Abstract
Although traditional ryegrass pastures are still commonly used, they require intense management and input and do not perform well during the summer months. Multispecies systems have been recognised as more sustainable, needing less fertiliser and being tolerant to drought and heat. While data on monoculture system fat composition exist, information for multispecies is scarce. The present study compared the fatty acid (FA), carotenoid and fat-soluble composition of a multispecies system (consisting of perennial ryegrass, timothy, white clover, red clover, chicory and plantain) (MULTI) with two other conventional grazing systems (monoculture perennial ryegrass (PRG) and a binary mixture of perennial ryegrass and white clover (PRG+WC)) over the Irish late-summer grazing season of dairy cattle (July–September). The results showed that the three systems had similar levels of FAs (p > 0.05), with mean α-linolenic acid values of 20.00 mg/g in PRG, 18.51 mg/g in PRG+WC, and 17.90 mg/g in MULTI and mean linoleic acid values of 3.84 mg/g in PRG, 4.16 mg/g in PRG+WC, and 4.39 mg/g in MULTI. Fourfold and twofold increases in the concentrations of α-linolenic acid and linoleic acid, respectively, were observed throughout July–September. Variations in stearic acid concentrations were not significant (p > 0.05), and palmitic acid concentrations increased over time (p < 0.05). The average values of lutein (36.68 mg/kg in PRG, 31.26 mg/kg in PRG+WC, and 35.75 mg/kg in MULTI) and α-tocopherol (2.28 mg/kg in PRG, 1.49 mg/kg in PRG+WC, and 1.83 mg/kg in MULTI) were similar among the grazing systems (p > 0.05), and the average β-carotene value was 25% higher in monoculture ryegrass (140.74 mg/kg) than in the multispecies (102.51 mg/kg) (p < 0.05). Nonetheless, the levels of β-carotene found in the multispecies were still above the recommended intake required for ruminants. In conclusion, multispecies pastures could be included in dairy cattle production systems as a sustainable alternative to traditional pastures while also providing typically adequate sources of fats for animal diets.
1. Introduction
Dairy cattle consume about 6% fat in their diet, with 3–4% coming from feed and 2–3% from additional supplementation [1]. Fats participate in many important biological functions, such as phospholipid synthesis, cell proliferation and enzyme activity [2]. They play central roles in increasing energy density in the diet and improving animal performance and productivity [3]. Furthermore, some fats have bioactive properties; once absorbed, they can be directly transferred into milk and impact its nutritional value [4].
In Ireland, dairy cattle predominantly feed on fresh pastures, particularly during the summer months when the levels of dry matter are higher than the rest of the year [5]. The fat contents of pastures, including FAs, carotenoids and fat-soluble vitamins, vary with plant maturity and species composition [6]. The majority (>90%) of the FA content of pastures consists of palmitic, α-linolenic and linoleic acid. Palmitic acid has been shown to improve cattle performance and increase the production of milk [7], and α-linolenic and linoleic acid have been associated with increased fertility rates, immune response modulation and inflammation reduction [8]. Additionally, α-linolenic acid is directly related to the production of conjugated linoleic acid (CLA) isomers, which are health-promoting compounds found in milk; in particular, the cis-9, trans-12 CLA isomer has been linked with anti-obesity, anticarcinogenic, and antidiabetic properties [9]. Furthermore, α-linolenic acid is the precursor for very long-chain FAs (eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA)), which are associated with foetal development, cognitive functions and healthy aging [10]. Carotenoids and fat-soluble vitamins constitute the fat micronutrient fraction of pastures and are also associated with various health benefits. The antioxidant properties of the carotenoid lutein are linked to cancer prevention and immune function enhancement [11]. Studies on ruminants have also shown the ability of lutein to decrease somatic cell counts in milk [12]. Additionally, β-carotene and tocopherols are powerful antioxidants that can protect cells from oxidative damage and reduce the risk of intramammary infection and mastitis [13,14].
