
Morphological Insights into Echinometra lucunter Spines Reveal Cellular Sources of Bioactive Molecules

 , , , and
, , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Light Microscopy
2.2. Transmission Electron Microscopy (TEM)
2.3. Scanning Electron Microscopy (SEM)
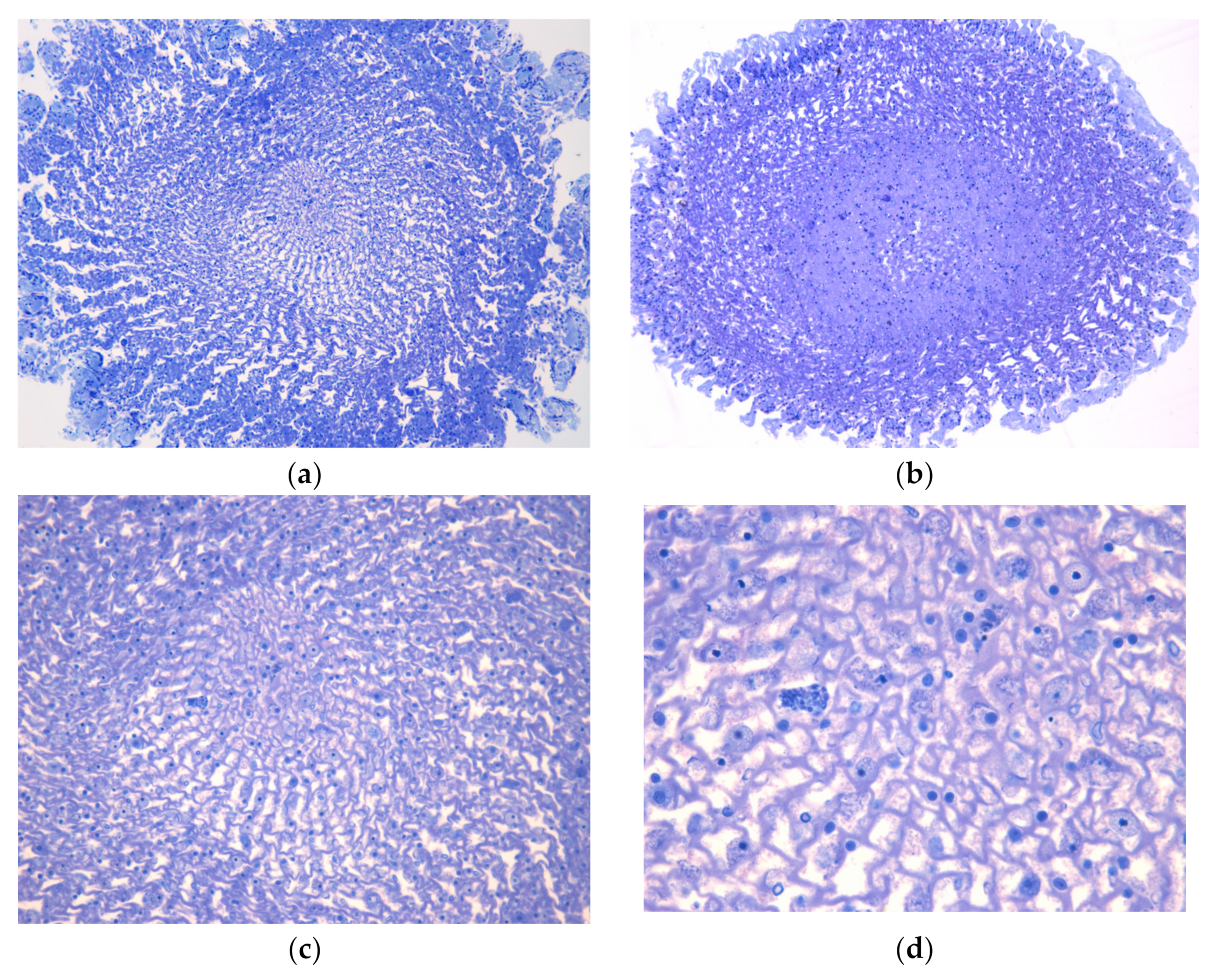
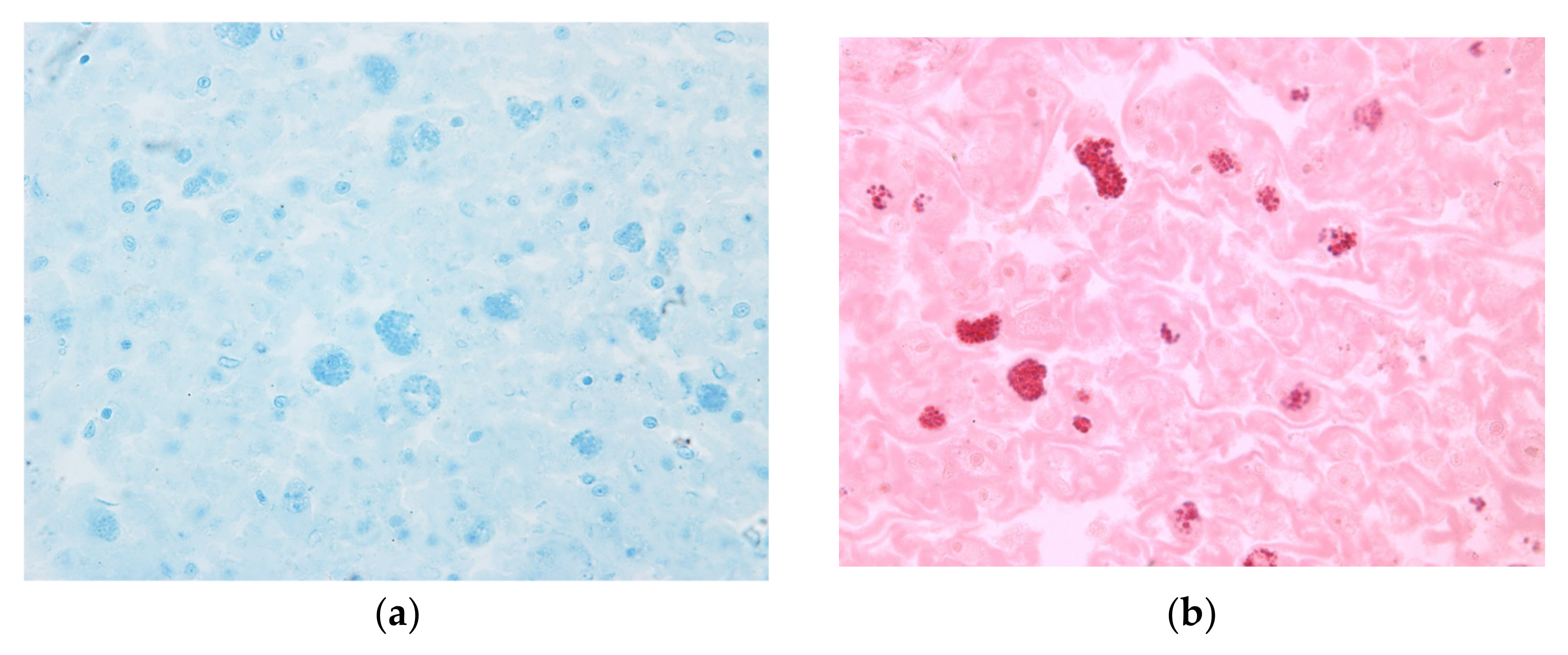
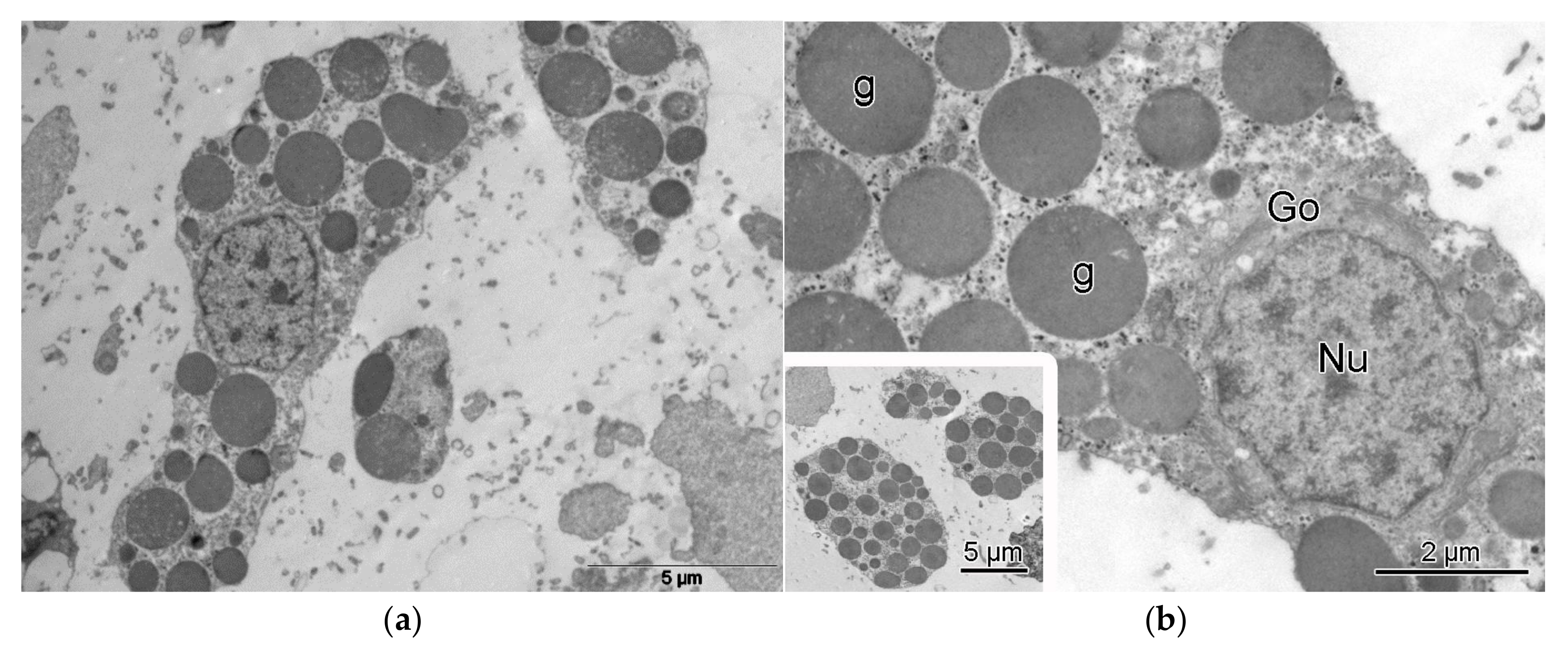
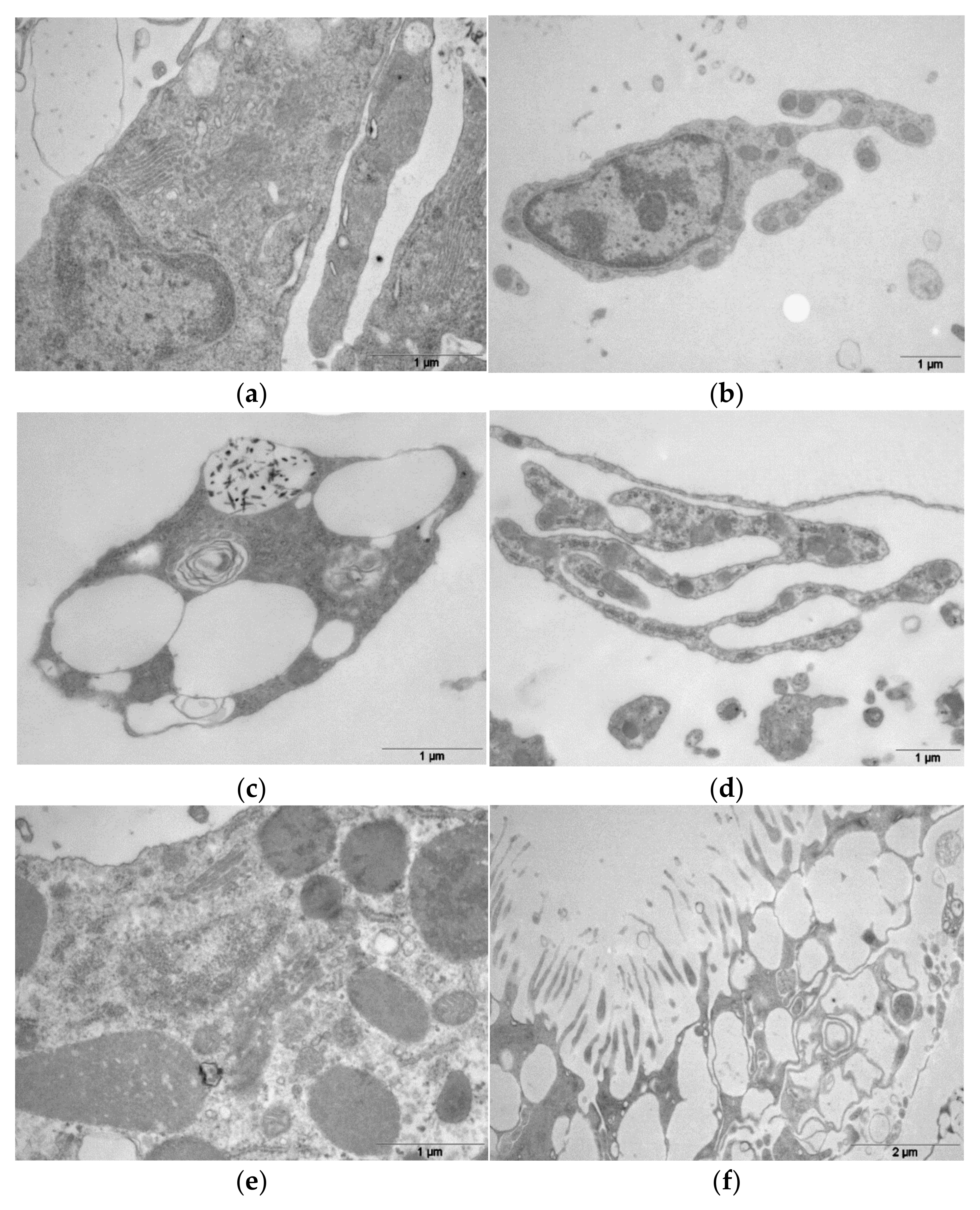
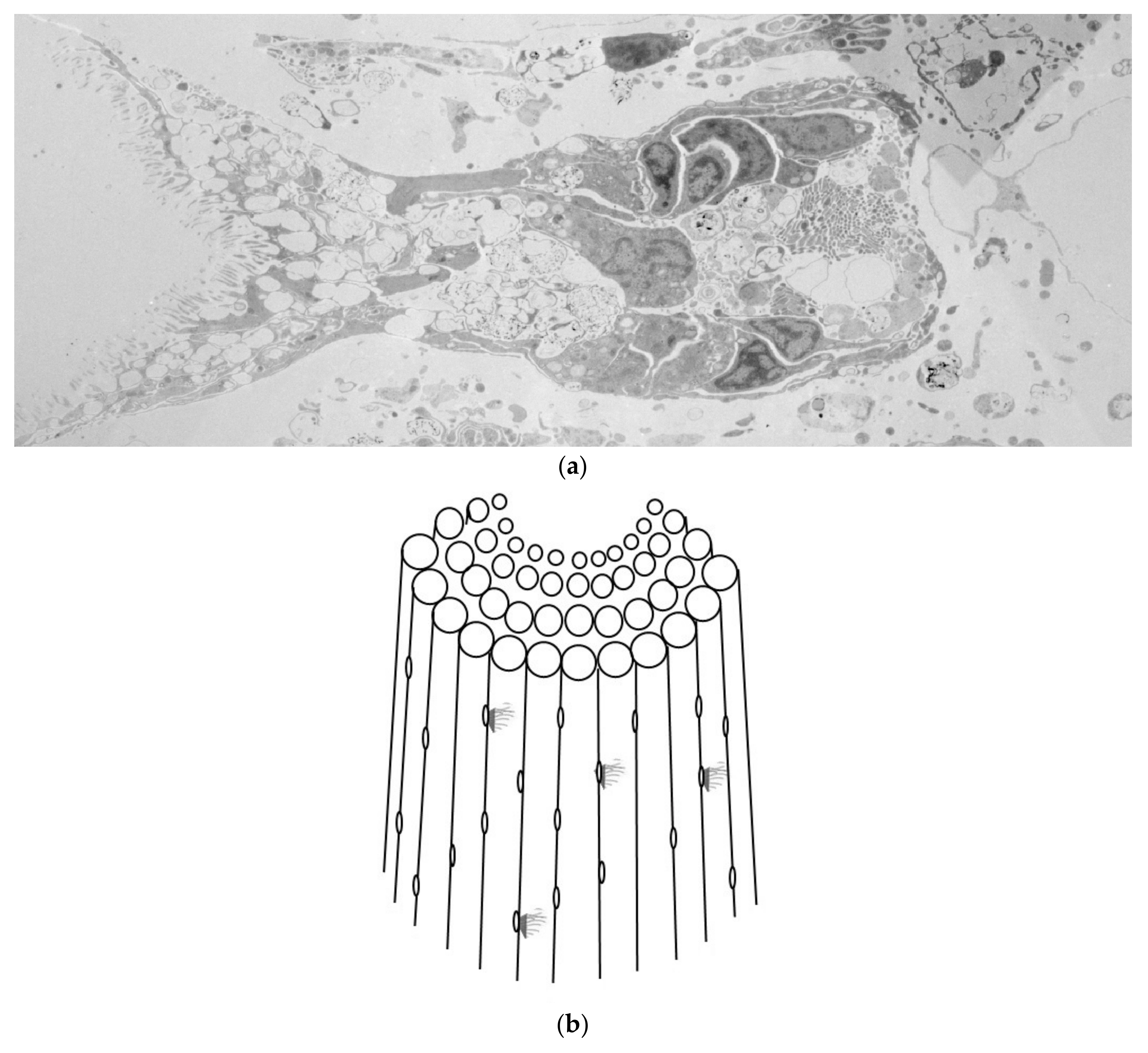
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Smith, A.B.; Peterson, K.J.; Wray, G.; Littlewood, D.T.J. From bilateral symmetry to pentaradiality. In Assembling the Tree of Life; Cracraft, J., Donoghue, M.J., Eds.; Oxford University Press: New York, NY, USA, 2004; pp. 365–383. [Google Scholar]
- Swalla, B.J.; Smith, A.B. Deciphering deuterostome phylogeny: Molecular, morphological and palaeontological perspectives. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2008, 363, 1557–1568. [Google Scholar] [CrossRef] [PubMed]
- Cloney, R.A.; Brocco, S.L. Chromatophore organs, reflector cells, iridocytes and leucophores in cephalopods. Am. Zool. 1983, 23, 581–592. [Google Scholar] [CrossRef]
- Holland, N.D. Epidermal cells. In Biology of the Integument—Invertebrates; Bereiter-Hahn, J., Matoltsy, A.G., Richards, K.S., Eds.; Springer: Berlin/Heidelberg, Germany, 1984; pp. 756–774. [Google Scholar]
