogr. 2010, 66 Pt 4, 486. [Google Scholar] [CrossRef]
- Bricogne, G.B.E.; Brandl, M.; Flensburg, C.; Keller, P.; Paciorek, W.; Roversi, P.S.A.; Smart, O.S.; Vonrhein, C.; Womack, T.O. Buster Version 2.10; Global Phasing Ltd.: Cambridge, UK, 2011. [Google Scholar]
- Can, H.; Koseoglu, A.E.; Erkunt Alak, S.; Guvendi, M.; Doskaya, M.; Karakavuk, M.; Gürüz, A.Y.; Ün, C. In silico discovery of antigenic proteins and epitopes of SARS-CoV-2 for the development of a vaccine or a diagnostic approach for COVID-19. Sci. Rep. 2020, 10, 22387. [Google Scholar] [CrossRef] [PubMed]
- Cun, Y.; Li, C.; Shi, L.; Sun, M.; Dai, S.; Sun, L.; Yao, Y. COVID-19 coronavirus vaccine T cell epitope prediction analysis based on distributions of HLA class I loci (HLA-A, -B, -C) across global populations. Hum. Vaccin. Immunother. 2020, 17, 1097–1108. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Kumar, P.; Saumya, K.U.; Kapuganti, S.K.; Bhardwaj, T.; Giri, R. Exploring the SARS-CoV-2 structural proteins for multi-epitope vaccine development: An in-silico approach. Expert Rev. Vaccines 2020, 19, 887. [Google Scholar] [CrossRef] [PubMed]
- Poran, A.; Harjanto, D.; Malloy, M.; Arieta, C.M.; Rothenberg, D.A.; Lenkala, D.; van Buuren, M.M.; Addona, T.A.; Rooney, M.S.; Srinivasan, L. Sequence-based prediction of SARS-CoV-2 vaccine targets using a mass spectrometry-based bioinformatics predictor identifies immunogenic T cell epitopes. Genome Med. 2020, 12, 70. [Google Scholar] [CrossRef] [PubMed]
- Tarke, A.; Sidney, J.; Kidd, C.K.; Dan, J.M.; Ramirez, S.I.; Yu, E.D.; Mateus, J.; da Silva Antunes, R.; Moore, E.; Rubiro, P. Comprehensive analysis of T cell immunodominance and immunoprevalence of SARS-CoV-2 epitopes in COVID-19 cases. Cell Rep. Med. 2021, 2, 100204. [Google Scholar] [CrossRef]
- Fiorentini, S.; Messali, S.; Zani, A.; Caccuri, F.; Giovanetti, M.; Ciccozzi, M.; Caruso, A. First detection of SARS-CoV-2 spike protein N501 mutation in Italy in August, 2020. Lancet Infect Dis. 2021. [Google Scholar] [CrossRef]
- Guan, P.; Doytchinova, I.A.; Flower, D.R. HLA-A3 supermotif defined by quantitative structure-activity relationship analysis. Protein Eng. 2003, 16, 11. [Google Scholar] [CrossRef] [PubMed]
- Grant, E.J.; Josephs, T.M.; Loh, L.; Clemens, E.B.; Sant, S.; Bharadwaj, M.; Chen, W.; Rossjohn, J.; Gras, S.; Kedzierska, K. Broad CD8(+) T cell cross-recognition of distinct influenza A strains in humans. Nat. Commun. 2018, 9, 5427. [Google Scholar] [CrossRef]
- Josephs, T.M.; Grant, E.J.; Gras, S. Molecular challenges imposed by MHC-I restricted long epitopes on T cell immunity. Biol. Chem. 2017, 398, 1027. [Google Scholar] [CrossRef] [PubMed]
- Hansasuta, P.; Dong, T.; Thananchai, H.; Weekes, M.; Willberg, C.; Aldemir, H.; Rowland-Jones, S.; Braud, V.M. Recognition of HLA-A3 and HLA-A11 by KIR3DL2 is peptide-specific. Eur. J. Immunol. 2004, 34, 1673. [Google Scholar] [CrossRef] [PubMed]