Traditional monoculture ryegrass pastures are rich in FAs, carotenoids and fat-soluble vitamins [15,16]. However, monocultures may not be suitable for sustainable environmental strategies, as they require intense management and inputs (e.g., fertilisers and pesticides). In order to decrease fertiliser requirements, white clover can be added as part of a ryegrass–clover binary mixture, but this type of mixture does not perform well in the summer, often leading to reduced dry matter (DM) intake and low animal performance [17].
Multispecies pastures consisting of different forages (e.g., grass, legume forages, and herbs) have been recognised as more sustainable, as they can increase biodiversity, improve soil function and reduce fertiliser requirements [18]. Furthermore, multispecies are tolerant to drought and heat, and they can produce more DM than perennial ryegrass–clover cultures [19]. Nevertheless, information on the fat composition of multispecies is scarce, and potential changes in concentrations during the summer grazing season still need to be assessed.
The present study investigated and compared the FA, carotenoid and fat-soluble vitamin composition of multispecies (MULTI) (consisting of perennial ryegrass (Lolium perenne), timothy (Phleum pratense), white clover (Trifolium repens), red clover (Trifolium pratense), chicory (Chicorium intybus) and plantain (Plantago lanceolata)) with two other conventional grazing systems (i.e., monoculture perennial ryegrass (PRG) and a binary mixture of perennial ryegrass and white clover (PRG+WC)) over the Irish late-summer grazing season (July–September). FAs were profiled using gas chromatography–triple-quadruple–mass spectrometry (GC–QqQ–MS), and carotenoids and vitamins were assessed using liquid chromatography–electrospray ionisation–triple-quadruple–mass spectrometry (LC–ESI–QqQ–MS) in the multiple reaction monitoring (MRM) mode.
An improved understanding of the fat composition in multispecies pastures may help farmers choose feeding strategies that are not only beneficial to the environment but also rich in nutrients that could potentially enhance dairy cattle health and productivity.
2. Materials and Methods
2.1. Chemicals and Reagents
HPLC grade (≥99.9) methanol, pentane, and hexane and LCMS grade (≥99.9) methanol were purchased from Fisher Scientific (Fisher Scientific Ireland Ltd., Dublin, Ireland). Butylated hydroxytoluene (BHT), potassium hydroxide, acetyl chloride, sodium chloride and sodium sulphate were purchased from Sigma-Aldrich (Sigma-Aldrich Ireland Ltd., Arklow, Ireland). A range of fatty acids, carotenoids and vitamin standards were also bought from Sigma-Aldrich.
2.2. Samples
The three grazing systems (PRG, PRG+WC and MULTI) were sowed in adjacent fields at University College Dublin, Lyons Research Farm (Co. Kildare, Ireland) and repeatedly harvested between July and September 2020 for three weeks per month (Figure 1). Four experimental paddocks were established, with each paddock comprising 2 ha of each grazing system. Initially, the site received 40 kg N ha−1, 25 kg P ha−1 and 80 kg K ha−1, which was followed by 205 kg N ha−1, 14 kg P ha−1 and 115 kg K ha−1 for PRG and 92 kg N ha−1, 18 kg P ha−1 and 115 kg K ha−1 for PRG+WC and MULTI. The seeding rates for individual species are shown in Table 1. All PRG, PRG+WC and MULTI swards were randomly collected to a residual of 5 cm throughout the experimental analysis still in their vegetative regrowth stage. Shortly after collection, the swards were washed with running water to remove any soil and dirt, frozen for 24 h, and lyophilized. The dried forages were milled using a blender and stored away from light until further analysis.
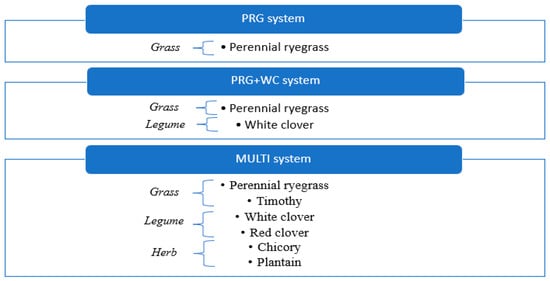
Figure 1.
