
The Effect of Calcium Ions on Mechanosensation and Neuronal Activity in Proprioceptive Neurons

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
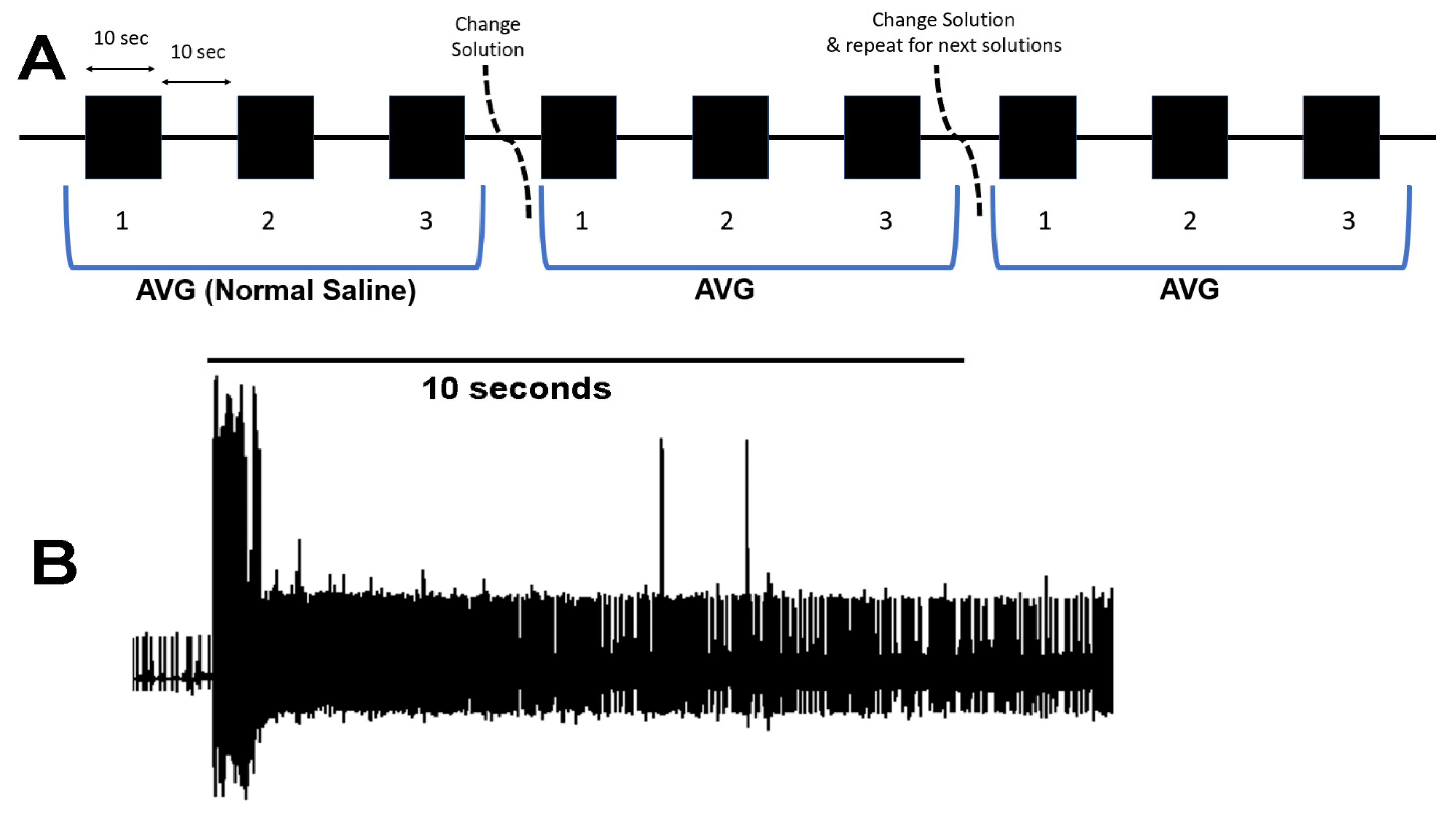
2.2. Dissection and Physiology
2.3. Statistical Analysis
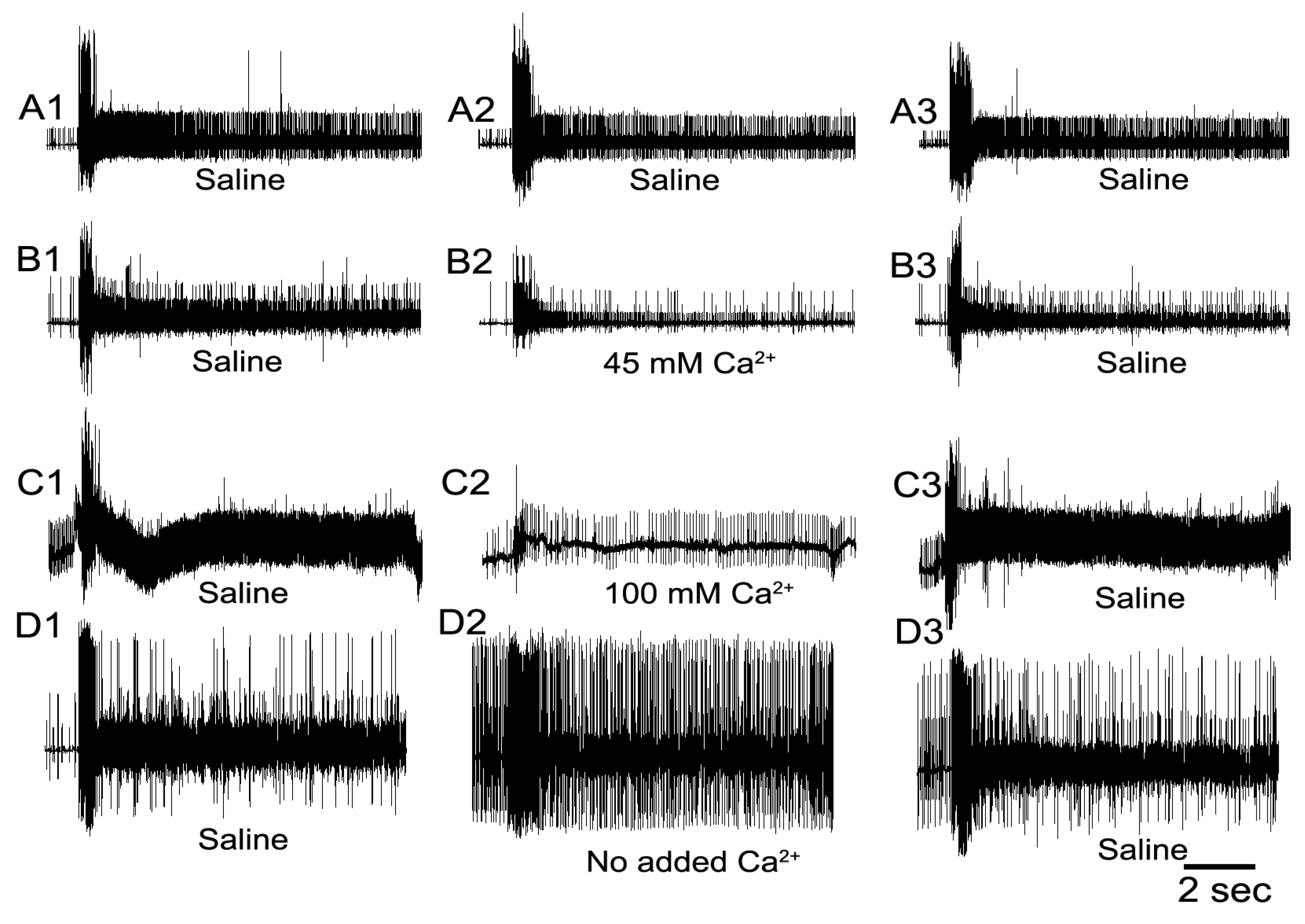
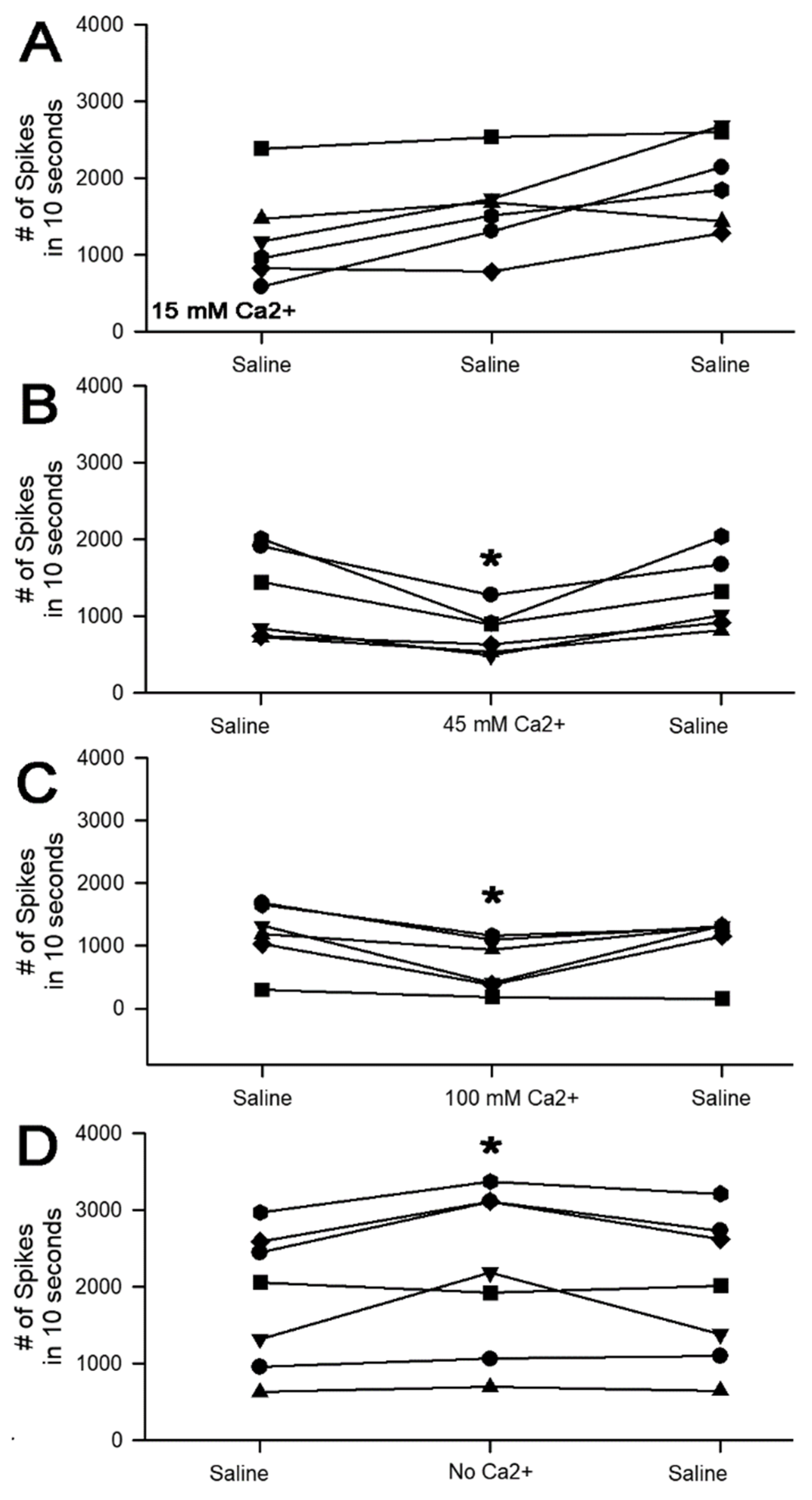
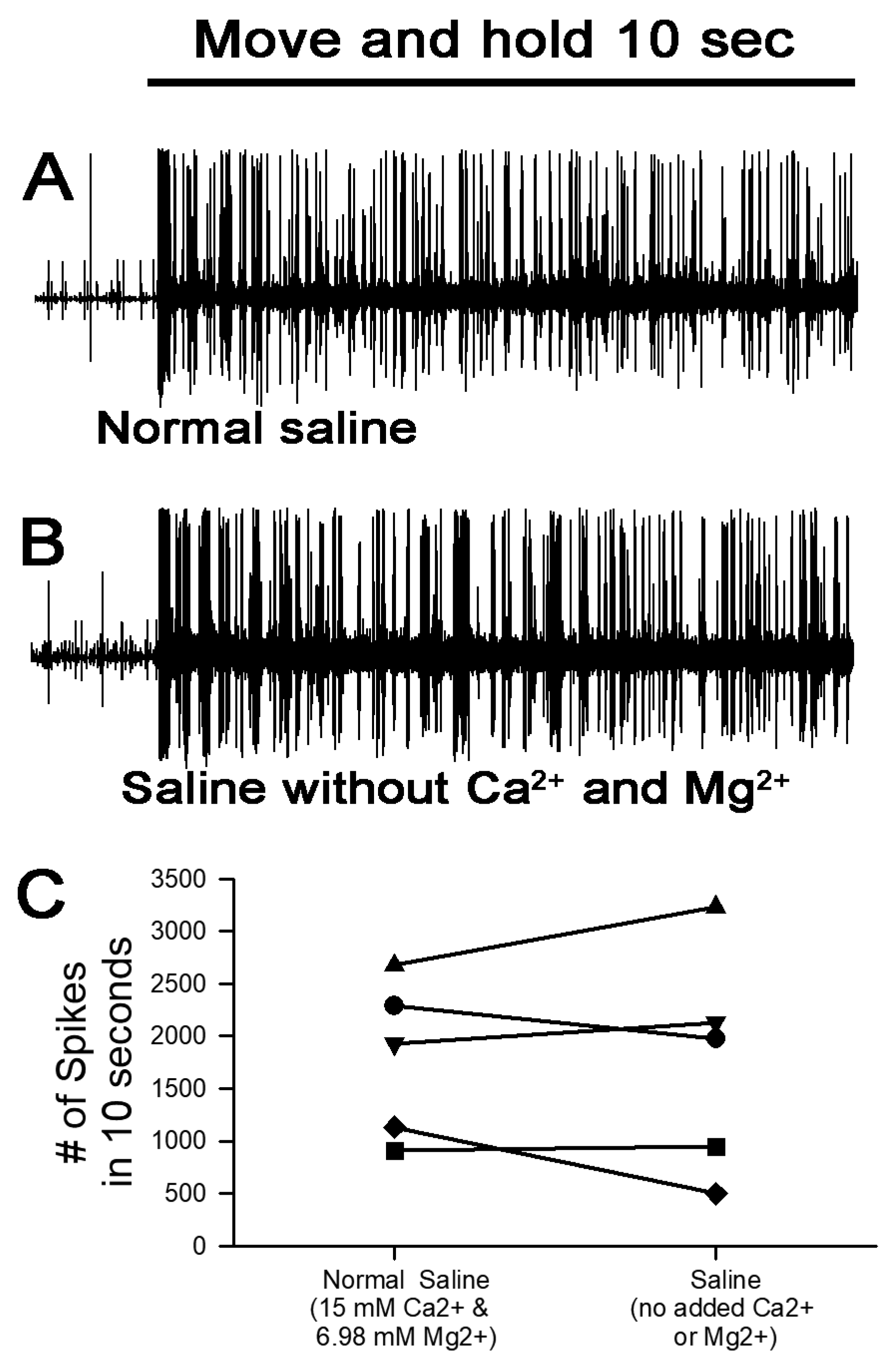
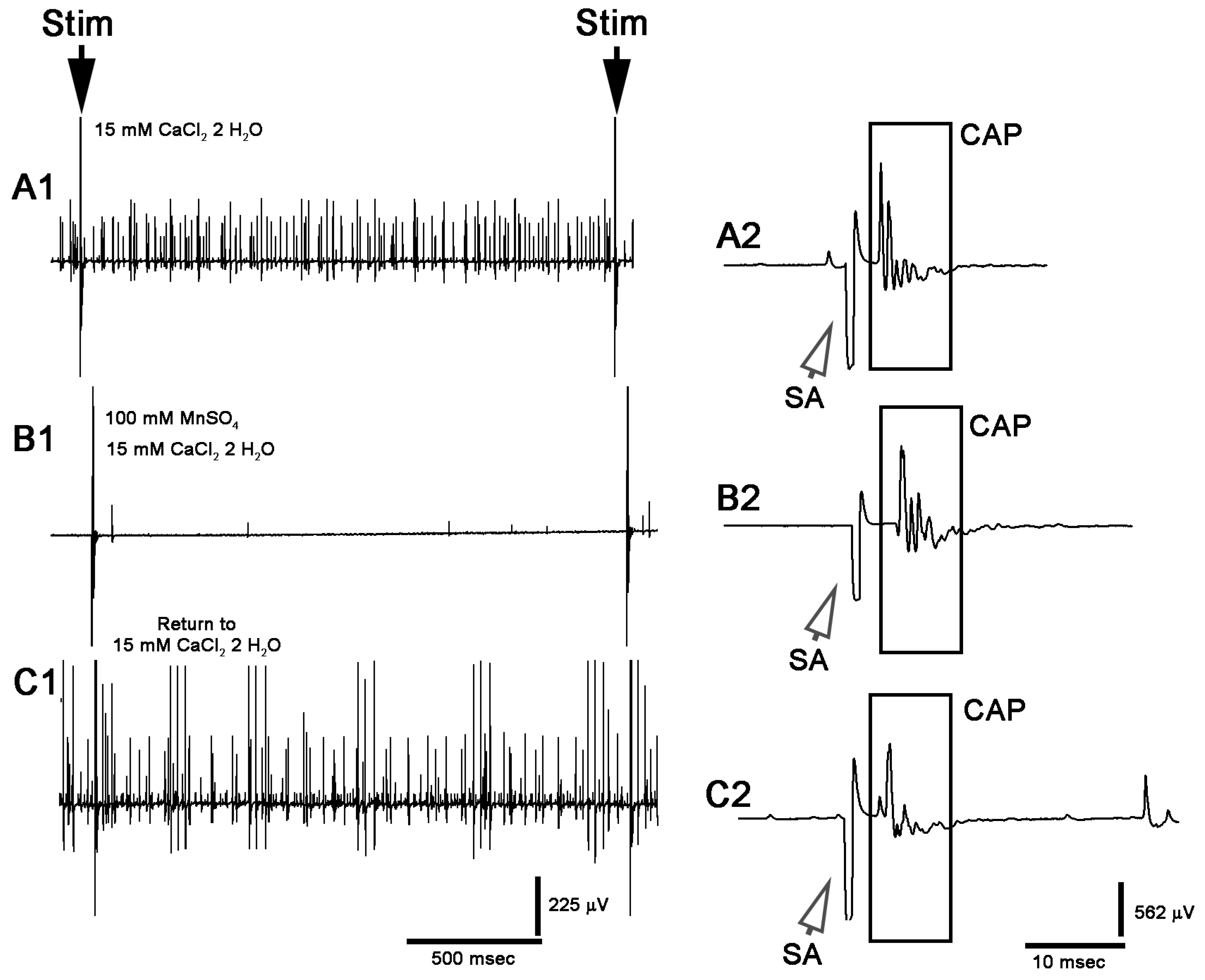
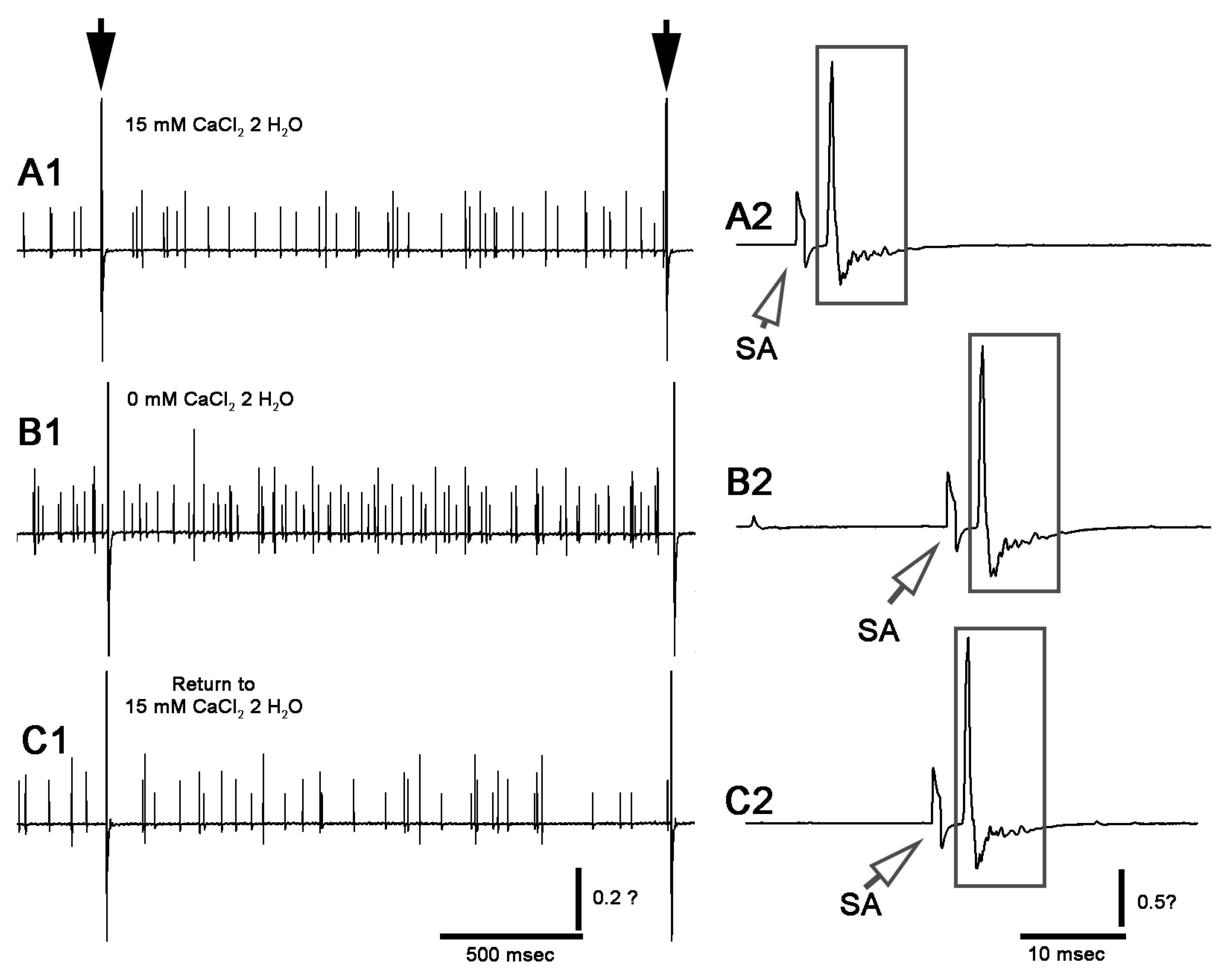
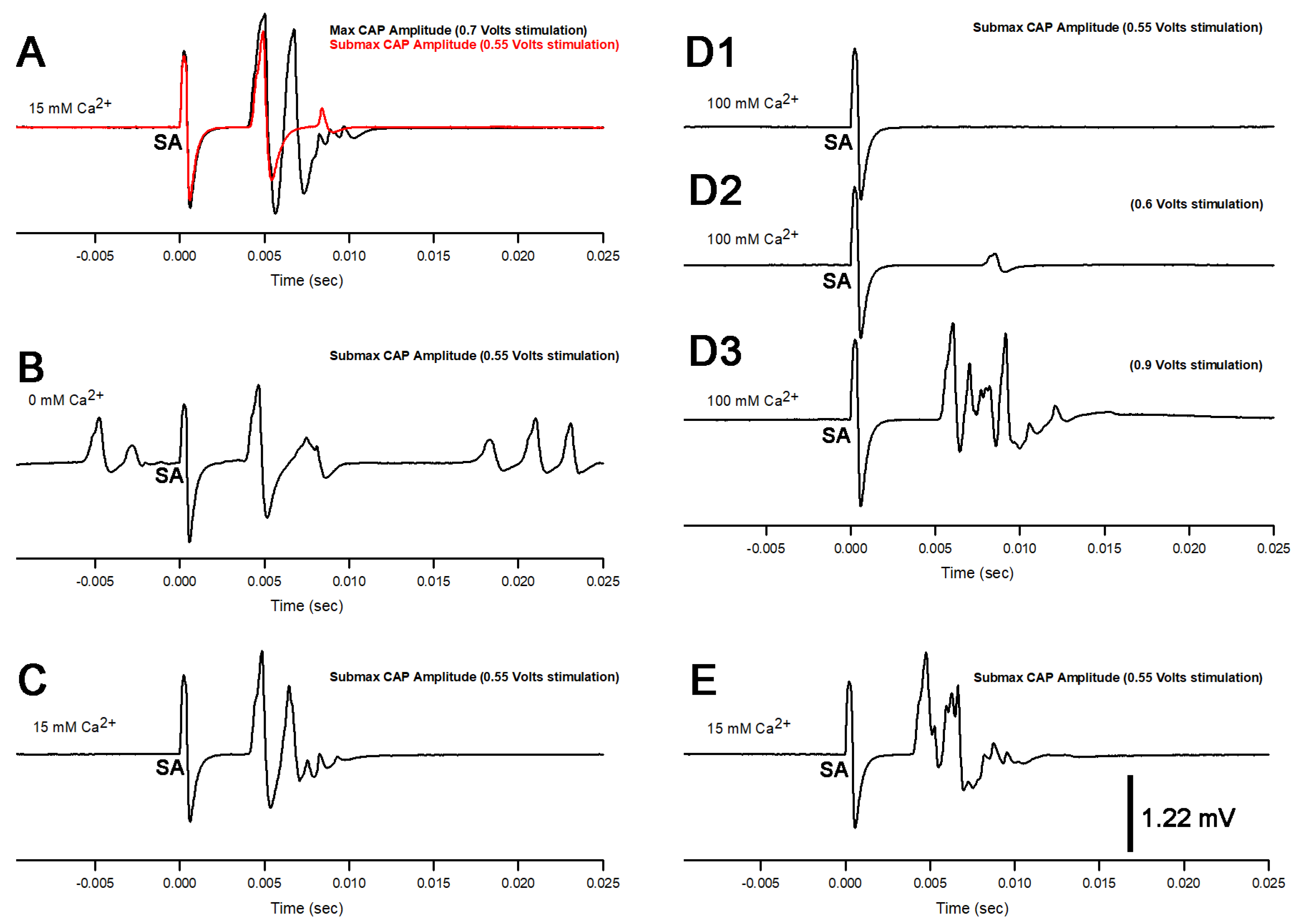
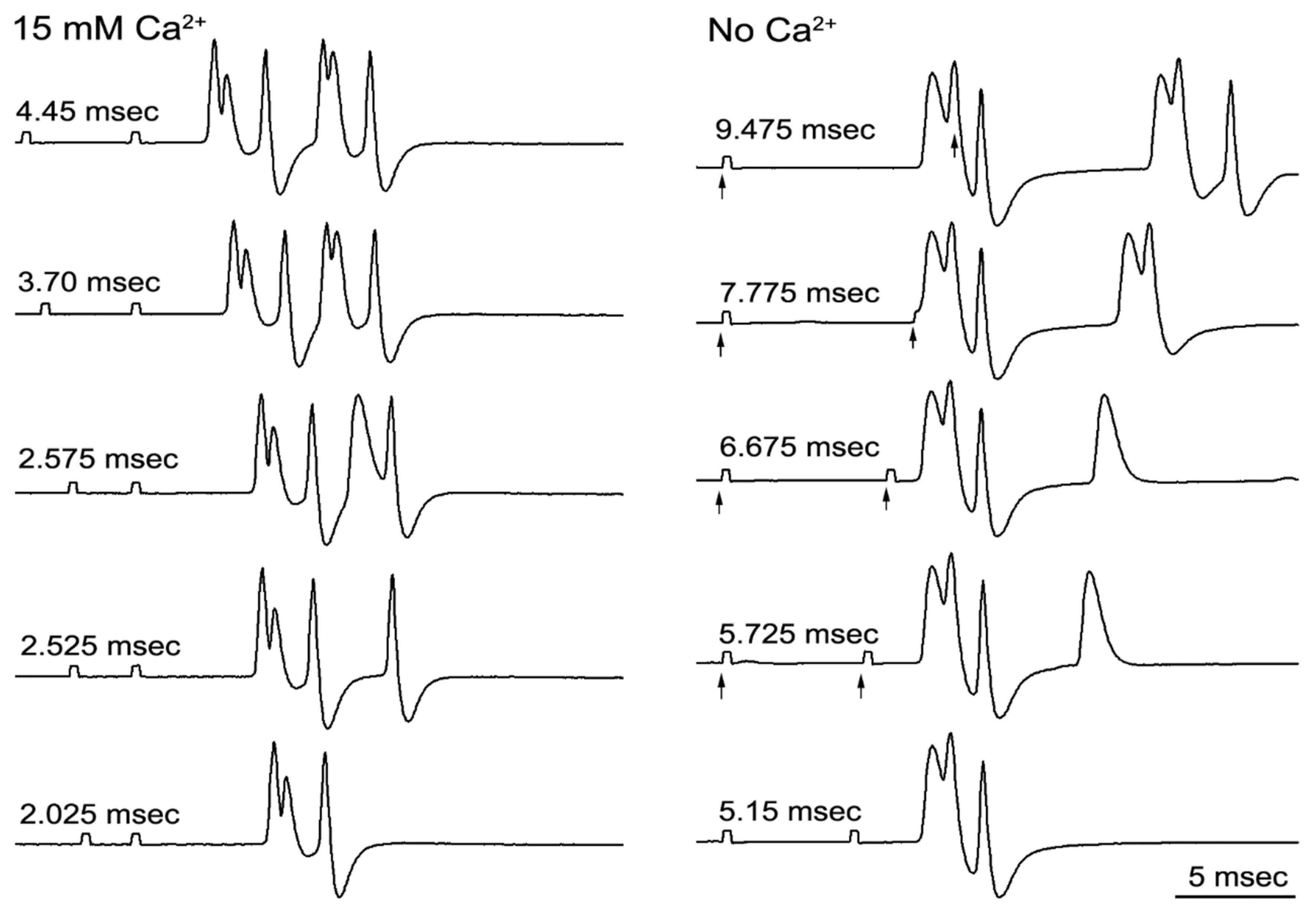
3. Results
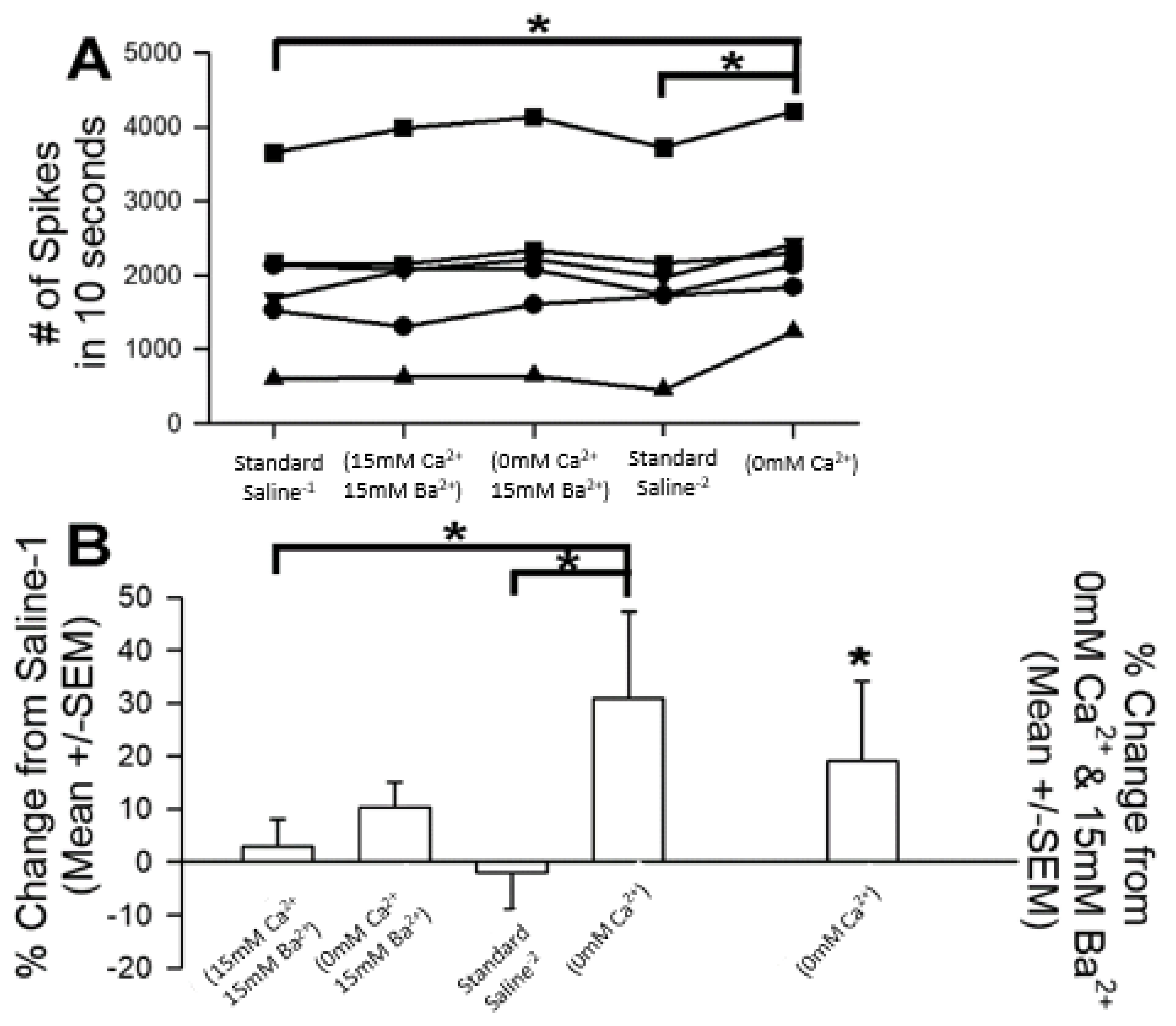
3.1. Altering Extracellular [Ca2+] Effects on Proprioceptive Activity
3.2. Refractory Periods
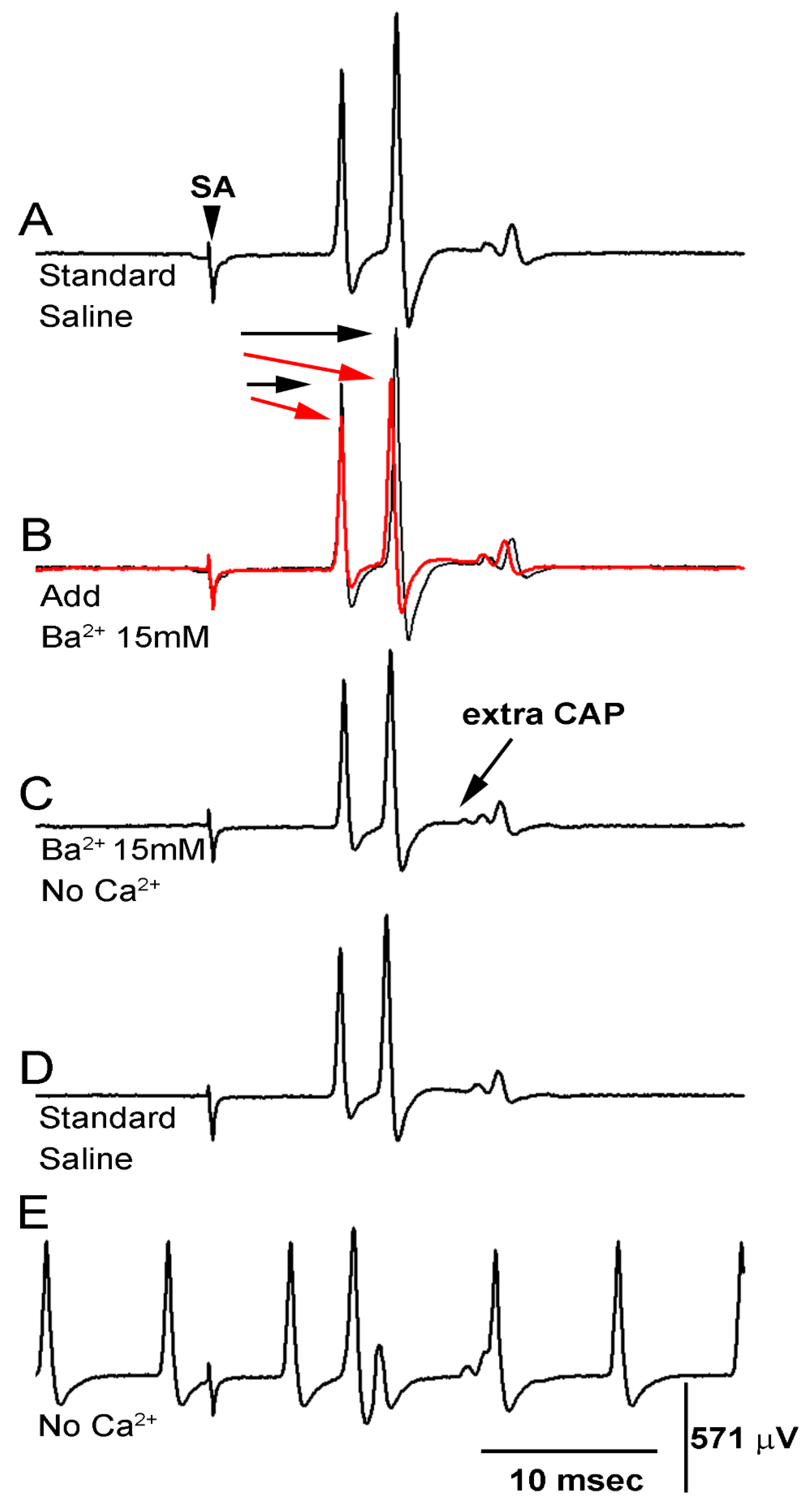
3.3. BaCl2 Experiments
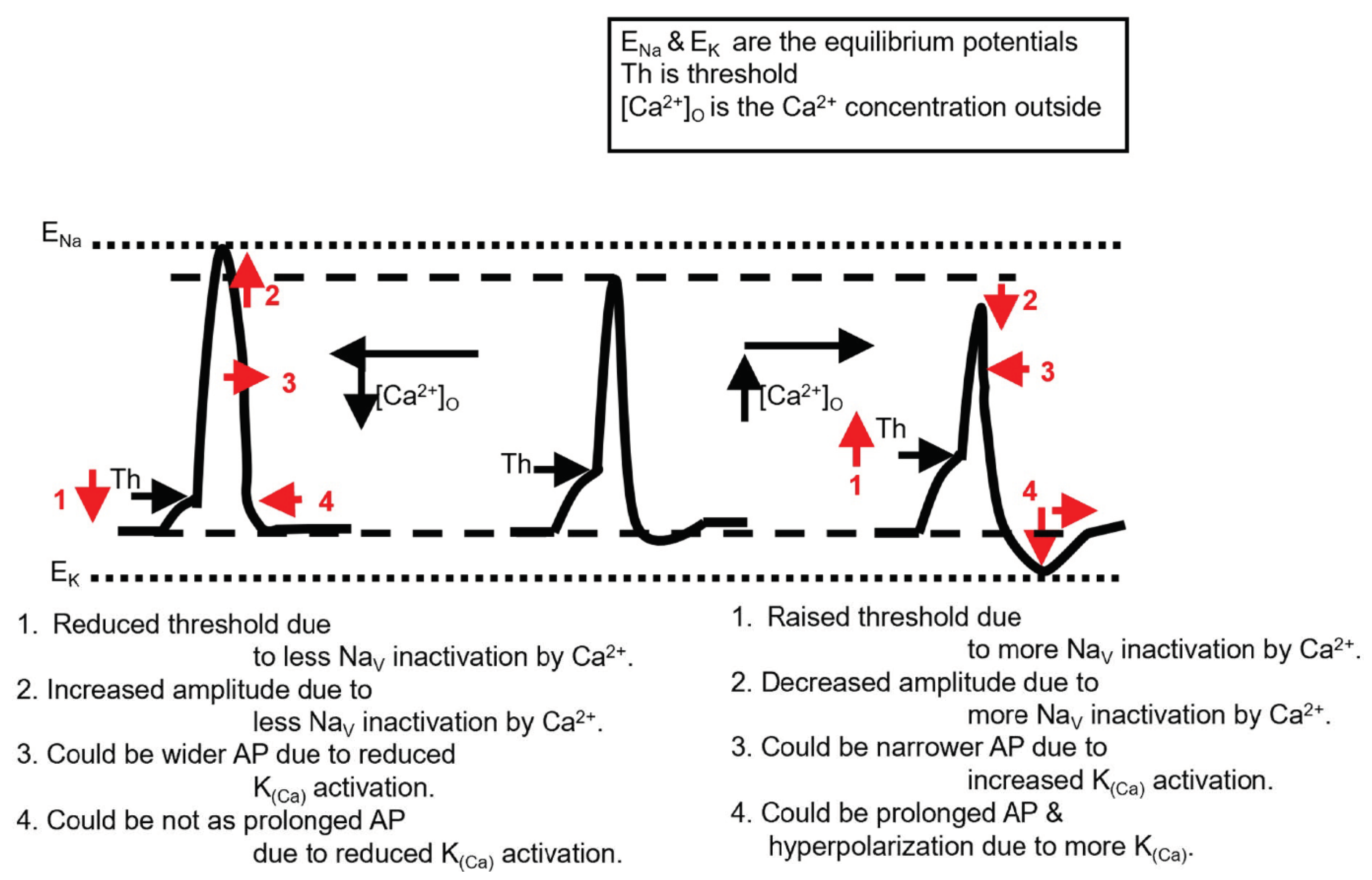
3.4. Model