- Hidaka, M.; Takahashi, K. Fine structure and mechanical properties of the catch apparatus of the sea urchin spine, a collagenous connective tissue with muscle-like holding capacity. J. Exp. Biol. 1983, 103, 1–14. [Google Scholar] [CrossRef]
- Dubois, P.; Ameye, L. Regeneration of spines and pedicellariae in Echinoderms: A review. Microsc. Res. Tech. 2001, 55, 427–437. [Google Scholar] [CrossRef]
- Moureaux, C.; Pérez-Huerta, A.; Compère, P.; Zhu, W.; Leloup, T.; Cusack, M.; Dubois, P. Structure, composition and mechanical relations to function in sea urchin spine. J. Struct. Biol. 2010, 170, 41–49. [Google Scholar] [CrossRef]
- Berman, A.; Addadi, L.; Kvick, A.; Leiserowitz, L.; Nelson, M.; Weiner, S. Intercalation of sea urchin proteins in calcite: Study of a crystalline composite material. Science 1990, 250, 664–667. [Google Scholar] [CrossRef]
- Weiner, S. Organic matrix-like macromolecules associated with the mineral phase of sea urchin skeletal plates and teeth. J. Exp. Zool. 1985, 234, 7–15. [Google Scholar] [CrossRef]
- Kanold, J.M.; Guichard, N.; Immel, F.; Plasseraud, L.; Corneillat, M.; Alcaraz, G.; Brümmer, F.; Marin, F. Spine and test skeletal matrices of the Mediterranean sea urchin Arbacia lixula—A comparative characterization of their sugar signature. FEBS J. 2015, 282, 1891–18905. [Google Scholar] [CrossRef]