Herbage composition of the PRG, PRG+WC and MULTI systems.

Table 1.
Proportion rate (%) of each forage species sown per grazing system (from July to September 2020).
2.3. Derivatisation of Fatty Acids to FAME
FAs were derivatised into free FA methyl esters (FAMEs) following the work of Brunton et al. [20] and using microwave-assisted FAME preparation. Briefly, 1 g of a sample and 100 µL of an internal standard (C23:0-methyl ester) were added to a 55 mL MARSXpress PFA reaction vessel containing a 10 mm stir bar. In the first step, 10 mL of 2.5% KOH in methanol was added into the vessel and placed in the MARS 6 Express 40 position Microwave Reaction System (CEM Corporation, Matthews, NC, USA). The system was heated to 130 °C over 4 min and held at 130 °C for a further 4 min. Subsequently, the vessel was removed from the microwave and cooled on ice for 5 min. In the second step, 15 mL of 5% acetyl chloride in methanol was added to the vessel and again placed in the microwave. The system was heated to 120 °C over 4 min and held at 120 °C for a further 2 min. Afterwards, the vessel was removed and placed in an ice bath for 5 min. For the extraction of FAMEs, 10 mL of pentane was added and mixed for 2 min. Finally, 20 mL of a saturated NaCl solution (30% w/v) was added, followed by mixing for a further 2 min. After 30 min, the top pentane layer was aliquoted in amber vials (1.5 mL, Agilent, Cork, Ireland) containing 0.2 g of sodium sulphate. Vials were stored at −18 °C before GC–MS analysis.
2.4. Identification and Quantification of Fatty Acids Using GC–MS
Mass spectrometry was used to identify and quantify the FAs present in the pastures. The instrument was composed of an Agilent Technologies GC 7890B coupled with an Agilent Technologies Triple Quadruple Mass Spectrometry detector. The separation was carried out with a CP-Sil 88 column ((100 m × 0.25 mm 0.20 mm) Agilent Technologies, Cork, Ireland)), the carrier gas was helium, and the split ratio was set at 15:1. The injection volume was 1 µL, with an inlet temperature of 280 °C. The oven was programmed to 80 °C, heated to 220 °C at a rate of 3.5 °C/min and held for 5 min. Subsequently, it was heated to 225 °C at a rate of 2 °C/min and held for 22.5 min (the overall run was 70 min). The mass spectrometer was in the positive mode and scanned 29 to 550 m/z every 0.3 s. The ion source was set at 230 °C and spectra were obtained via electron impact at 70 eV. The identification of compounds was performed using commercial standards and the NIST08 mass spectra library. Calibration curves were constructed from the detector responses of different standard concentrations (20 to 1000 µg/mL) using the equation for linear regression obtained from the calibration curves (R2 = 0.99).
2.5. Identification and Quantification of Carotenoids and Fat-Soluble Vitamins Using LC–MS
The method for carotenoid and fat-soluble vitamin analysis was based on a modified version of the method used by Gentili et al. [21]. Briefly, samples were pre-treated using overnight cold saponification. Dried samples (0.2 g) were added to a 15 mL propylene tube in addition to 3 mL of 0.1% ethanolic BHT, 0.2 mL of 50% aqueous KOH and 100 µL of an internal standard (menaquinone (vitamin K2)). The tube was flushed with nitrogen gas, promptly closed, and left shaking at 150 rpm overnight (approximately 15 h). Subsequently, the analytes of interest were extracted two times using 2 mL of hexane with 0.1% BHT. After the addition of each hexane aliquot, the sample was vortexed and centrifuged at 4500 rpm for 10 min. The hexane aliquots were pooled together and evaporated to dryness under a nitrogen flow. The dried extracts were reconstituted with 2 mL of methanol and stored at −18 °C before LC–MS analysis.