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Katz, B. The Release of Neural Transmitter Substances; Liverpool University Press: Liverpool, UK, 1969. [Google Scholar]
- Augustine, G.J. How does calcium trigger neurotransmitter release? Curr. Opin. Neurobiol. 2001, 11, 320–326. [Google Scholar] [CrossRef]
- Peracchia, C. Chemical gating of gap junction channels: Roles of calcium, pH and calmodulin. Biochim. Biophys. Acta Biomembr. 2004, 1662, 61–80. [Google Scholar] [CrossRef]
- Duval, M.; Bach-Ngohou, K.; Masson, D.; Guimard, C.; Le Conte, P.; Trewick, D. Is severe hypocalcemia immediately life threatening? Endocr. Connect. 2018, 7, 1067–1074. [Google Scholar] [CrossRef]
- Odackal, J.; Sherpa, A.D.; Patel, N.; Colbourn, R.; Hrabetova, S. T-type calcium channels contribute to calcium disturbances in brain during hyponatremia. Exp. Neurol. 2015, 273, 105–113. [Google Scholar] [CrossRef][Green Version]
- Chebabo, S.R.; Hester, M.A.; Jing, J.; Aitken, P.G.; Somjen, G.G. Interstitial space, electrical resistance and ion concentrations during hypotonia of rat hippocampal slices. J. Physiol. 1995, 487, 685–697. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.P.; Dai, Z.-S. The Neuromuscular Junction Revisited: Ca2+ Channels and Transmitter Release in Cholinergic Neurones in Xenopus Nerve and Muscle Cell Culture. J. Exp. Biol. 1990, 153, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Scarnati, M.S.; Clarke, S.G.; Pang, Z.P.; Paradiso, K.G. Presynaptic Calcium Channel Open Probability and Changes in Calcium Influx Throughout the Action Potential Determined Using AP-Waveforms. Front. Synaptic Neurosci. 2020, 12, 17. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, C.M.; Cota, G. Calcium block of Na+ channels and its effect on closing rate. Proc. Natl. Acad. Sci. USA 1999, 96, 4154–4157. [Google Scholar] [CrossRef]
- Kamalanathan, S.; Balachandran, K.; Parthan, G.; Hamide, A. Chvostek’s sign: A video demonstration. BMJ Case Rep. 2012, 2012, bcr2012007098. [Google Scholar] [CrossRef]
- Patel, M.; Hu, E.W. Trousseau Sign. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Hujoel, I.A. The association between serum calcium levels and Chvostek sign: A population-based study. Neurol. Clin. Pract. 2016, 6, 321–328. [Google Scholar] [CrossRef]
- Shrimanker, I.; Bhattarai, S. Electrolytes. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Mackie, G.O.; Meech, R. Separate sodium and calcium spikes in the same axon. Nat. Cell Biol. 1985, 313, 791–793. [Google Scholar] [CrossRef]
- Jirounek, P.; Chardonness, E.; Brunet, P.C. Afterpotentials in nonmyelinated nerve fibers. J. Neurophysiol. 1991, 65, 860–873. [Google Scholar] [CrossRef] [PubMed]
- Robert, A.; Jirounek, P. Uptake of potassium by nonmyelinating Schwann cells induced by axonal activity. J. Neurophysiol. 1994, 72, 2570–2579. [Google Scholar] [CrossRef] [PubMed]
- Mert, T.; Gunes, Y.; Guven, M.; Günay, I.; Ozcengiz, D. Effects of calcium and magnesium on peripheral nerve conduction. Pol. J. Pharmacol. 2003, 55, 25–30. [Google Scholar]
- Han, P.; Trinidad, B.J.; Shi, J. Hypocalcemia-induced seizure: Demystifying the calcium paradox. ASN Neuro. 2015, 7, 1759091415578050. [Google Scholar] [CrossRef] [PubMed]
- Bostock, H.; Sears, T.A.; Sherratt, R.M. The effects of 4-aminopyridine and tetraethylammonium ions on normal and demyelinated mammalian nerve fibres. J. Physiol. 1981, 313, 301–315. [Google Scholar] [CrossRef]
