Expression of Wilm’s Tumor Gene (WT1) in Endometrium with Potential Link to Gestational Vascular Transformation


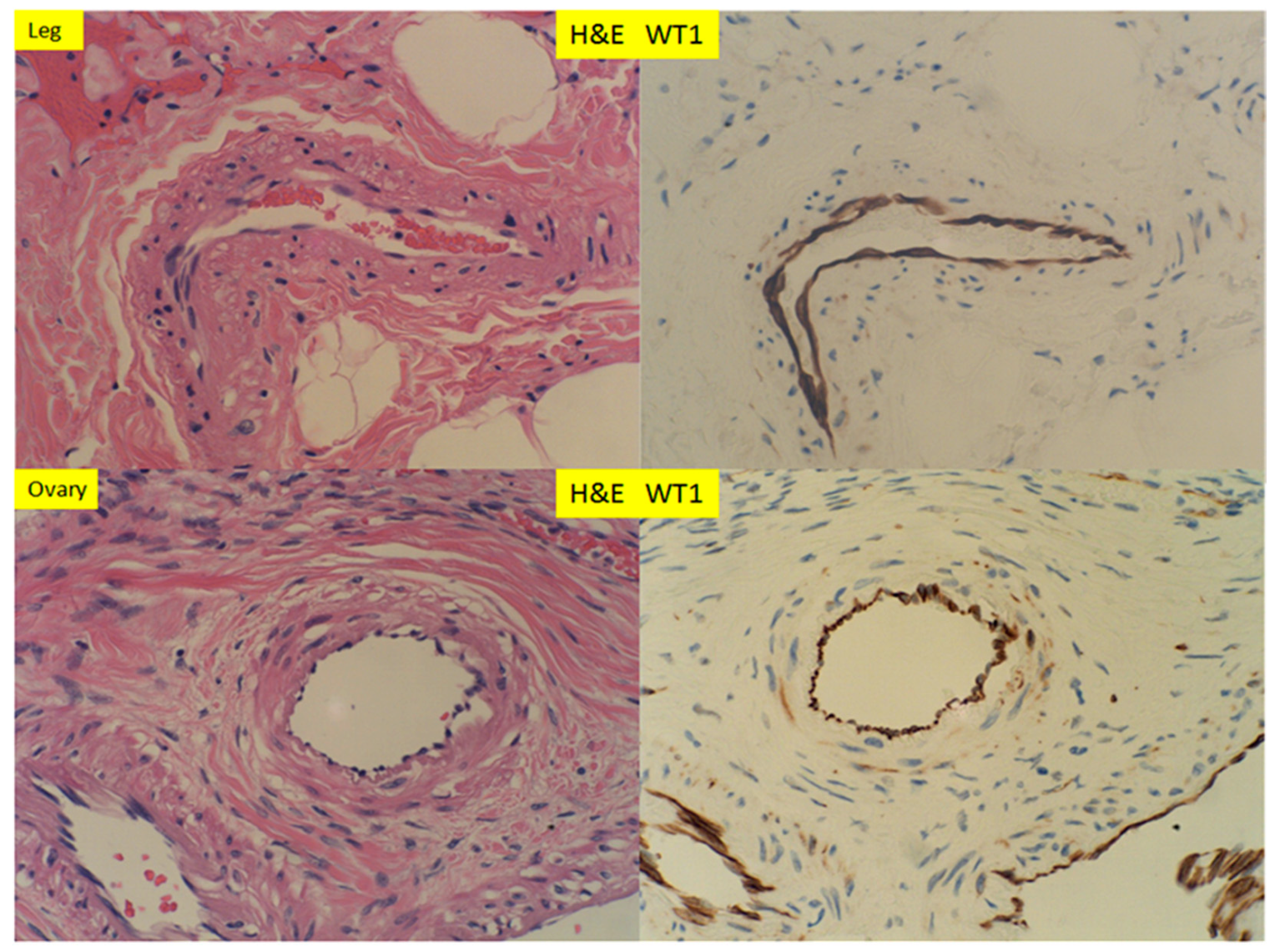{kind=link}
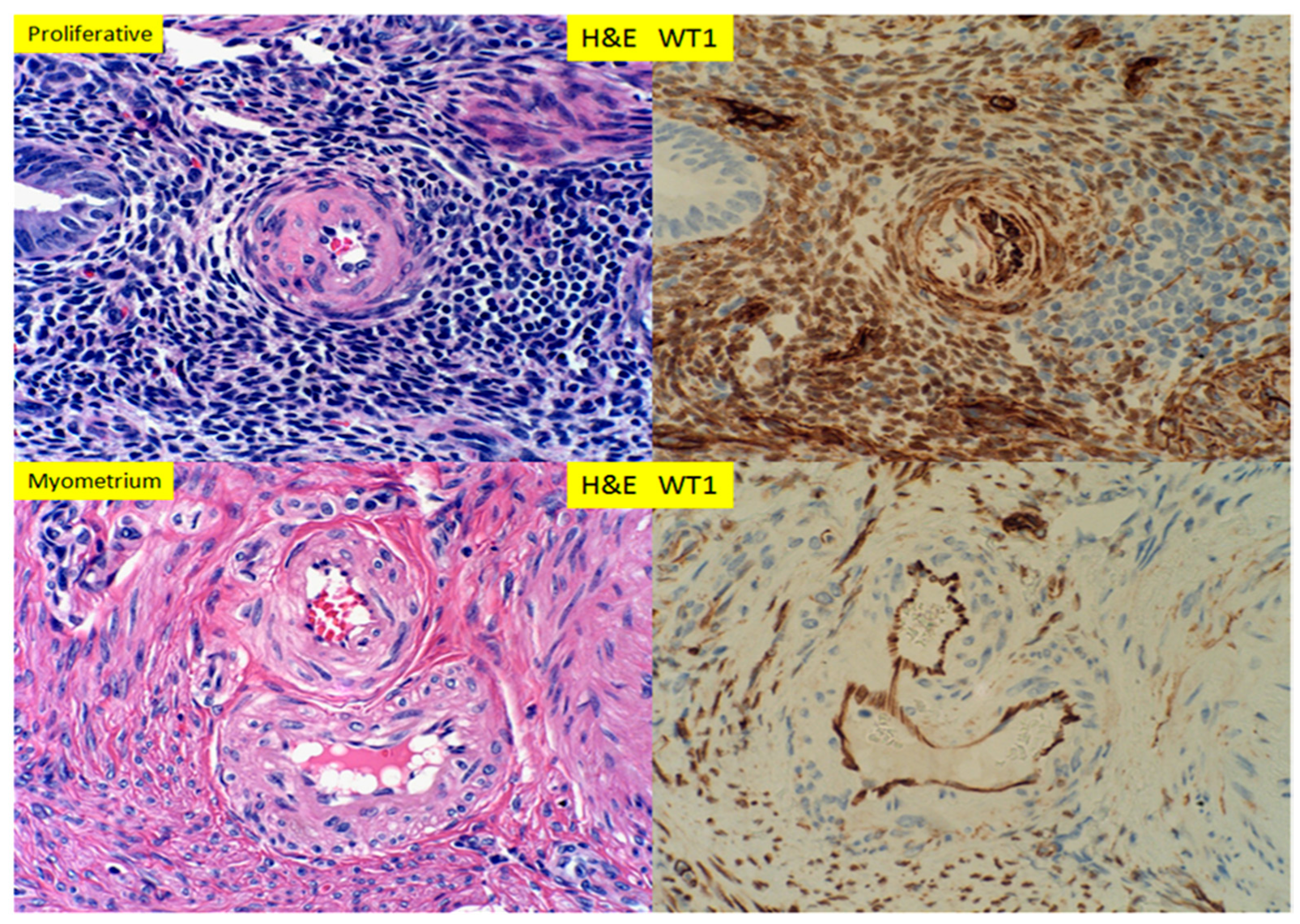{kind=link}
{kind=link}
{kind=link}
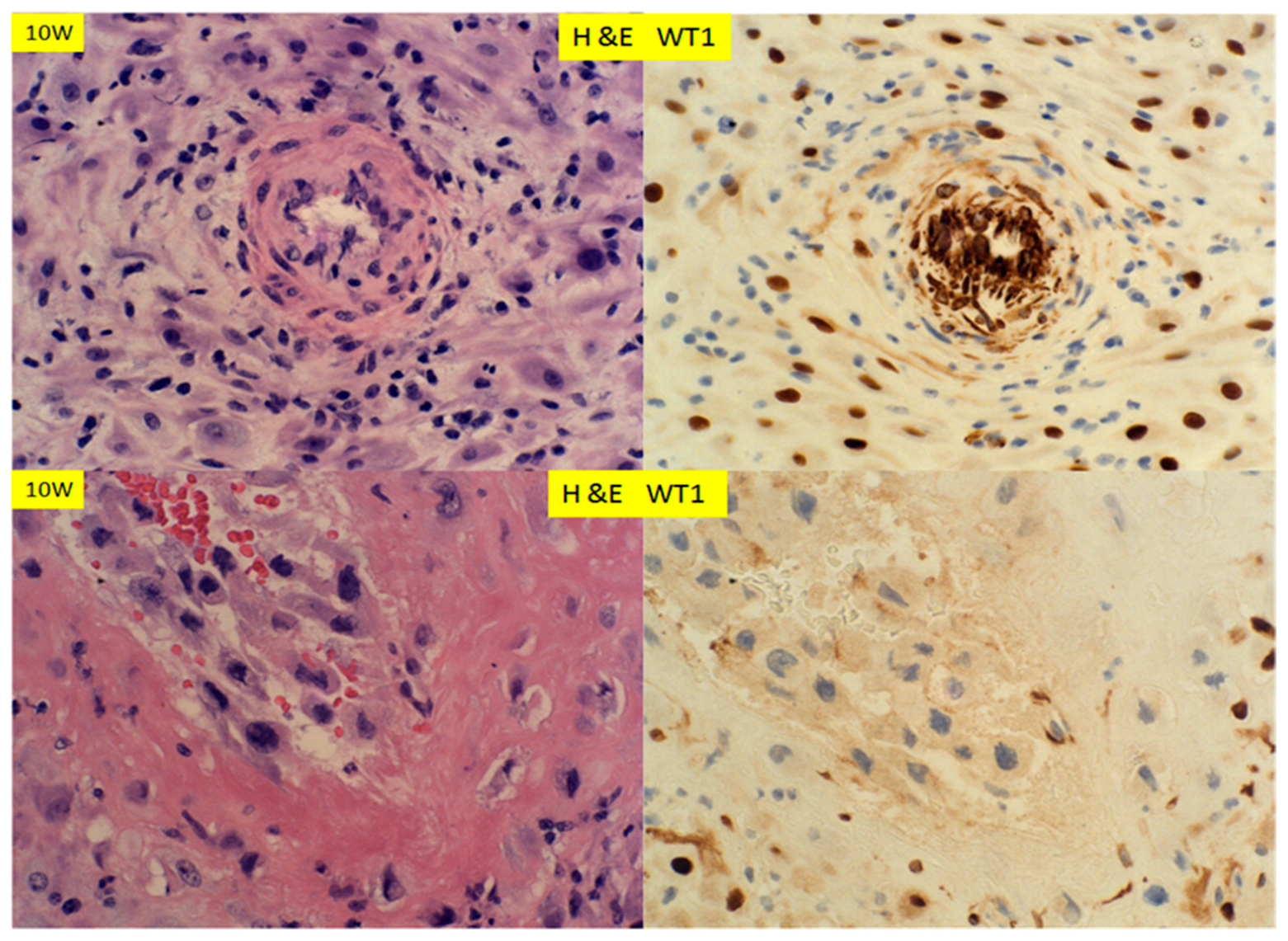{kind=link}
{kind=link}
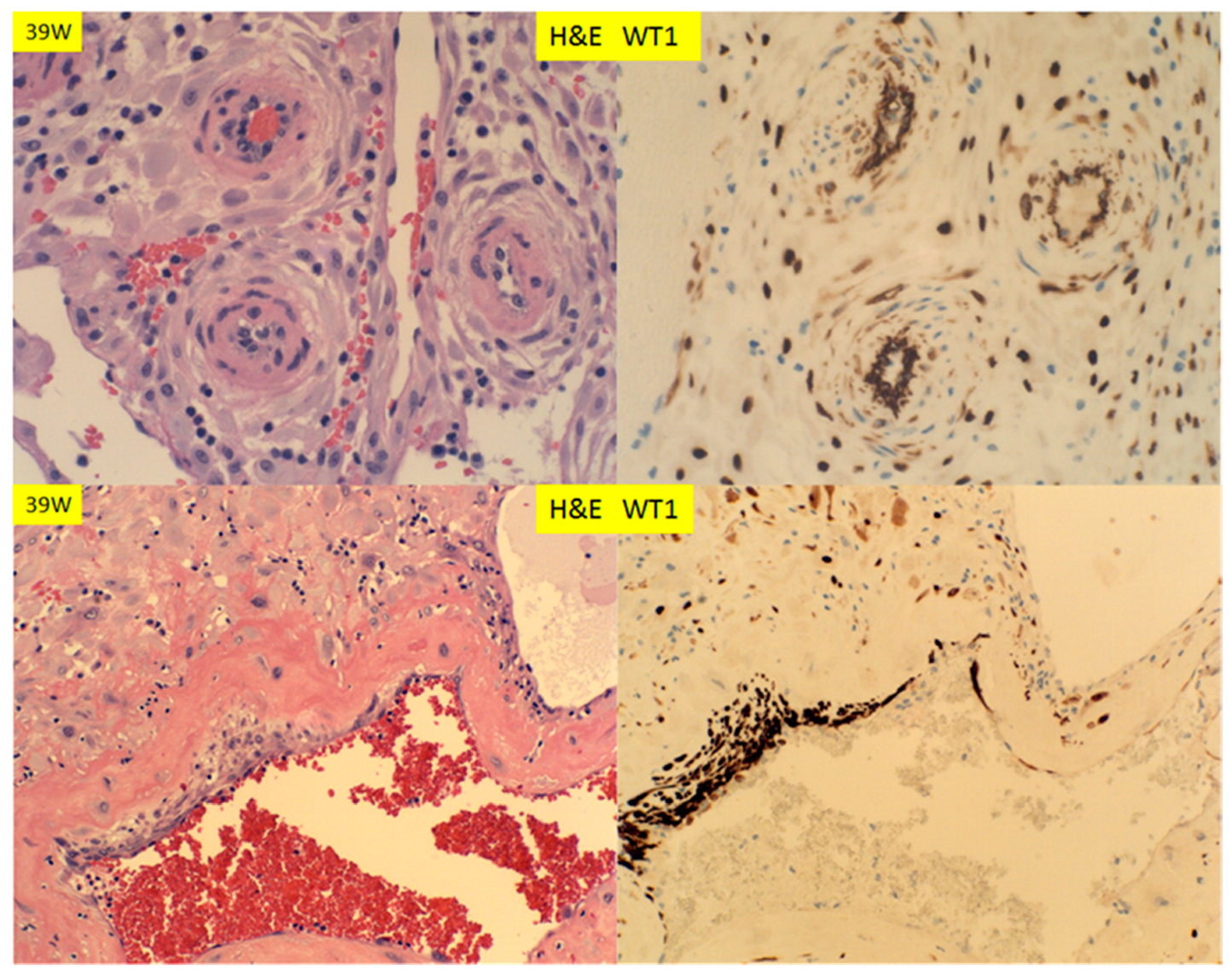{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
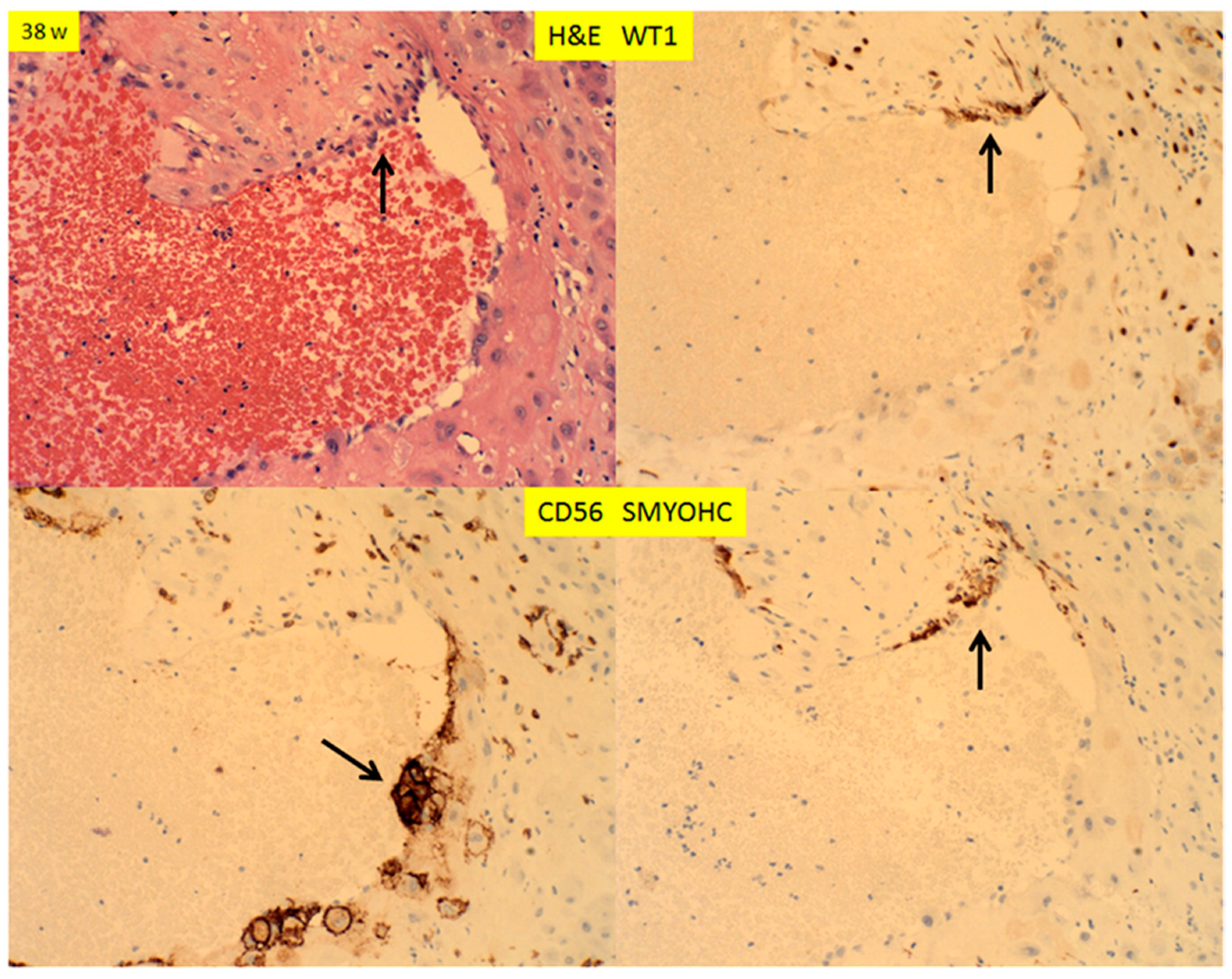
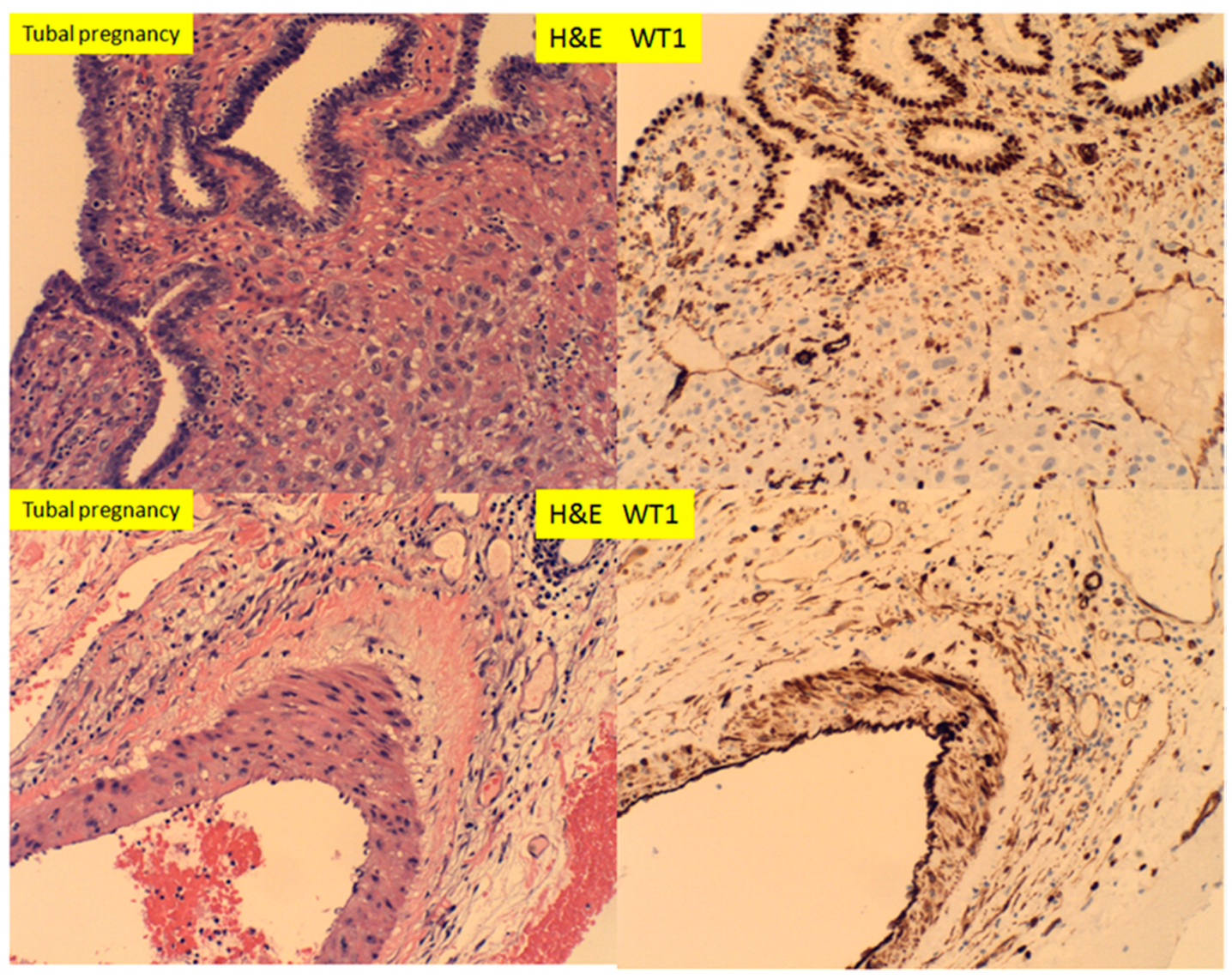
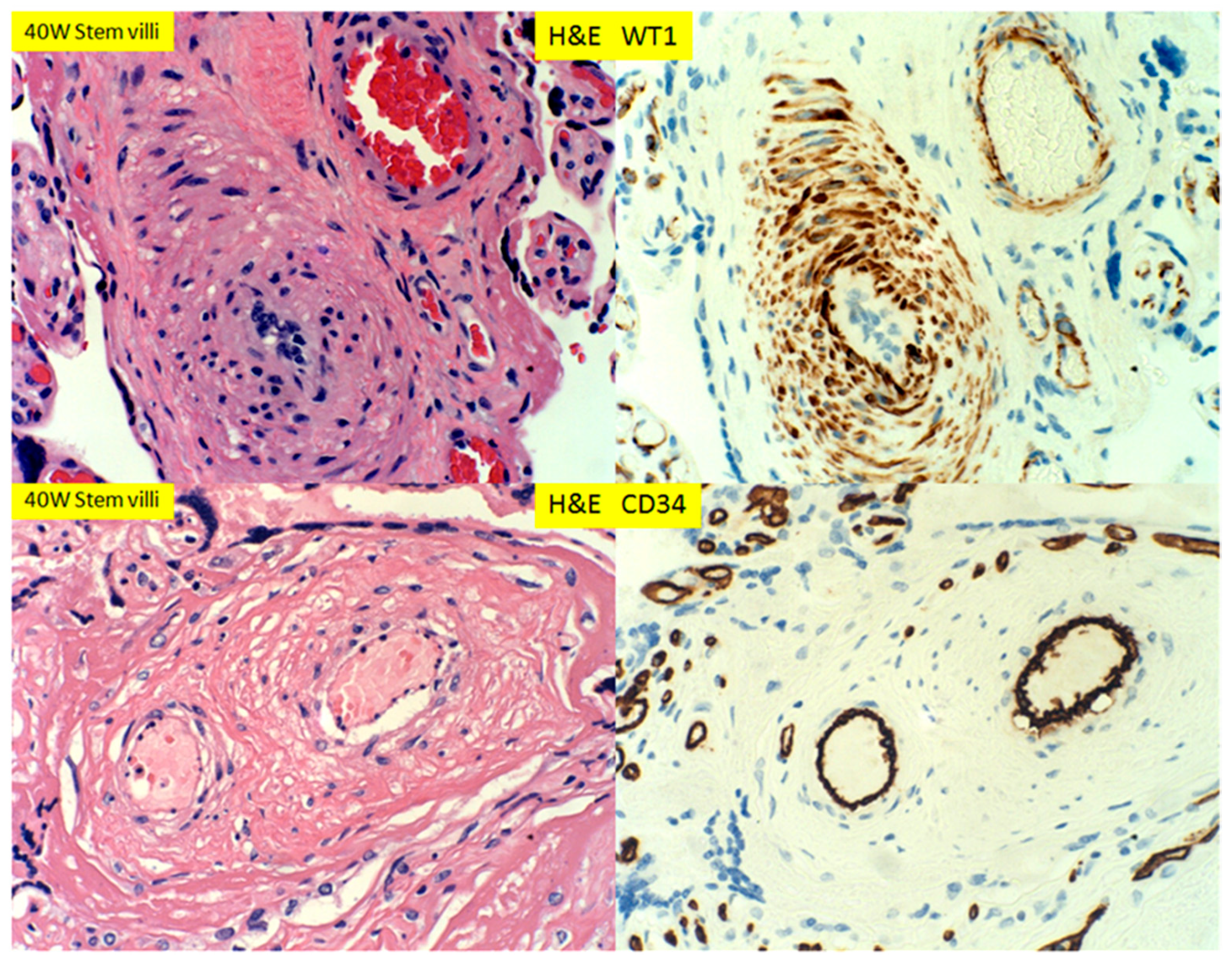
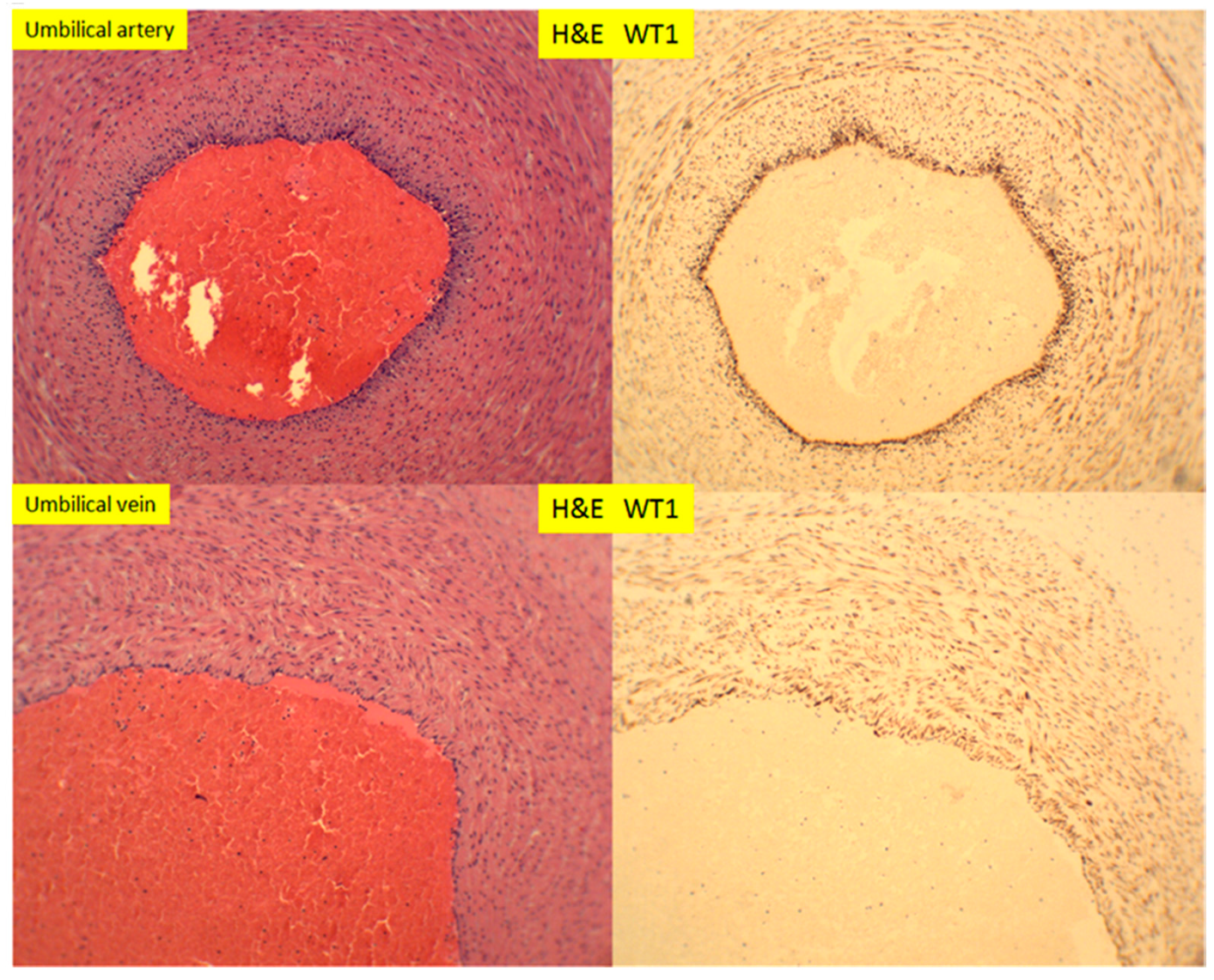
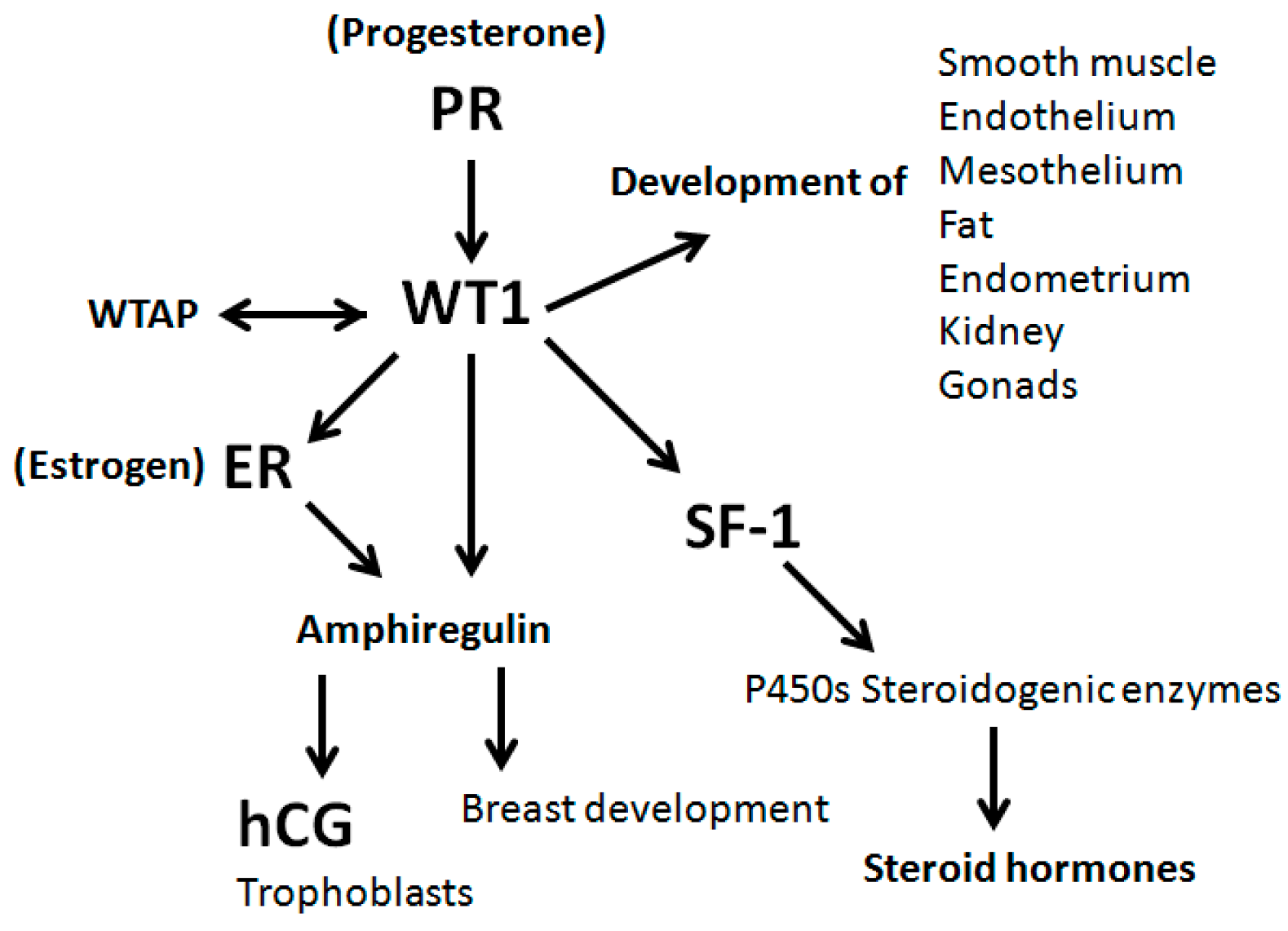
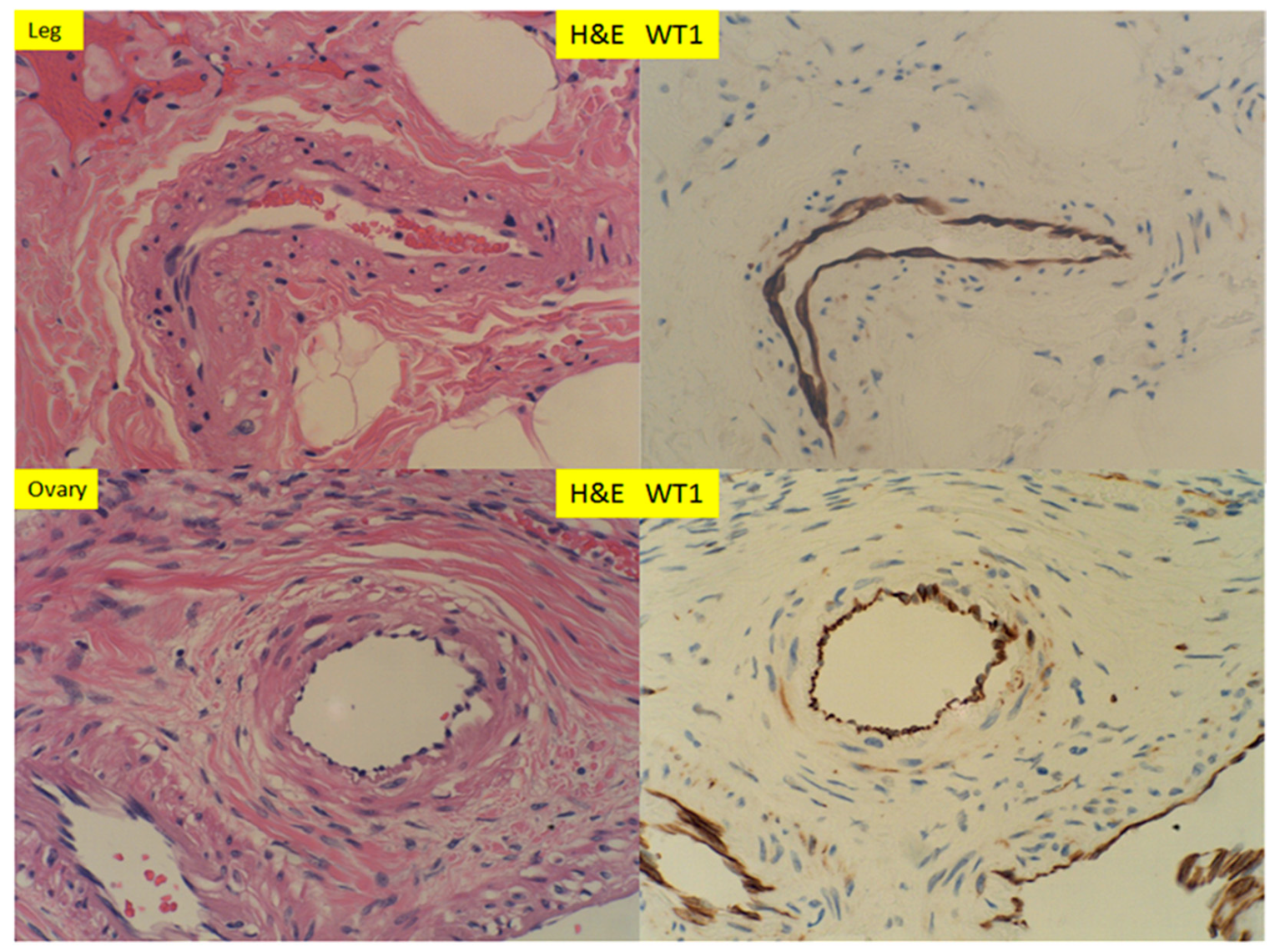