Mass spectrometry was used to identify and quantify the carotenoids and fat-soluble vitamins present in the grass samples. The instrument used was an Agilent Technologies 1290 Infinity series HPLC coupled with an Agilent Technologies 6470 triple quadruple with electrospray ionisation. A 5 µL sample was injected, and separation was carried out using a Poroshell 120 column (3.0 mm × 100 mm × 2.7 µm (Agilent Technologies, Cork, Ireland)) held at 40 °C. The mobile phase consisted of: (A) 0.1% formic acid in water and (B) 0.1% formic acid in methanol. Elution was performed at a flow rate of 1 mL/min using the following gradient: starting with 85% B; then reaching 90% B in 10 min, 92% in 4 min, and 100% in 6 min; and held for 7 min with post-time of 3 min.
The detection of eluted vitamins was performed using multiple reaction monitoring (MRM) with the following ion source: positive ion polarity, a gas flow of 5 L/min, a 45 psi nebulizer, a sheath gas temperature of 250 °C, and a sheath gas flow of 11 L/min. The parameters concerning the MRM mode are presented in Table 2. Data were analysed using the Agilent MassHunter Workstation Software—Qualitative Analysis (vers. 7). Calibration curves were constructed from the peak areas of different standard concentrations (0.6 to 5 µg/mL) using the equation for linear regression obtained from the calibration curves (R2 = 0.99).
Table 2.
LC–QqQ parameters of the carotenoids and fat-soluble vitamins selected in this study.
2.6. Statistical Analysis
All data are reported as the means ± standard deviations of triplicate determinations. Statistical analysis was performed with SPSS Statistical Software (vers. 28.0.0) using a one-way analysis of variance (ANOVA). Differences at p < 0.05 were considered statistically different. GC–MS and LC–MS data were analysed using Agilent MassHunter Workstation Software—Qualitative Analysis (vers. 10.0) and MassHunter Workstation Software—Quantitative Analysis (vers. 9.0).
3. Results
3.1. Identification and Quantification of Fatty Acids of the Grazing Systems
The FA profiles of the three grazing systems (PRG, PRG+WC and MULTI) were determined over the Irish late summer, typically between July and September (Table 3). Prior to GC analysis, FAs were derivatised into FAMEs; derivatisation was carried out to decrease the polarity of the free FAs that, under normal circumstances, would form hydrogen bonds with other molecules in the sample and negatively impact the chromatographic separation [22]. Individual FAMEs were identified and quantified using highly selective GC–QqQ–MS. A representative chromatogram of the predominant FAMEs identified in the pastures is shown in Figure 2. In accordance with previous studies [15,23], α-linolenic acid, linoleic acid, palmitic, and stearic acid ranged within 43–77%, 6–20%, 5–18% and 3–10% of the total FA composition in the forages, respectively. Small amounts of oleic acid (0.5–6%) were also found.
Table 3.
FAs composition (mg/g) of the grazing systems (PRG = perennial ryegrass; PRG+WC = perennial ryegrass and white clover; MULTI = perennial ryegrass, timothy, white clover, red clover, chicory, and plantain) estimated weekly (W1, W2, and W3) throughout the summer months (July, August, and September). Values are represented as the mean of three replicates. Significant differences (p < 0.05) within each system are indicated with letter superscripts. ND = not detected.
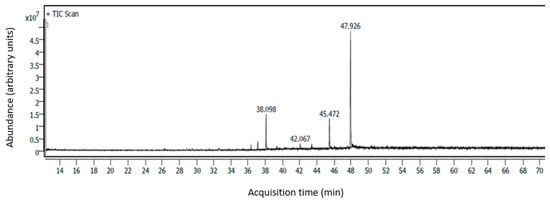
Figure 2.
CP Sil 88 column chromatogram of the major FAs found in the three grazing systems (38.098 min = palmitic acid; 42.067 min = stearic acid; 45.472 min = linoleic acid; 47.926 min = α-linolenic acid).