- Armstrong, C.M. Distinguishing surface effects of calcium ion from pore-occupancy effects in Na+ channels. Proc. Natl. Acad. Sci. USA 1999, 96, 4158–4163. [Google Scholar] [CrossRef]
- Narahashi, T.; Frazier, D.T.; Takeno, K. Effects of calcium on the local anesthetic suppression of ionic conductances in squid axon membranes. J. Pharmacol. Exp. Ther. 1976, 197, 426–438. [Google Scholar]
- Majeed, Z.R.; Titlow, J.; Hartman, H.B.; Cooper, R. Proprioception and tension receptors in crab limbs: Student laboratory exercises. J. Vis. Exp. 2013, 80, e51050. [Google Scholar] [CrossRef]
- Woo, S.-H.; Lukacs, V.; De Nooij, J.C.; Zaytseva, D.; Criddle, C.R.; Francisco, A.G.; Jessell, T.M.; Wilkinson, K.A.; Patapoutian, A. Piezo2 is the principal mechanotransduction channel for proprioception. Nat. Neurosci. 2015, 18, 1756–1762. [Google Scholar] [CrossRef]
- Dayaram, V.; Malloy, C.; Martha, S.R.; Alvarez, B.; Chukwudolue, I.; Dabbain, N.; Mahmood, D.; Goleva, S.; Hickey, T.; Ho, A.; et al. The Effect of CO2, Intracellular pH and Extracellular pH on Mechanosensory Proprioceptor Responses in Crayfish and Crab. Am. J. Undergrad. Res. 2017, 14, 85–99. [Google Scholar] [CrossRef]
- Dayaram, V.; Malloy, C.; Martha, S.; Alvarez, B.; Chukwudolue, I.; Dabbain, N.; D.mahmood, D.; Goleva, S.; Hickey, T.; Ho, A.; et al. Stretch activated channels in proprioceptive chordotonal organs of crab and crayfish are sensitive to Gd3+ but not amiloride, ruthenium red or low pH. IMPLUSE Prem. Undergrad. Neurosci. J. 2017. Available online: https://impulse.appstate.edu/issues/2017 (accessed on 15 October 2021).