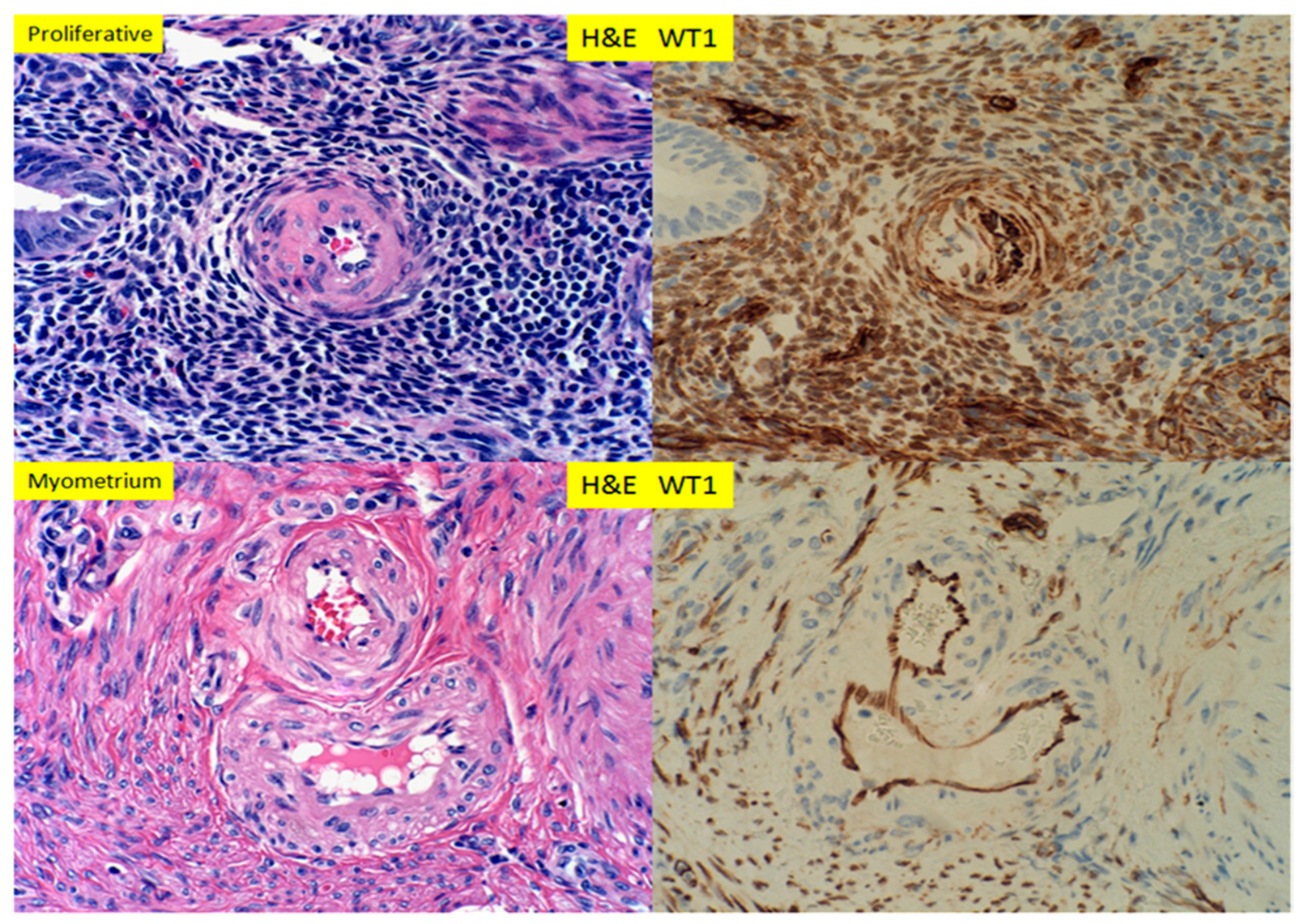
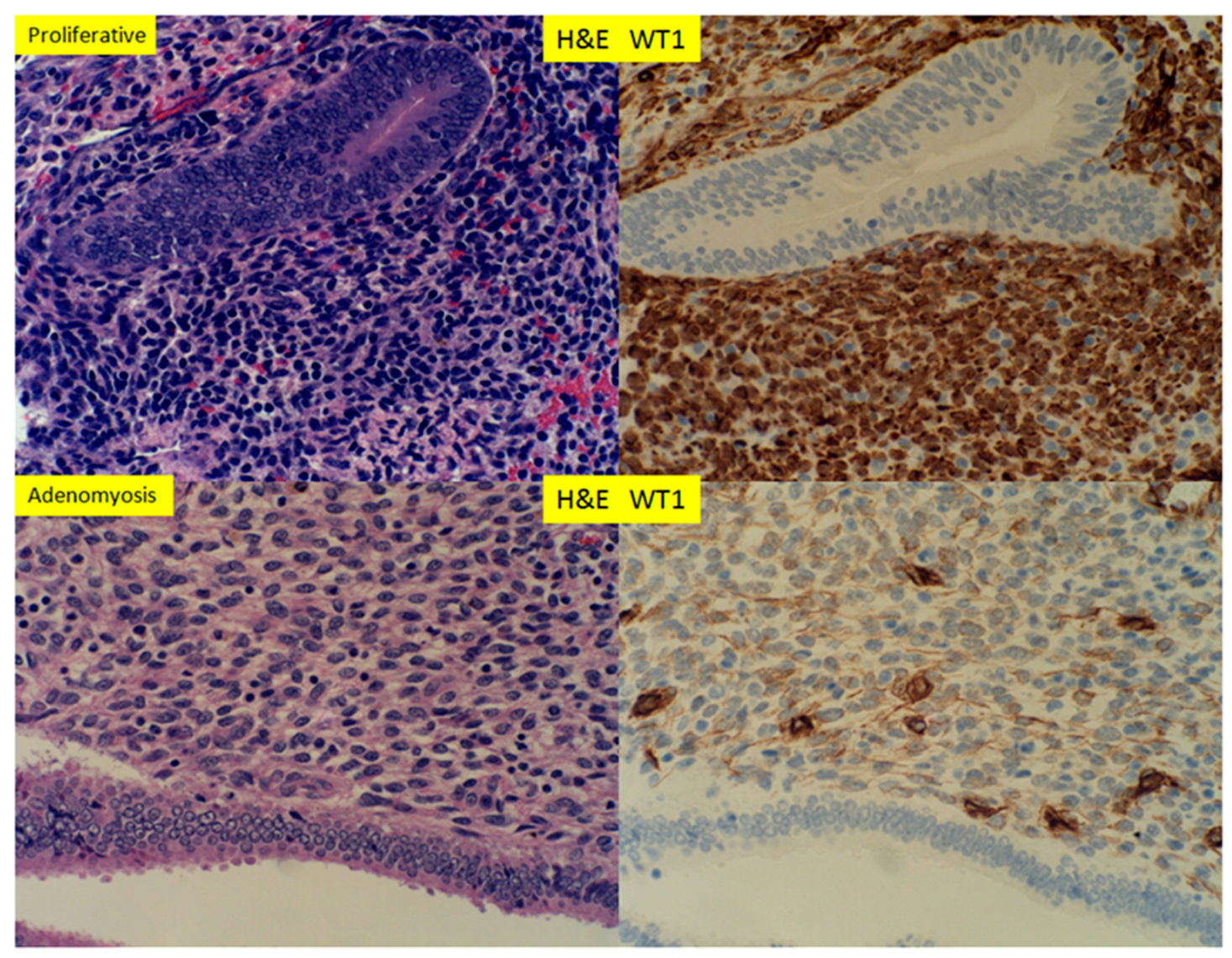
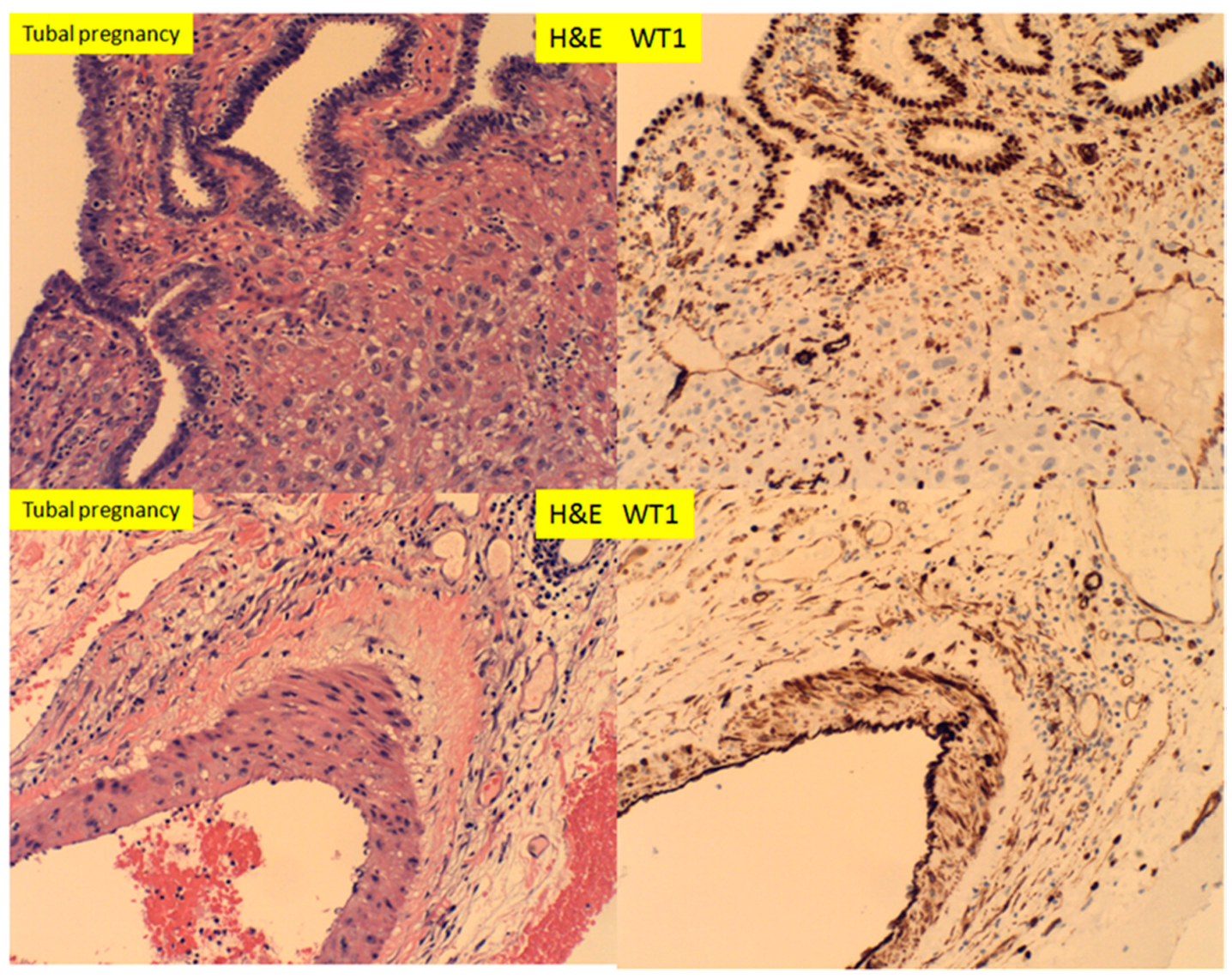
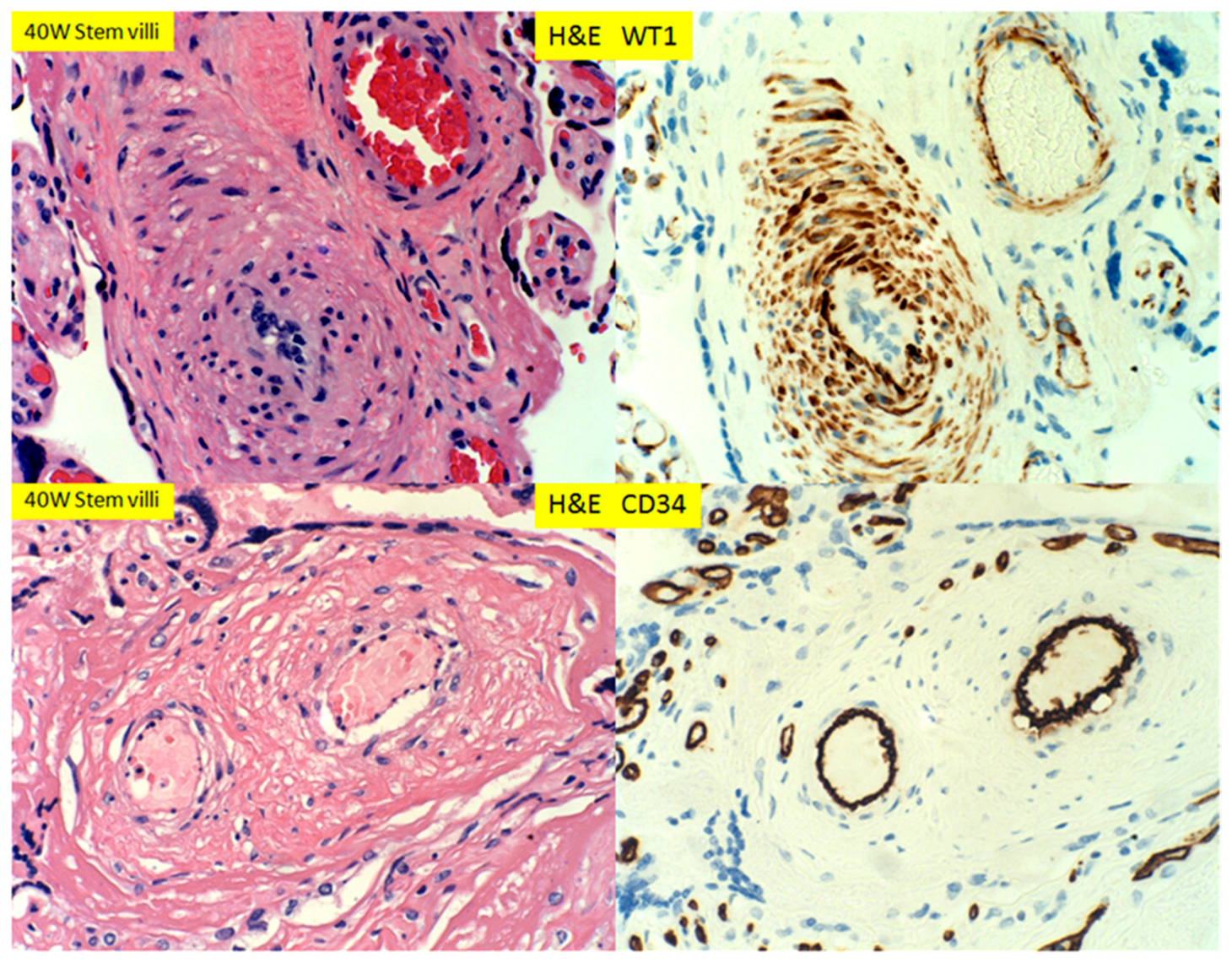
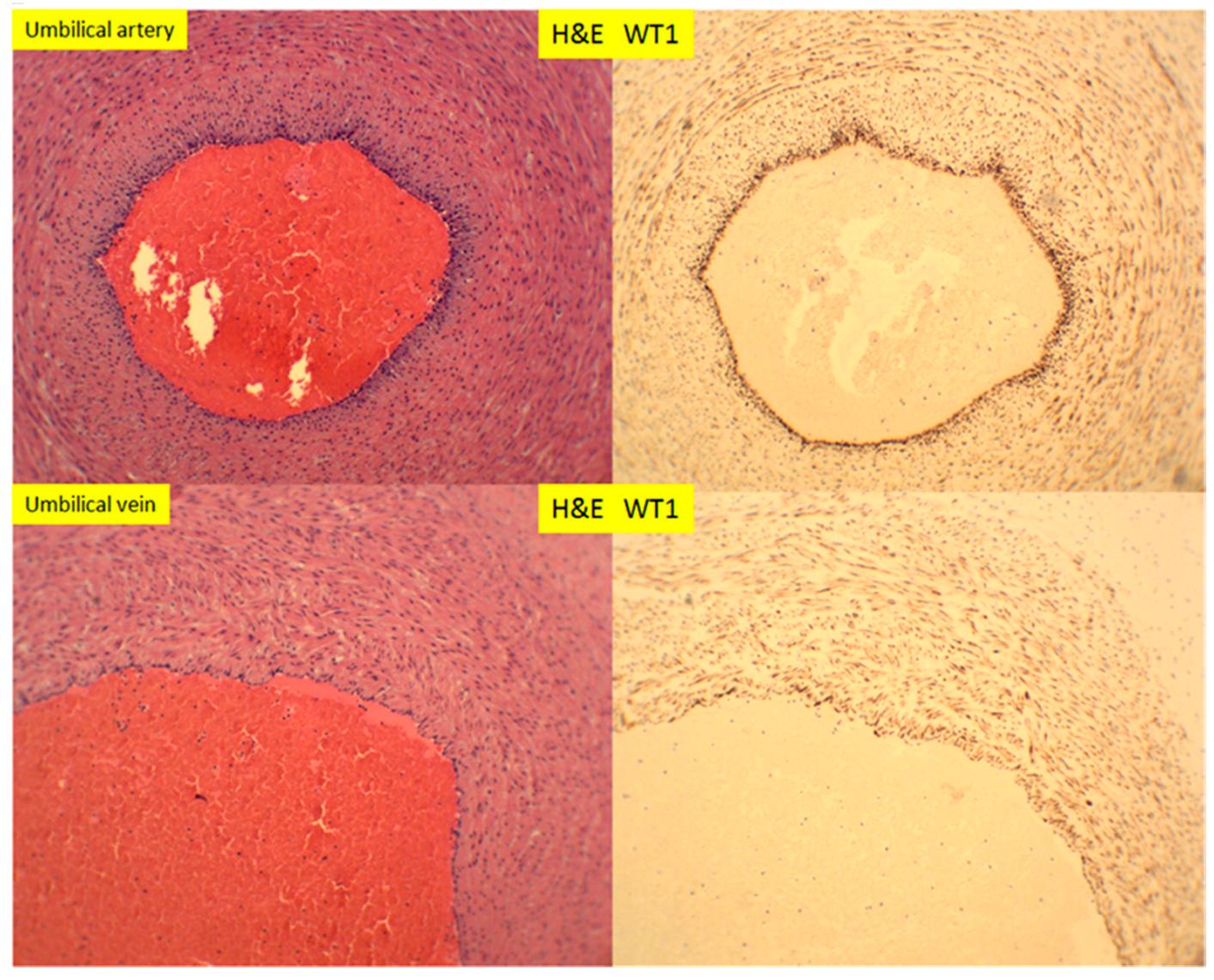
3. Results
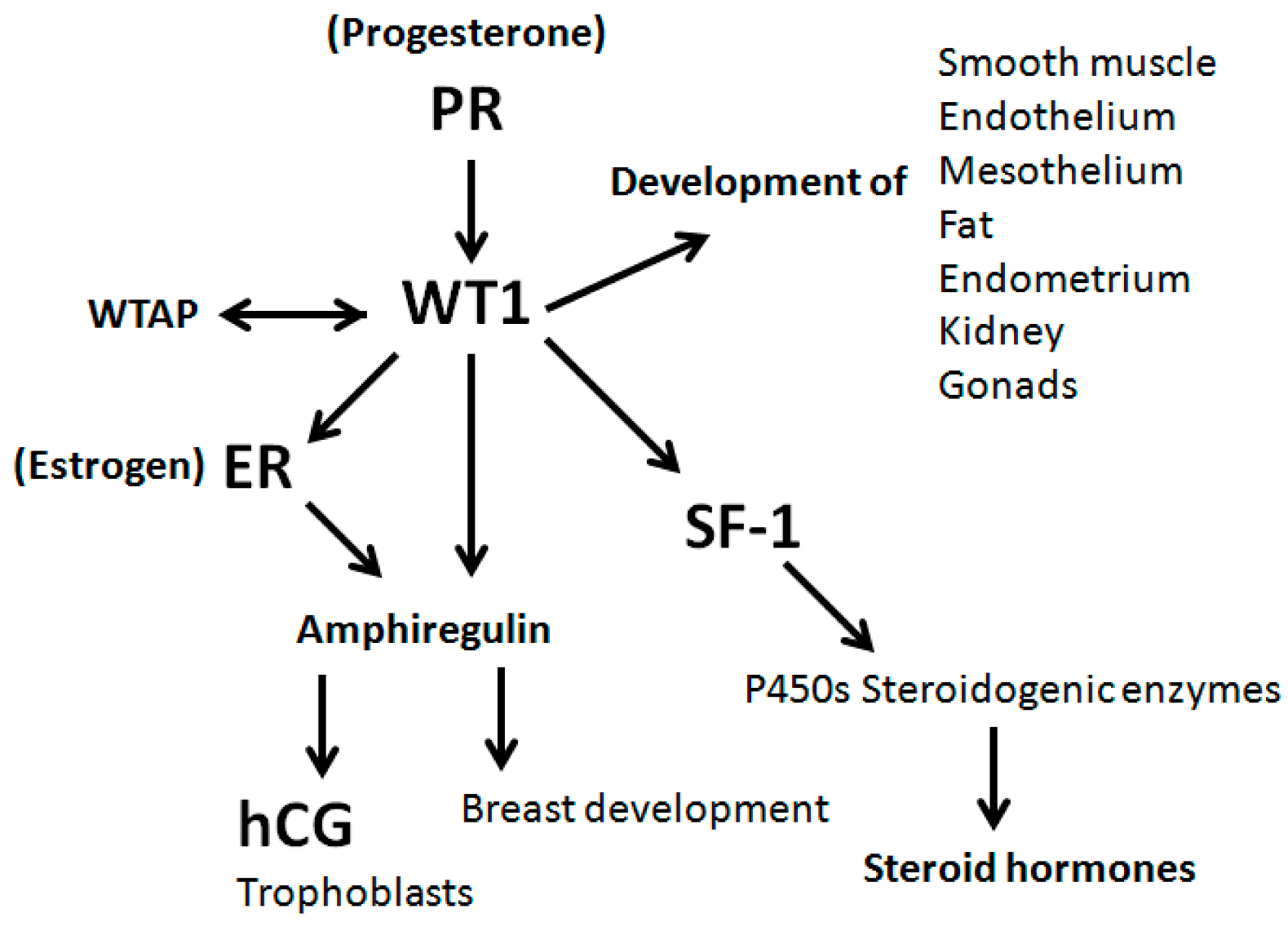
4. Discussion
Funding
Acknowledgments
Conflicts of Interest
References
- Benirschke, K.; Burton, G.J.; Baergen, R.N. Pathology of the Human Placenta, 6th ed.; Springer: New York, NY, USA, 2012. [Google Scholar]
- Cunningham, F.G. Williams Obstetrics, 25th ed.; McGraw-Hill: New York, NY, USA, 2018; Volume xvi, p. 1328. [Google Scholar]