Overall, there were no significant differences in the α-linolenic acid and linoleic acid concentrations among the three grazing systems (p > 0.05). The mean α-linolenic acid values in PRG, PRG+WC and MULTI were found to be 20.00, 18.51 and 17.90 mg/g, respectively, and the mean linoleic acid values were 3.84, 4.16 and 4.39 mg/g, respectively. Variations in the concentrations of α-linolenic and linoleic acid were consistently observed among the three grazing systems throughout the season (p > 0.05) (Figure 3a,b). For example, α-linolenic acid had a fourfold concentration increase from July to September (9.61–35.94 mg/g in PRG, 8.73–35.15 mg/g in PRG+WC, and 8.81–34.25 mg/g in MULTI), and linoleic acid had up to twofold increases (3.34–5.21 mg/g in PRG, 3.57–6.32 mg/g in PRG+WC, and 2.74–6.10 mg/g in MULTI).
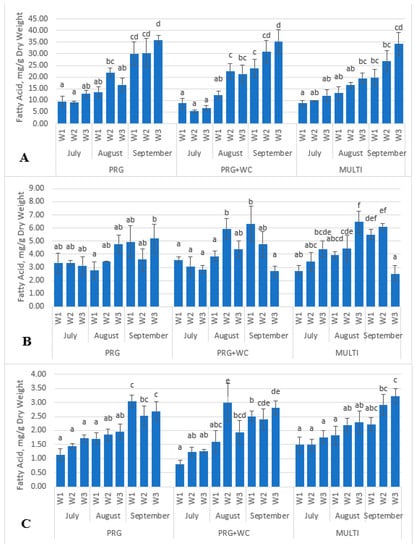
Figure 3.
Variations in the α-linolenic acid (A), linoleic acid (B) and palmitic acid (C) levels of the PRG (perennial ryegrass), PRG+WC (perennial ryegrass and white clover) and MULTI (perennial ryegrass, white clover, timothy, red clover, chicory, and plantain) systems estimated weekly (W1, W2, and W3) throughout the summer months (July, August, and September). Different superscript letters indicate significant difference (p < 0.05) within each system.
The three grazing systems were found to have similar levels of omega-3 PUFAs (59–62% of total FAs) and omega-6:3 PUFA ratios (0.23 in PRG, 0.29 in PRG+WC, and 0.27 in MULTI). Stearic acid concentrations did not differ between the three systems (p > 0.05). The mean palmitic acid value in PRG (2.34 mg/g) was higher than that in PRG+WC (1.95 mg/g) and MULTI (2.15 mg/g) (p < 0.05). Increasing levels of palmitic acid were observed throughout the season (p < 0.05) in the PRG (from 1.46 to 2.69 mg/g), PRG+WC (from 0.81 to 2.81 mg/g) and MULTI (from 1.49 to 3.22 mg/g) (Figure 3c). In addition to being present in MULTI, white clover, red clover and plantain were the species with the highest amounts of palmitic acid. Thus, the higher palmitic acid levels observed in the MULTI system could be attributed to the herbage composition of the system.
3.2. Identification and Quantification of Carotenoids and Fat-Soluble Vitamins in Grazing Systems
Although previous studies have investigated the benefits of feeding ruminants with diets high in carotenoids and fat-soluble vitamins [24,25], information on composition in multispecies systems is still scarce. In this study, the carotenoid (β-carotene and lutein) and vitamin (D2, D3, K1, α-tocopherol and γ-tocopherol) profiles of three grazing systems (PRG, PRG+WC and MULTI) were studied over the Irish late-summer grazing season using LC–ESI–QqQ–MS in the positive MRM acquisition mode (Table 4). The MRM mode is an accurate technique that monitors ions of the compounds of interest and provides more precise quantification at lower detection limits.
Table 4.
Carotenoid and fat-soluble vitamin composition (mg/kg) of the grazing systems (PRG = perennial ryegrass; PRG+WC = perennial ryegrass and white clover; MULTI = perennial ryegrass, timothy, white clover, red clover, chicory, and plantain) estimated weekly (W1, W2, and W3) throughout the summer months (July, August, and September). Values are represented as the mean of three replicates. Significant differences (p < 0.05) within each system are indicated with letter superscripts. ND = not detected.