- Morocco, A. Sea urchin envenomation. Clin. Toxicol. 2005, 43, 119–120. [Google Scholar] [CrossRef]
- Sciani, J.M.; Zychar, B.C.; Gonçalves, L.R.; Nogueira, T.O.; Giorgi, R.; Pimenta, D.C. Pro-inflammatory effects of the aqueous extract of Echinometra lucunter sea urchin spines. Exp. Biol. Med. 2011, 236, 277–280. [Google Scholar] [CrossRef]
- Sciani, J.M.; Zychar, B.C.; Gonçalves, L.R.; Giorgi, R.; Nogueira, T.; Pimenta, D.C. Preliminary molecular characterization of a proinflammatory and nociceptive molecule from the Echinometra lucunter spines extracts. J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 43. [Google Scholar] [CrossRef] [PubMed]
- Rosseto, A.L.; Mora, J.M.; Haddad Junior, V. Sea urchin granuloma. Rev. Inst. Med. Trop. São Paulo 2006, 48, 303–306. [Google Scholar] [CrossRef] [PubMed]
- Lima, E.J.B.; Gomes, P.B.; Souza, J.R.B. Reproductive biology of Echinometra lucunter (Echinodermata: Echinoidea) in a northeast Brazilian sandstone reef. Na. Acad. Bras. Cienc. 2009, 81, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Sciani, J.M.; Antoniazzi, M.M.; Neves, A.C.; Pimenta, D.C. Cathepsin B/X is secreted by Echinometra lucunter sea urchin spines, a structure rich in granular cells and toxins. J. Venom. Anim. Toxins Incl. Trop. Dis. 2013, 19, 33. [Google Scholar] [CrossRef]