- Pijnenborg, R.; Vercruysse, L.; Hanssens, M. The uterine spiral arteries in human pregnancy: Facts and controversies. Placenta 2006, 27, 939–958. [Google Scholar] [CrossRef] [PubMed]
- Harris, L.K.; Smith, S.D.; Keogh, R.J.; Jones, R.; Baker, P.; Knöfler, M.; Cartwright, J.E.; Whitley, G.S.; Aplin, J. Trophoblast- and vascular smooth muscle cell-derived MMP-12 mediates elastolysis during uterine spiral artery remodeling. Am. J. Pathol. 2010, 177, 2103–2115. [Google Scholar] [CrossRef] [PubMed]
- Harris, L.K.; Benagiano, M.; D’Elios, M.M.; Brosens, I.; Benagiano, G. Placental bed research: II. Functional and immunological investigations of the placental bed. Am. J. Obstet. Gynecol. 2019, 221, 457–469. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P. Decidual vasculopathy and spiral artery remodeling revisited II: Relations to trophoblastic dependent and independent vascular transformation. J. Matern. Neonatal Med. 2020, 1–7. [Google Scholar] [CrossRef]
- Zhang, P. Phenotypic Switch of Endovascular Trophoblasts in Decidual Vasculopathy with Implication for Preeclampsia and Other Pregnancy Complications. Fetal Pediatr. Pathol. 2020, 1–20. [Google Scholar] [CrossRef]
- Zhang, P. Decidual Vasculopathy in Preeclampsia and Spiral Artery Remodeling Revisited: Shallow Invasion versus Failure of Involution. Am. J. Perinatol. Rep. 2018, 8, e241–e246. [Google Scholar] [CrossRef] [Green Version]