- Gupta, E.M.R.K.; Kumar Niraj, R.R. Identification of potential vaccine candidates against SARS-CoV-2, A step forward to fight COVID-19: A Reverse Vaccinology Approach. bioRxiv 2020. [Google Scholar] [CrossRef]
- Kared, H.; Redd, A.D.; Bloch, E.M.; Bonny, T.S.; Sumatoh, H.; Kairi, F.; Carbajo, D.; Abel, B.; Newell, E.W.; Bettinotti, M.P. SARS-CoV-2-specific CD8+ T cell responses in convalescent COVID-19 individuals. J. Clin. Investig. 2021, 131, 5. [Google Scholar] [CrossRef]
- Snyder, T.M.; Gittelman, R.M.; Klinger, M.; May, D.H.; Osborne, E.J.; Taniguchi, R.; Zahid, H.J.; Kaplan, I.M.; Dines, J.N.; Noakes, N.T.; et al. Magnitude and Dynamics of the T-Cell Response to SARS-CoV-2 Infection at Both Individual and Population Levels. medRxiv 2020. [Google Scholar] [CrossRef]
- Lineburg, K.E.; Grant, E.J.; Swaminathan, S.; Chatzileontiadou, D.S.; Szeto, C.; Sloane, H.; Panikkar, A.; Raju, J.; Crooks, P.; Rehan, S. CD8 + T cells specific for an immunodominant SARS-CoV-2 nucleocapsid epitope cross-react with selective seasonal coronaviruses. Immunity 2021, 54, 1055–1065. [Google Scholar] [CrossRef] [PubMed]
- Sekine, T.; Perez-Potti, A.; Rivera-Ballesteros, O.; Strålin, K.; Gorin, J.-B.; Olsson, A.; Llewellyn-Lacey, S.; Kamal, H.; Bogdanovic, G.; Muschiol, S. Robust T Cell Immunity in Convalescent Individuals with Asymptomatic or Mild COVID-19. Cell 2020, 183, 158. [Google Scholar] [CrossRef]
- Peng, Y.; Mentzer, A.J.; Liu, G.; Yao, X.; Yin, Z.; Dong, D.; Dejnirattisai, W.; Rostron, T.; Supasa, P.; Liu, C. Broad and strong memory CD4(+) and CD8(+) T cells induced by SARS-CoV-2 in UK convalescent individuals following COVID-19. Nat. Immunol. 2020, 21, 1336. [Google Scholar] [CrossRef] [PubMed]
- Lineburg, K.E.; Srihari, S.; Altaf, M.; Swaminathan, S.; Panikkar, A.; Raju, J.; Crooks, P.; Ambalathingal, G.R.; Martins, J.P.; Matthews, K.K. Rapid detection of SARS-CoV-2-specific memory T-cell immunity in recovered COVID-19 cases. Clin. Transl. Immunol. 2020, 9, e1219. [Google Scholar] [CrossRef]
- Le Bert, N.; Tan, A.T.; Kunasegaran, K.; Tham, C.Y.L.; Hafezi, M.; Chia, A.; Chng, M.H.Y.; Lin, M.; Tan, N.; Linster, M. SARS-CoV-2-specific T cell immunity in cases of COVID-19 and SARS, and uninfected controls. Nature 2020, 584, 457. [Google Scholar] [CrossRef]
- Wu, G.; Yan, S. Reasoning of spike glycoproteins being more vulnerable to mutations among 158 coronavirus proteins from different species. J. Mol. Model. 2005, 11, 8. [Google Scholar] [CrossRef]
- Korber, B.; Fischer, W.M.; Gnanakaran, S.; Yoon, H.; Theiler, J.; Abfalterer, W.; Hengartner, N.; Giorgi, E.E.; Bhattacharya, T.; Foley, B. Tracking Changes in SARS-CoV-2 Spike: Evidence that D614G Increases Infectivity of the COVID-19 Virus. Cell 2020, 182, 812. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Jackson, C.B.; Mou, H.; Ojha, A.; Peng, H.; Quinlan, B.D.; Rangarajan, E.S.; Pan, A.; Vanderheiden, A.; Suthar, M.S. SARS-CoV-2 spike-protein D614G mutation increases virion spike density and infectivity. Nat. Commun. 2020, 11, 6013. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| pHLA Complex | Peptide Sequence | Tm 1 (°C) |