- Sciani, J.M.; Emerenciano, A.K.; Cunha da Silva, J.R.; Pimenta, D.C. Initial peptidomic profiling of Brazilian sea urchins: Arbacia lixula, Lytechinus variegatus and Echinometra lucunter. J. Venom. Anim. Toxins Incl. Trop. Dis. 2016, 22, 17. [Google Scholar] [CrossRef]
- da Silva, A.G.; Alves, M.D.M.; da Cunha, A.A.; Caires, G.A.; Kerkis, I.; Vigerelli, H.; Sciani, J.M. Echinometra lucunter molecules reduce Aβ42-induced neurotoxicity in SH-SY5Y neuron-like cells: Effects on disaggregation and oxidative stress. J. Venom. Anim. Toxins Incl. Trop. Dis. 2023, 29, e20230031. [Google Scholar] [CrossRef]
- Ebert, T.A. Growth and Repair of Spines in the Sea Urchin Strongylocentrotus purpuratus (Stimpson). Biol. Bull. 1967, 133, 141–149. [Google Scholar] [CrossRef]
- Donovan, S.K. The internal morphology of primary spines of extant regular echinoids in the tropical western Atlantic: A SEM atlas. Swiss J. Palaeontol. 2018, 137, 363–377. [Google Scholar] [CrossRef]
- Smith, D.S.; Brink, D.; Del Castillo, J. Nerves in the spine of a sea urchin: A neglected division of the echinoderm nervous system. Proc. Natl. Acad. Sci. USA 1985, 82, 1555–1557. [Google Scholar] [CrossRef]