- McCubbin, S.; Jeoung, A.; Waterbury, C.; Cooper, R.L. Pharmacological profiling of stretch activated channels in proprioceptive neurons. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2020, 233, 108765. [Google Scholar] [CrossRef]
- Stanback, M.; Stanback, A.E.; Akhtar, S.; Basham, R.; Chithrala, B.; Collis, B.; Heberle, B.A.; Higgins, E.; Lane, A.; Marella, S.; et al. The Effect of Lipopolysaccharides on Primary Sensory Neurons in Crustacean Models. IMPLUSE 2019. Available online: https://impulse.appstate.edu/articles/2019/effect-lipopolysaccharides-primary-sensory-neurons-crustacean-models (accessed on 20 October 2021).
- Stanley, C.E.; Adams, R.; Nadolski, J.; Amrit, E.; Barrett, M.; Bohnett, C.; Campbell, K.; Deweese, K.; Dhar, S.; Gillis, B.; et al. The effects of tricaine mesylate on arthropods: Crayfish, crab and Drosophila. Invertebr. Neurosci. 2020, 20, 1–16. [Google Scholar] [CrossRef]
- Murthy, S.E.; Dubin, A.E.; Whitwam, T.; Jojoa-Cruz, S.; Cahalan, S.M.; Mousavi, S.A.R.; Ward, A.B.; Patapoutian, A. OSCA/TMEM63 are an evolutionarily conserved family of mechanically activated ion channels. eLife 2018, 7, 41844. [Google Scholar] [CrossRef] [PubMed]
- Malloy, C.; Dayaram, V.; Martha, S.R.; Alvarez, B.; Chukwudolue, I.; Dabbain, N.; Mahmood, D.D.; Goleva, S.; Hickey, T.; Ho, A.; et al. The effects of potassium and muscle homogenate on proprioceptive responses in crayfish and crab. J. Exp. Zool. Part A Ecol. Integr. Physiol. 2017, 327, 366–379. [Google Scholar] [CrossRef]
- Pankau, C.; Nadolski, J.; Tanner, H.; Cryer, C.; Di Girolamo, J.; Haddad, C.; Lanning, M.; Miller, M.; Neely, D.; Wilson, R.; et al. Effects of manganese on physiological processes in Drosophila, crab and crayfish: Cardiac, neural and behavioral assays. Comp. Biochem. Physiol. C 2021, in press. [Google Scholar]
- Castelfranco, A.M.; Hartline, D.K. Evolution of rapid nerve conduction. Brain Res. 2016, 1641, 11–33. [Google Scholar] [CrossRef]
- Cooper, R.L. Mapping proprioceptive neurons on chordotonal organs in the crab, Cancer magister. Crustaceana 2008, 81, 447–475. [Google Scholar] [CrossRef]
- Clare, J.D.J.; Townsend, P.A.; Anhalt-Depies, C.; Locke, C.; Stenglein, J.L.; Frett, S.; Martin, K.J.; Singh, A.; Van Deelen, T.R.; Zuckerberg, B. Making inference with messy (citizen science) data: When are data accurate enough and how can they be improved? Ecol. Appl. 2019, 29, e01849. [Google Scholar] [CrossRef] [PubMed]
- Wycoff, S.; Weineck, K.; Conlin, S.; Grau, E.; Bradley, A.; Cantrell, D.; Eversole, S.; Grachen, C.; Hall, K.; Hawthorne, D.; et al. Investigating Potential Effects of Clove Oil (Eugenol) in Model Crustaceans. IMPLUSE 2018, 1–21. Available online: https://impulse.appstate.edu/articles/2018/effects-clove-oil-eugenol-proprioceptive-neurons-heart-rate-and-behavior-model-crustac (accessed on 20 October 2021).
- Bakshi, A.; Patrick, L.E.; Wischusen, E.W. A Framework for Implementing Course-Based Undergraduate Research Experiences (CUREs) in Freshman Biology Labs. Am. Biol. Teach. 2016, 78, 448–455. [Google Scholar] [CrossRef]
- Linn, M.C.; Palmer, E.; Baranger, A.; Gerard, E.; Stone, E. Undergraduate research experiences: Impacts and opportunities. Science 2015, 347, 1261757. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | PD Organ | CAPs |
|---|---|---|
| Standard saline | Normal | Normal |
| Saline + Ba2+ (15 mM) | Normal | ↓ Amplitude |
| Saline + Ba2+ (15 mM) & No Ca2+ | slight ↑ Activity | ↓ Amplitude |
| ↑ Activity | ||
| Standard saline | Normal | Normal |
| Saline no Ca2+ | ↑↑ Activity | ↓ Amplitude |
| ↑↑ Activity |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Atkins, D.E.; Bosh, K.L.; Breakfield, G.W.; Daniels, S.E.; Devore, M.J.; Fite, H.E.; Guo, L.Z.; Henry, D.K.J.; Kaffenberger, A.K.; Manning, K.S.; et al. The Effect of Calcium Ions on Mechanosensation and Neuronal Activity in Proprioceptive Neurons. NeuroSci 2021, 2, 353-371. https://doi.org/10.3390/neurosci2040026
Atkins DE, Bosh KL, Breakfield GW, Daniels SE, Devore MJ, Fite HE, Guo LZ, Henry DKJ, Kaffenberger AK, Manning KS, et al. The Effect of Calcium Ions on Mechanosensation and Neuronal Activity in Proprioceptive Neurons. NeuroSci. 2021; 2(4):353-371. https://doi.org/10.3390/neurosci2040026
Chicago/Turabian StyleAtkins, Devan E., Kimberly L. Bosh, Grace W. Breakfield, Sydney E. Daniels, Makayla J. Devore, Hailey E. Fite, Landys Z. Guo, Danielle K. J. Henry, Alana K. Kaffenberger, Katherine S. Manning, and et al. 2021. "The Effect of Calcium Ions on Mechanosensation and Neuronal Activity in Proprioceptive Neurons" NeuroSci 2, no. 4: 353-371. https://doi.org/10.3390/neurosci2040026
APA StyleAtkins, D. E., Bosh, K. L., Breakfield, G. W., Daniels, S. E., Devore, M. J., Fite, H. E., Guo, L. Z., Henry, D. K. J., Kaffenberger, A. K., Manning, K. S., Mowery, T. E., Pankau, C. L., Parker, N., Serrano, M. E., Shakhashiro, Y., Tanner, H. N., Ward, R. A., Wehry, A. H., & Cooper, R. L. (2021). The Effect of Calcium Ions on Mechanosensation and Neuronal Activity in Proprioceptive Neurons. NeuroSci, 2(4), 353-371. https://doi.org/10.3390/neurosci2040026