- Craven, C.M.; Morgan, T.; Ward, K. Decidual spiral artery remodelling begins before cellular interaction with cytotrophoblasts. Placenta 1998, 19, 241–252. [Google Scholar] [CrossRef]
- Hastie, N.D. Wilms’ tumour 1 (WT1) in development, homeostasis and disease. Development 2017, 144, 2862–2872. [Google Scholar] [CrossRef] [Green Version]
- Call, K.M.; Glaser, T.; Ito, C.Y.; Buckler, A.J.; Pelletier, J.; Haber, D.A.; Rose, E.A.; Kral, A.; Yeger, H.; Lewis, W.H.; et al. Isolation and characterization of a zinc finger polypeptide gene at the human chromosome 11 Wilms’ tumor locus. Cell 1990, 60, 509–520. [Google Scholar] [CrossRef]
- Haber, D.A.; Buckler, A.J.; Glaser, T.; Call, K.M.; Pelletier, J.; Sohn, R.L.; Douglass, E.C.; Housman, D.E. An internal deletion within an 11p13 zinc finger gene contributes to the development of Wilms’ tumor. Cell 1990, 61, 1257–1269. [Google Scholar]
- Haber, D.A.; Housman, D.E. The genetics of Wilms’ tumor. Adv. Cancer Res. 1992, 59, 41–68. [Google Scholar]
- Haber, D.A.; Buckler, A.J. WT1: A novel tumor suppressor gene inactivated in Wilms’ tumor. New Biol. 1992, 4, 97–106. [Google Scholar]
- Van den Heuvel-Eibrink, M.M. Wilms Tumor. 2016. Available online: https://www.ncbi.nlm.nih.gov/books/NBK373360/?term=Wilms%20tumor (accessed on 30 April 2020).