- Hebert, E.; Silvia, M.; Wessel, G.M. Structural and molecular distinctions of primary and secondary spines in the sea urchin Lytechinus variegatus. Sci. Rep. 2024, 14, 28525. [Google Scholar] [CrossRef]
- Borges, J.C.S.; Jensch-Junior, B.E.; Garrido, P.A.G.; Cepellos, M.B.B.; Mangiaterra, D.; Silva, J.R.M.C. Phagocytic Amoebocyte Sub Populations in the Perivisceral Coelom of the Sea Urchin Lytechinus variegatus (Lamarck, 1816). J. Exp. Zool. 2005, 303, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Arizza, V.; Giaramita, F.T.; Parrinello, D.; Cammarata, M.; Parrinello, N. Cell cooperation in coelomocyte cytotoxic activity of Paracentrotus lividus coelomocytes. Comp. Biochem. Physiol. Part A 2007, 147, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.R.M.C. Immunology in sea urchins. In Sea Urchins: Biology and Ecology; Lawrence, J.M., Ed.; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Heatfield, B.M.; Travis, D.F. Ultrastructural studies of regenerating spines of the sea urchin Strongylocentrotus purpuratus I. Cell types without spherules. J. Morphol. 1975, 145, 13–49. [Google Scholar] [CrossRef] [PubMed]
- Xing, K.; Yang, H.S.; Chen, M.Y. Morphological and ultrastructural characterization of the coelomocytes in Apostichopus japonicus. Aquat. Biol. 2008, 2, 85–92. [Google Scholar] [CrossRef]