|---|---|---|
| HLA-A*02:01-S386–395 | KLNDLCFTNV | 54.4 ± 1.3 |
| HLA-A*11:01-S370–378 | NSASFSTFK | 54.2 ± 0.2 |
| HLA-B*35:01-S896–904 | IPFAMQMAY | 61.6 ± 0.6 |
| Peptide | Africa | Asia | Europe | North America | Oceania | South America |
|---|---|---|---|---|---|---|
| S386–395 | 100% (770) | 100% (2195) | 99.89% (888) | 99.99% (49,376) | 100% (9919) | 100% (449) |
| KLNDLCFTNV | ||||||
| S370–378 | 99.87% (773) | 100% (2198) | 99.78% (889) | 99.89% (49,369) | 100% (9919) | 100% (441) |
| NSASFSTFK | ||||||
| S896–904 | 100% (774) | 99.37% (2208) | 100% (889) | 99.93% (49,386) | 99.90% (9919) | 99.78% (451) |
| IPFAMQMAY |
| Data Collection Statistics | HLA-A*02:01-S386–395 | HLA-A*11:01-S370–378 | HLA-B*35:01-S896–904 |
|---|---|---|---|
| Space group | P212121 | P21 | P212121 |
| Cell dimensions (a,b,c) (Å) | 64.60, 86.69, 163.21 | 49.87, 38.45, 110.06 | 51.24, 82.38, 111.01 |
| β = 94.63° | |||
| Resolution (Å) | 46.08–2.35 (2.43–2.35) | 46.72–1.50 (1.53–1.50) | 46.52–1.44 (1.47–1.44) |
| Total number of observations | 266,968 (25,197) | 458,019 (22,980) | 1,114,777 (47,083) |
| Number of unique observations | 39,044 (3769) | 67,146 (3290) | 84,730 (3897) |
| Multiplicity | 6.8 (6.7) | 6.8 (7.0) | 1.2 (12.1) |
| Data completeness (%) | 100 (100) | 100 (100) | 99.7 (93.6) |
| I/σI | 10.0 (2.0) | 13.8 (1.8) | 16.4 (1.8) |
| Rpim a (%) | 5.2 (40.9) | 2.9 (43.2) | 1.9 (40.5) |
| CC1/2 (%) | 98.2 (67.0) | 99.9 (75.4) | 99.9 (76.4) |
| Refinement Statistics | |||
| Rfactorb (%) | 18.1 | 18.1 | 20.4 |
| Rfreeb (%) | 23.0 | 21.1 | 22.9 |
| rmsd from ideality | |||
| Bond lengths (Å) | 0.010 | 0.010 | 0.010 |
| Bond angles (°) | 0.127 | 0.103 | 0.103 |
| Ramachandran plot (%) | |||
| Favoured | 96.0 | 99.0 | 99.0 |
| Allowed | 4.0 | 1.0 | 1.0 |
| Disallowed | 0.0 | 0.0 | 0.0 |
| PBD code | 7M8S | 7M8T | 7M8U |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, A.T.; Szeto, C.; Jayasinghe, D.; Lobos, C.A.; Halim, H.; Chatzileontiadou, D.S.M.; Grant, E.J.; Gras, S. SARS-CoV-2 Spike-Derived Peptides Presented by HLA Molecules. Biophysica 2021, 1, 194-203. https://doi.org/10.3390/biophysica1020015
Nguyen AT, Szeto C, Jayasinghe D, Lobos CA, Halim H, Chatzileontiadou DSM, Grant EJ, Gras S. SARS-CoV-2 Spike-Derived Peptides Presented by HLA Molecules. Biophysica. 2021; 1(2):194-203. https://doi.org/10.3390/biophysica1020015
Chicago/Turabian StyleNguyen, Andrea T., Christopher Szeto, Dhilshan Jayasinghe, Christian A. Lobos, Hanim Halim, Demetra S. M. Chatzileontiadou, Emma J. Grant, and Stephanie Gras. 2021. "SARS-CoV-2 Spike-Derived Peptides Presented by HLA Molecules" Biophysica 1, no. 2: 194-203. https://doi.org/10.3390/biophysica1020015
APA StyleNguyen, A. T., Szeto, C., Jayasinghe, D., Lobos, C. A., Halim, H., Chatzileontiadou, D. S. M., Grant, E. J., & Gras, S. (2021). SARS-CoV-2 Spike-Derived Peptides Presented by HLA Molecules. Biophysica, 1(2), 194-203. https://doi.org/10.3390/biophysica1020015