Carotenoids were abundantly present in all three grazing systems, with the average β-carotene level being three times higher than that of lutein. PRG had the highest β-carotene concentrations throughout the grazing season (from 101.61 to 164.91 mg/kg DM), which were 25–28% higher than those in PRG+WC (from 86.07 to 130.03 mg/kg DM) and MULTI (from 58.11 to 151-0.83 mg/kg DM). Although changes in β-carotene concentrations during the season were not considered significant (p > 0.05), increasing levels were observed in PRG and PRG+WC while decreasing levels were observed in MULTI (Figure 4a). There were no significant differences (p < 0.05) in the seasonal lutein variations among PRG (from 24.55 to 54.15 mg/kg DM), PRG+WC (from 25.55 to 49.18 mg/kg DM) and MULTI (from 29.52 to 56.55 mg/kg DM). However, higher levels of lutein were found in the three systems early in the season (July), though these levels decreased by 20–30% in August (Figure 4b).
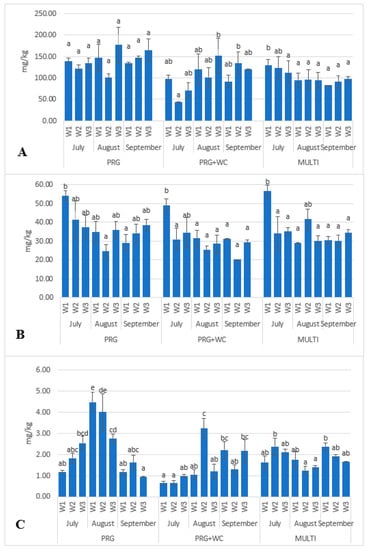
Figure 4.
Variations in the β-carotene (A), lutein (B) and α-tocopherol (C) levels of the PRG (perennial ryegrass), PRG+WC (perennial ryegrass and white clover) and MULTI (perennial ryegrass, white clover, timothy, red clover, chicory, and plantain) systems estimated weekly (W1, W2, and W3) throughout the summer months (July, August, and September). Different superscript letters indicate significant difference (p < 0.05) within each system.
PRG and PRG+WC were found to contain higher levels of β-carotene compared with MULTI. Indeed, the analysis of the individual forages (Table 5) revealed that perennial ryegrass contained the highest levels of β-carotene, up to almost two times more than the clover species and four times higher than the chicory species. A strong correlation (p < 0.01) was also found between the percentage of perennial ryegrass in the grazing systems (Table 1) and the concentration of β-carotene.
Table 5.
α-linolenic acid (mg/g), linoleic acid (mg/g), palmitic acid (mg/g), stearic acid (mg/g), lutein (mg/kg), β-carotene (mg/kg) and α-tocopherol (mg/kg) concentrations of the MULTI system components (perennial ryegrass, timothy, white clover, red clover, chicory, and plantain). Values are represented as the mean of five months of sampling. Significant differences (p < 0.05) within each forage ecotype are indicated with letter superscripts.
The concentrations of α-tocopherol throughout the grazing season were higher in PRG (from 1.14 to 4.01 mg/kg DM) and MULTI (from 0.83 to 2.38 mg/kg DM) than in PRG+WC (from 0.26 to 3.24 mg/kg DM). Spikes in α-tocopherol were observed in July in PRG (4.01 mg/kg DM) and PRG+WC (3.24 mg/kg DM), while levels remained consistent throughout the grazing season in MULTI (Figure 4c). This could be attributed to perennial ryegrass containing high levels of α-tocopherol and PRG and PRG+WC having higher percentages of perennial ryegrass compared with the MULTI system (Table 1).