- Bachmann, S.; Pohla, H.; Goldschmid, A. Phagocytes in the Axial Complex of the Sea Urchin, Sphaerechinus granularis. Cell Tissue Res. 1980, 213, 109–120. [Google Scholar] [CrossRef]
- Faria, M.T.; Silva, J.R.M.C. Innate immune response in the sea urchin Echinometra lucunter (Echinodermata). J. Invert. Pathol. 2008, 98, 58–62. [Google Scholar] [CrossRef]
- Gross, P.S.; Al-Sharif, W.A.; Clow, L.A.; Smith, L.C. Echinoderm immunity and the evolution of the complement system. Dev. Comp. Immunol. 1999, 23, 429–442. [Google Scholar] [CrossRef]
- Spurrell, M.; Oulhen, N.; Foster, S.; Perillo, M.; Wessel, G. Gene regulatory divergence amongst echinoderms underlies appearance of pigment cells in sea urchin development. Dev. Biol. 2023, 494, 13–25. [Google Scholar] [CrossRef]
- Perillo, M.; Oulhen, N.; Foster, S.; Spurrell, M.; Calestani, C.; Wessel, G. Regulation of dynamic pigment cell states at single-cell resolution. eLife 2020, 9, e60388. [Google Scholar] [CrossRef]
- Del Castillo, J.; Smith, D.S.; Vidal, A.M.; Sierra, C. Catch in the primary spines of the sea urchin Eucidaris tribuloides: A brief review and a new interpretation. Biol. Bull. 1995, 188, 120–127. [Google Scholar] [CrossRef]
- Albeck, S.; Weiner, S.; Addadi, L. Polysaccharides of intracrystalline glycoproteins modulate calcite crystal growth in vitro. Chem. A Eur. J. 1996, 2, 278–284. [Google Scholar] [CrossRef]