- Buckler, A.J.; Pelletier, J.; Haber, D.A.; Glaser, T.; Housman, D.E. Isolation, characterization, and expression of the murine Wilms’ tumor gene (WT1) during kidney development. Mol. Cell. Boil. 1991, 11, 1707–1712. [Google Scholar] [CrossRef] [Green Version]
- Pelletier, J.; Schalling, M.; Buckler, A.J.; Rogers, A.; Haber, D.A.; Housman, D. Expression of the Wilms’ tumor gene WT1 in the murine urogenital system. Genes Dev. 1991, 5, 1345–1356. [Google Scholar] [CrossRef] [Green Version]
- Patek, C.E.; Little, M.H.; Fleming, S.; Miles, C.; Charlieu, J.-P.; Clarke, A.; Miyagawa, K.; Christie, S.; Doig, J.; Harrison, D.J.; et al. A zinc finger truncation of murine WT1 results in the characteristic urogenital abnormalities of Denys-Drash syndrome. Proc. Natl. Acad. Sci. USA 1999, 96, 2931–2936. [Google Scholar] [CrossRef] [Green Version]
- Francke, U.; Holmes, L.; Atkins, L.; Riccardi, V. Aniridia-Wilms’ tumor association: Evidence for specific deletion of 11p13. Cytogenet. Genome Res. 1979, 24, 185–192. [Google Scholar] [CrossRef]
- Chau, Y.-Y.; Brownstein, D.; Mjoseng, H.K.; Lee, W.-C.; Buza-Vidas, N.; Nerlov, C.; Jacobsen, S.E.W.; Perry, P.; Berry, R.; Thornburn, A.; et al. Acute multiple organ failure in adult mice deleted for the developmental regulator WT1. PLoS Genet. 2011, 7, e1002404. [Google Scholar] [CrossRef] [Green Version]
- Kaspar, H.G.; Crum, C.P. The utility of immunohistochemistry in the differential diagnosis of gynecologic disorders. Arch. Pathol. Lab. Med. 2015, 139, 39–54. [Google Scholar] [CrossRef] [Green Version]
- Husain, A.N.; Colby, T.V.; Ordóñez, N.G.; Allen, T.C.; Attanoos, R.L.; Beasley, M.B.; Butnor, K.J.; Chirieac, L.R.; Churg, A.M.; Dacic, S.; et al. Guidelines for Pathologic Diagnosis of Malignant Mesothelioma 2017 Update of the Consensus Statement From the International Mesothelioma Interest Group. Arch. Pathol. Lab. Med. 2018, 142, 89–108. [Google Scholar] [CrossRef] [Green Version]
- Khong, T.Y.; Mooney, E.E.; Ariel, I.; Balmus, N.C.M.; Boyd, T.K.; Brundler, M.-A.; Derricott, H.; Evans, M.J.; Faye-Petersen, O.M.; Gillan, J.E.; et al. Sampling and Definitions of Placental Lesions: Amsterdam Placental Workshop Group Consensus Statement. Arch. Pathol. Lab. Med. 2016, 140, 698–713. [Google Scholar] [CrossRef] [Green Version]
- Gebrane-Younes, J.; Hoang, N.M.; Orcel, L. Ultrastructure of human umbilical vessels: A possible role in amniotic fluid formation? Placenta 1986, 7, 173–185. [Google Scholar] [CrossRef]
- Anthony, F.W.; Mukhtar, D.D.; Pickett, M.A.; Cameron, I.T. Progesterone up-regulates WT1 mRNA and protein, and alters the relative expression of WT1 transcripts in cultured endometrial stromal cells. J. Soc. Gynecol. Investig. 2003, 10, 509–516. [Google Scholar] [CrossRef]
- Werner, H.; Rauscher, F.J.; Sukhatme, V.P.; A Drummond, I.; Roberts, C.T.; Leroith, D. Transcriptional repression of the insulin-like growth factor I receptor (IGF-I-R) gene by the tumor suppressor WT1 involves binding to sequences both upstream and downstream of the IGF-I-R gene transcription start site. J. Boil. Chem. 1994, 269, 12577–12582. [Google Scholar]
- Roberts, C.T. Control of insulin-like growth factor (IGF) action by regulation of IGF-I receptor expression. Endocr. J. 1996, 43, S49–S55. [Google Scholar] [CrossRef] [Green Version]
- Moffett, P.; Bruening, W.; Nakagama, H.; Bardeesy, N.A.; Housman, D.; Housman, D.E.; Pelletier, J. Antagonism of WT1 activity by protein self-association. Proc. Natl. Acad. Sci. USA 1995, 92, 11105–11109. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Huang, K.; Palmer, R.; Truong, V.B.; Herzlinger, D.; A Kolquist, K.; Wong, J.; Paulding, C.; Yoon, S.K.; Gerald, W.; et al. The Wilms tumor suppressor WT1 encodes a transcriptional activator of amphiregulin. Cell 1999, 98, 663–673. [Google Scholar] [CrossRef] [Green Version]
- Ciarloni, L.; Mallepell, S.; Brisken, C. Amphiregulin is an essential mediator of estrogen receptor alpha function in mammary gland development. Proc. Natl. Acad. Sci. USA 2007, 104, 5455–5460. [Google Scholar] [CrossRef] [Green Version]
- Pronier, E.; Bowman, R.L.; Ahn, J.; Glass, J.; Kandoth, C.; Merlinsky, T.R.; Whitfield, J.T.; Durham, B.H.; Gruet, A.; Somasundara, A.V.H.; et al. Genetic and epigenetic evolution as a contributor to WT1-mutant leukemogenesis. Blood 2018, 132, 1265–1278. [Google Scholar] [CrossRef]