4. Discussion
Changes in FA concentration over time might be dependent on several factors, such as weather, species, nitrogen availability, and the phenological stage of a plant. The frequent cutting regime performed in this study maintained the forages in a constant vegetative stage and stopped the flowering phase. Usually, the vegetative stage coincides with increased leaf production [26], and because most of the fats in plants are located in the chloroplasts of the leaves, increments in FA concentrations can be expected when a plant matures.
The mean linoleic acid value for MULTI in this study was found to be somewhat higher than that of PRG+WC and PRG, while the mean α-linolenic acid value of PRG was higher than that of PRG+WC and MULTI. The FA analysis of the individual forage species (Table 5) revealed that the highest concentrations of linoleic acid could be found in plantains, with levels up to twice as much as legumes and almost three times higher than grasses; since MULTI was the only pasture system containing plantain, higher concentrations of linoleic acid were expected. High levels of α-linolenic acid were found in red clover, with concentrations of up to almost twice as much as those of herbs and grasses; however, MULTI contained the least amount of α-linolenic acid despite being the only pasture system to have red clover. Since the lipase oxidation of α-linolenic acid is twice as fast as the lipase oxidation of linoleic acid [27], it is possible that the original high levels of α-linolenic acid in MULTI were depleted. Indeed, the pastures in this study did not undergo any preservation treatment (e.g., siling), so sufficient time might have been available for the lipase to degrade α-linolenic acid [28].
The omega-6:3 ratios found in PRG, PRG+WC and MULTI were much lower than most common feedstuffs added to ruminant rations (e.g., soybean and corn) [8]. In general, omega-6 PUFAs (linoleic acid) have pro-inflammatory, pro-allergic and pro-thrombotic properties, whereas omega-3 PUFAs (α-linolenic acid) are less pro-inflammatory and thrombotic. Although PUFAs undergo extensive hydrogenation in the rumen, feeding cattle with diets low in omega-6:3 could represent an opportunity to reduce the omega-6:3 ratio in milk and enhance its nutritional properties. Indeed, the human consumption of foods with low omega-6:3 ratios (4:1 or less) is considered a good strategy for CDV prevention [29].
In vitro studies have shown that α-linolenic acid can protect the maturation of oocytes under lipotoxic conditions and improve their development [30]. Indeed, the growth of oocytes in the ovarian follicles is a key factor in cattle fertility. Palmitic acid (42.66 mg/g) and stearic acid (19.23 mg/g) have been found to be toxic to ovarian follicles, affecting the quality of oocytes and their subsequent development [31]. The concentrations of palmitic and stearic acid reported in this study were below the toxic levels reported by Van Hoeck [31]. α-linolenic acid can also increase overall health of the reproduction system of cows, as it leads to more menstrual cycles and higher opportunities to expel contaminants from the uterus [32]. While the specific concentrations required to achieve these animal health benefits have not yet been established in in vivo studies, feeding multispecies pastures that have high levels of α-linolenic acid and low levels of palmitic and stearic acid might present sustainable, natural and economic opportunities for enhancing the reproductive health of animals.
A-linolenic acid and linoleic acid also contribute to the nutritional quality of milk fat via their direct transfer from the feed into the milk. However, these FAs are extensively hydrogenated in the rumen, resulting in the production of saturated fatty acids (SFAs) that are subsequently transferred into the milk [33]. As the human consumption of foods high in SFAs has been linked to cardiovascular diseases, obesity and diabetes, there is a scientific interest to find strategies to decrease the rate of biohydrogenation. The legume and herbs in multispecies pasture systems contain phenolic compounds that could potentially decrease the biohydrogenation rate in the rumen [34]. Upon contact with oxygen, plant enzymes (e.g., polyphenol oxidase (PPO)) reduce phenols into quinones, which can link to the lipase and slow down its activity [35]. Less lipolysed fats can reduce the amount of free FAs available in the rumen, decrease biohydrogenation rates, and lower the formation rate of SFAs. Quinones could also affect the biological activity of the Butyrivibrio genus, which is the bacterium that is mainly responsible for the hydrogenation of α-linolenic and linoleic acid [36]. In vitro studies have revealed that quinones can inhibit the growth of Butyrivibrio, resulting in the accumulation of health-promoting biohydrogenation-intermediate compounds that flow out of the rumen and transfer into milk [37]. More studies on the impact of the multispecies sward FAs on the composition of milk are certainly necessary in order to understand the association between the multispecies pastures and improved milk quality.