- MacKenzie, C.R.; Wilbanks, S.M.; McGrath, K.M. Superimposed effect of kinetics and echinoderm glycoproteins on hierarchical growth of calcium carbonate. J. Mat. Chem. 2004, 14, 1238–1244. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sciani, J.M.; Antoniazzi, M.M.; Jared, C.; Silva, J.R.M.C.d.; Pimenta, D.C. Morphological Insights into Echinometra lucunter Spines Reveal Cellular Sources of Bioactive Molecules. Oceans 2025, 6, 33. https://doi.org/10.3390/oceans6020033
Sciani JM, Antoniazzi MM, Jared C, Silva JRMCd, Pimenta DC. Morphological Insights into Echinometra lucunter Spines Reveal Cellular Sources of Bioactive Molecules. Oceans. 2025; 6(2):33. https://doi.org/10.3390/oceans6020033
Chicago/Turabian StyleSciani, Juliana Mozer, Marta Maria Antoniazzi, Carlos Jared, José Roberto Machado Cunha da Silva, and Daniel Carvalho Pimenta. 2025. "Morphological Insights into Echinometra lucunter Spines Reveal Cellular Sources of Bioactive Molecules" Oceans 6, no. 2: 33. https://doi.org/10.3390/oceans6020033
APA StyleSciani, J. M., Antoniazzi, M. M., Jared, C., Silva, J. R. M. C. d., & Pimenta, D. C. (2025). Morphological Insights into Echinometra lucunter Spines Reveal Cellular Sources of Bioactive Molecules. Oceans, 6(2), 33. https://doi.org/10.3390/oceans6020033