- Moser, G.; Windsperger, K.; Pollheimer, J.; Lopes, S.C.D.S.; Huppertz, B. Human trophoblast invasion: New and unexpected routes and functions. Histochem. Cell Boil. 2018, 150, 361–370. [Google Scholar] [CrossRef] [Green Version]
- Scholz, H.; Kirschner, K.M. Oxygen-Dependent Gene Expression in Development and Cancer: Lessons Learned from the Wilms’ Tumor Gene, WT1. Front. Mol. Neurosci 2011, 4, 4. [Google Scholar] [CrossRef] [Green Version]
- Small, T.W.; Bolender, Z.; Bueno, C.; O’Neil, C.; Nong, Z.; Rushlow, W.; Rajakumar, N.; Kandel, C.; Strong, J.; Madrenas, J.; et al. Wilms’ tumor 1-associating protein regulates the proliferation of vascular smooth muscle cells. Circ. Res. 2006, 99, 1338–1346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Small, T.W.; Penalva, L.O.; Pickering, J.G. Vascular biology and the sex of flies: Regulation of vascular smooth muscle cell proliferation by wilms’ tumor 1-associating protein. Trends Cardiovasc. Med. 2007, 17, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Estrada, O.M.M.; Lettice, L.A.; Essafi, A.; Guadix, J.A.; Slight, J.; Velecela, V.; Hall, E.; Reichmann, J.; Devenney, P.S.; Hohenstein, P.; et al. WT1 is required for cardiovascular progenitor cell formation through transcriptional control of Snail and E-cadherin. Nat. Genet. 2010, 42, 89–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chau, Y.Y.; Hastie, N.D. The role of WT1 in regulating mesenchyme in cancer, development, and tissue homeostasis. Trends Genet. 2012, 28, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Chau, Y.-Y.; Bandiera, R.; Serrels, A.; Estrada, O.M.M.; Qing, W.; Lee, M.; Slight, J.; Thornburn, A.; Berry, R.; McHaffie, S.; et al. Visceral and subcutaneous fat have different origins and evidence supports a mesothelial source. Nat. Cell Biol. 2014, 16, 367–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chau, Y.Y.; Hastie, N. WT1, the mesothelium and the origins and heterogeneity of visceral fat progenitors. Adipocyte 2015, 4, 217–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuzaki, S.; Canis, M.; Darcha, C.; Déchelotte, P.J.; Pouly, J.-L.; Mage, G. Expression of WT1 is down-regulated in eutopic endometrium obtained during the midsecretory phase from patients with endometriosis. Fertil. Steril. 2006, 86, 554–558. [Google Scholar] [CrossRef]
- Sbracia, M.; Scarpellini, F.; Zupi, E.; Manna, C.; Marconi, D.; Romanini, C.; Alo, P.; Tondo, U.; Grasso, J. Differential expression of IGF-I and IGF-II in eutopic and ectopic endometria of women with endometriosis and in women without endometriosis. Am. J. Reprod. Immunol. 1997, 37, 326–329. [Google Scholar] [CrossRef]
- Gurgan, T.; Bukulmez, O.; Yarali, H.; Tanir, M.; Akyildiz, S. Serum and peritoneal fluid levels of IGF I and II and insulinlike growth binding protein-3 in endometriosis. J. Reprod. Med. 1999, 44, 450–454. [Google Scholar]
- Reeve, A.E.; Eccles, M.R.; Wilkins, R.J.; Bell, G.I.; Millow, L.J. Expression of insulin-like growth factor-II transcripts in Wilms’ tumour. Nature 1985, 317, 258–260. [Google Scholar] [CrossRef]

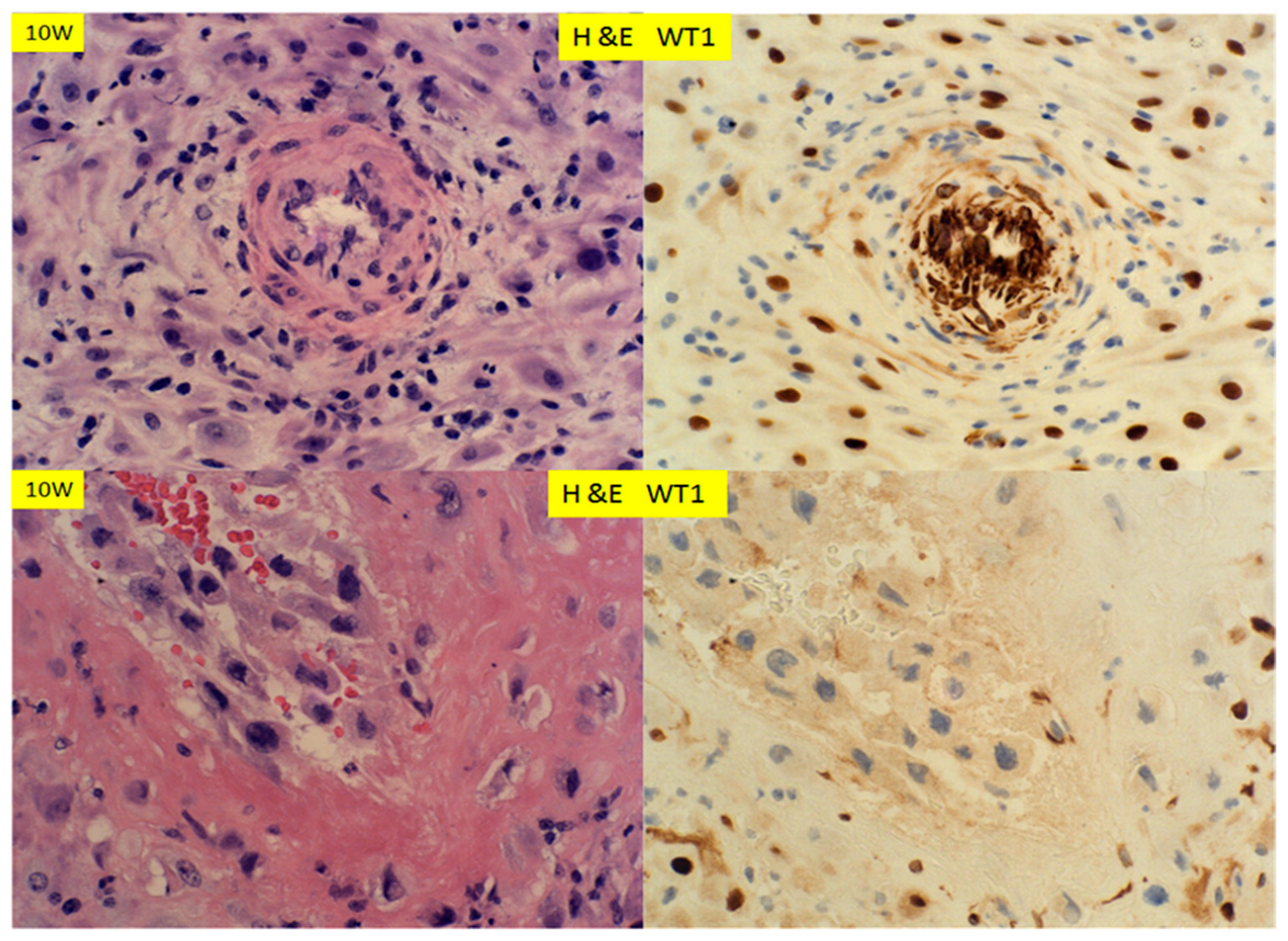
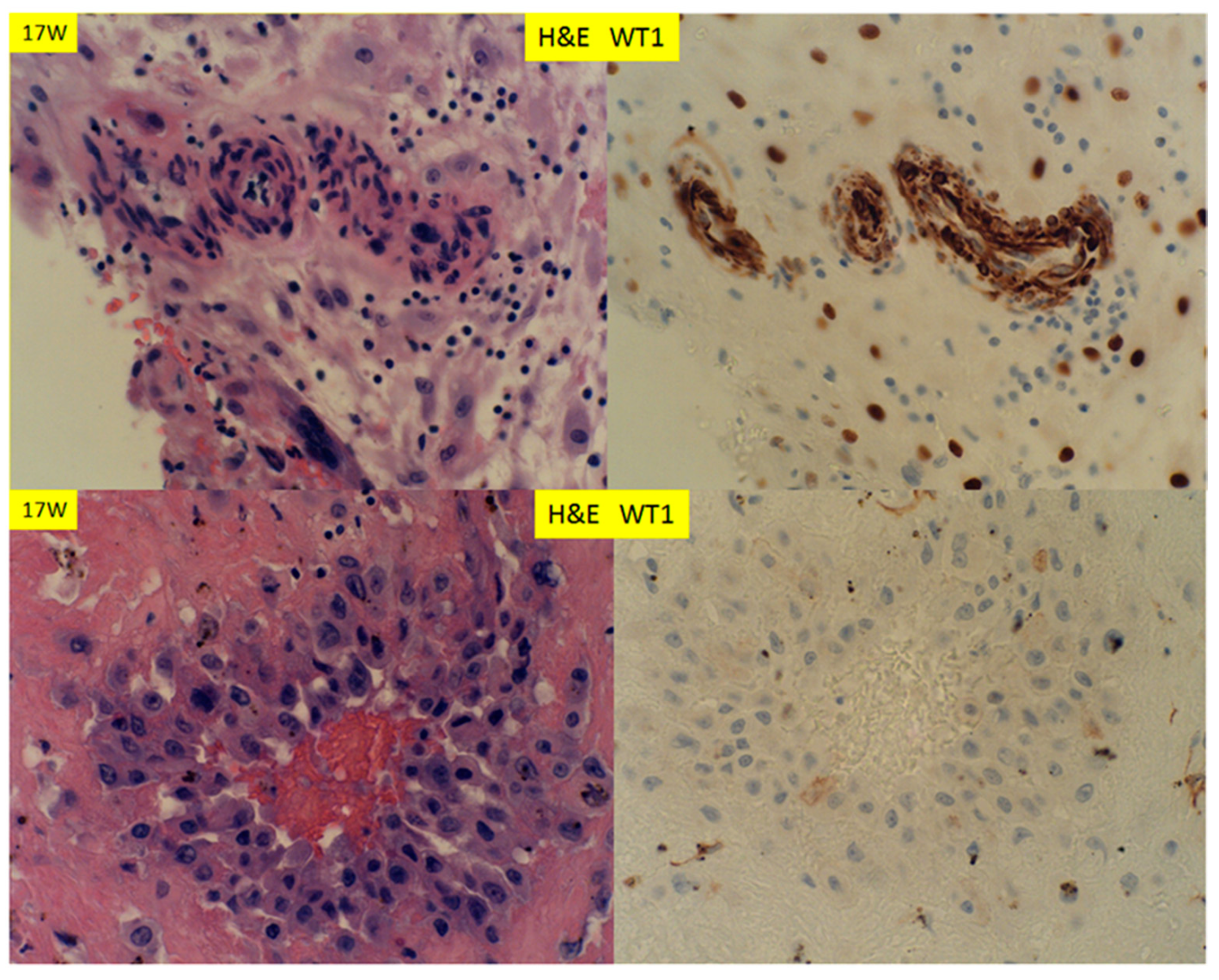
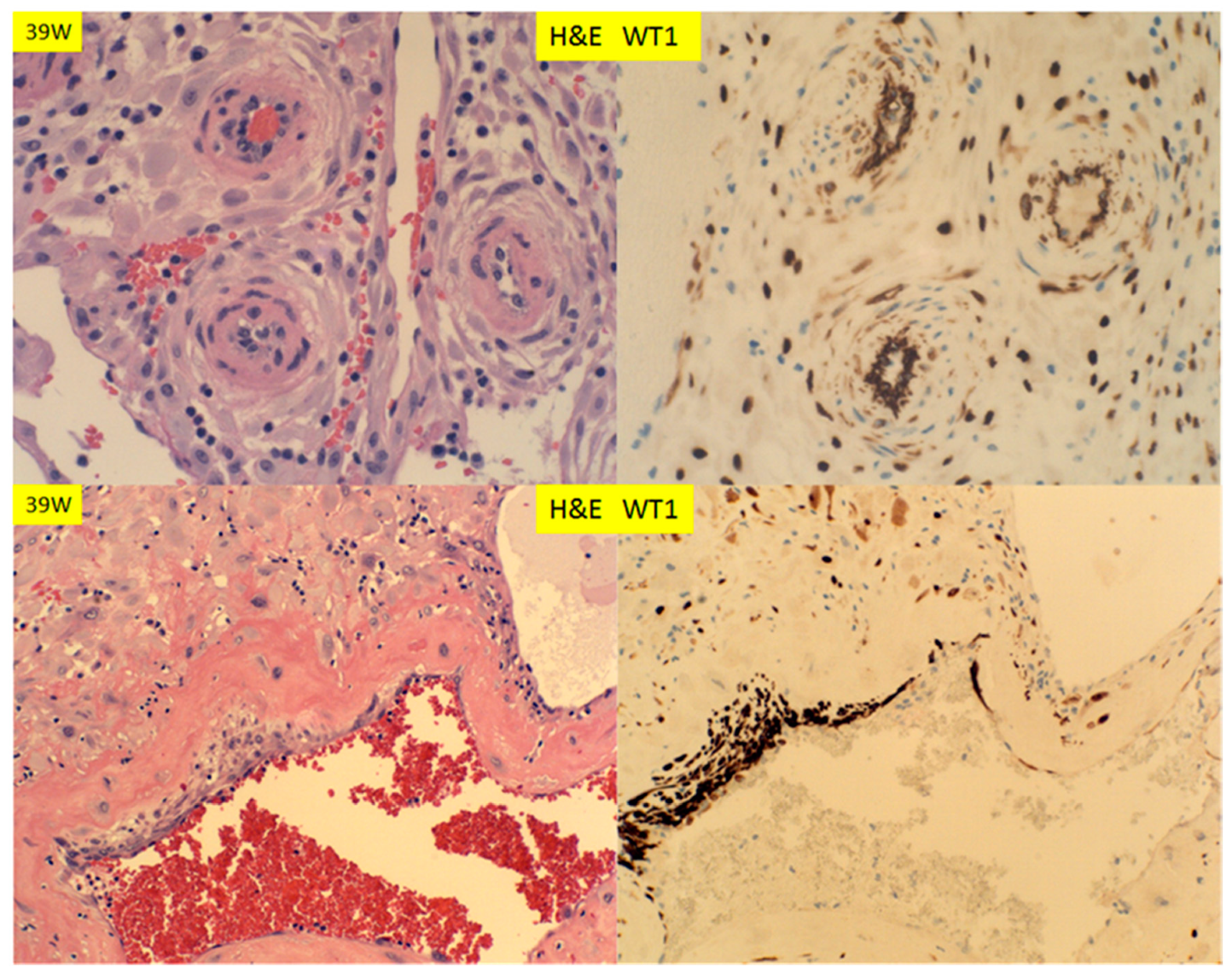
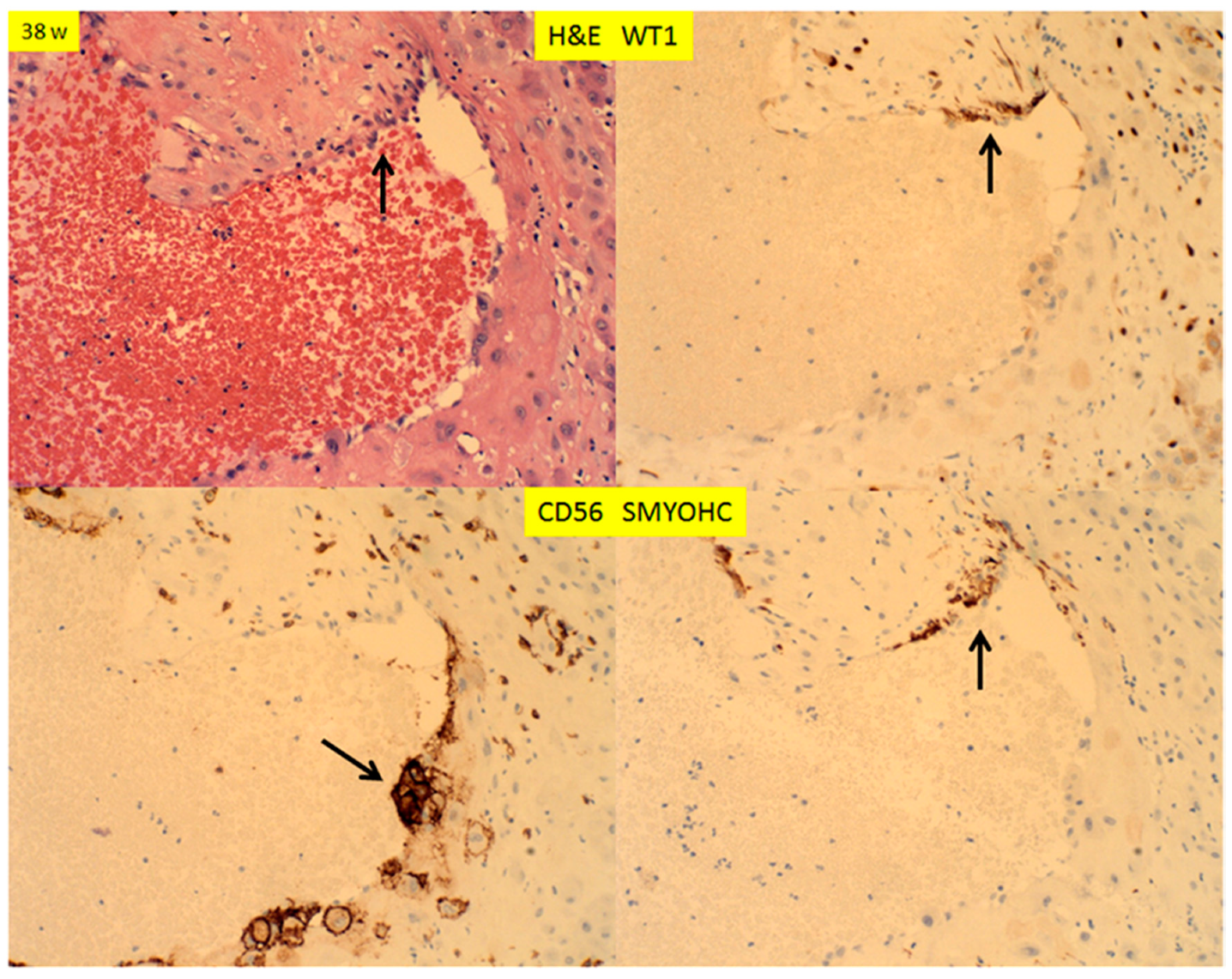
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, P. Expression of Wilm’s Tumor Gene (WT1) in Endometrium with Potential Link to Gestational Vascular Transformation. Reprod. Med. 2020, 1, 17-31. https://doi.org/10.3390/reprodmed1010003
Zhang P. Expression of Wilm’s Tumor Gene (WT1) in Endometrium with Potential Link to Gestational Vascular Transformation. Reproductive Medicine. 2020; 1(1):17-31. https://doi.org/10.3390/reprodmed1010003
Chicago/Turabian StyleZhang, Peilin. 2020. "Expression of Wilm’s Tumor Gene (WT1) in Endometrium with Potential Link to Gestational Vascular Transformation" Reproductive Medicine 1, no. 1: 17-31. https://doi.org/10.3390/reprodmed1010003
APA StyleZhang, P. (2020). Expression of Wilm’s Tumor Gene (WT1) in Endometrium with Potential Link to Gestational Vascular Transformation. Reproductive Medicine, 1(1), 17-31. https://doi.org/10.3390/reprodmed1010003