Changes in the concentrations of carotenoids depends on several factors, such as plant species, weather conditions, nitrogen availability and conservation method [38].
Pasture-fed cattle can maintain considerable levels of carotenoids circulating in their blood, with variations being dependent on dietary patterns. During lactation, carotenoids are absorbed from the blood, transported to the mammary glands, and directly transferred into milk [25]. The potential benefits of consuming dairy products rich in carotenoids include enhanced immune function, cancer prevention, and protection against oxidative cellular damage [39,40]. High concentrations of carotenoids in milk also contribute to the quality and sensory characteristics of dairy products as they prevent the development of rancid flavours and provide the natural yellowish colour that consumers associate with acceptability.
Animal diet supplementation with carotenoids is not a common practice; therefore, ruminants need to assimilate carotenoids from their diet, with recommendations of between 300 and 500 mg of β-carotene daily [41] and around 400 mg of lutein daily [14]. As dairy cattle consume between 16 and 18 kg of DM per day [42], multispecies are expected to provide sufficient levels of β-carotene and lutein.
As observed from previously reported studies, α-tocopherol typically dominates the total tocopherol pool (α-, γ-, and δ-) of pastures [25]. However, considerably lower concentrations of α-tocopherol were found here in comparison with other studies [38,43]. Lynch [44] also identified low levels of α-tocopherol when studying grass and legume species in Ireland. Vascular plants have been reported to have high variations in tocopherol contents [25]. Highland plants usually contain elevated concentrations due to the persistence of severe stress factors such as high UV radiance and low temperatures [45]. Indeed, tocopherols are produced as a defence mechanism to eliminate the harmful reactive oxygens originating from oxidative stress [46]. In this study, pastures were grown in lowlands, which might have affected the levels of tocopherols. There were sudden increases in the concentration of α-tocopherol in PRG and PRG+WC in the middle of the season (July), which could have been due to increases in UV light. In fact, higher temperatures and sunlight were recorded in Ireland in the month of July–August 2020 [47].
All the tocopherols and tocotrienol chemical forms (α-, β-, γ-, and δ-) can be included in the generic term vitamin E. Lactating cows are recommended to be fed with 335 mg of vitamin E/day [48]. Vitamin E plays an important role in the immune defence system of animals [49], improves reproductive performance, and reduces the incidence of mastitis [25]. Furthermore, it has powerful antioxidant properties that can prevent FA oxidation in milk and off-flavour development.
Although this study found that the concentrations of vitamin E in the three grazing systems were lower than the recommended amounts for lactating cows, multispecies could still have the potential to impact animal health and product quality. Indeed, multispecies contain various oxygen-species-scavenging compounds, such as polyphenols, iridoids (e.g., aucubin, catalpol, and acteoside), sesquiterpene lactones, and anthocyanins, which have also been shown to have anti-inflammatory and antioxidant properties [34,50]. In fact, a few studies have also proven the effectiveness of including herbs (chicory and plantain) in pasture systems to reduce mastitis in cows and increase the oxidative stability of FAs in milk [51]. Certainly, further research is needed to establish the relation between the multispecies swards, animal health and milk quality.
As consumers become more aware of the nutritional and sustainable value of foods, ruminant diets are often being used as a parameter to assess the quality of the produced dairy products. In fact, pasture-raised animal products are perceived as more nutritious, natural and eco-friendly [52]. The present study investigated the differences in the FA, carotenoid and fat-soluble vitamin composition between a novel (MULTI) and two conventional (PRG and PRG+WC) grazing systems. There were no significant differences found in the concentrations of α-linolenic acid, linoleic acid, lutein and tocopherol among the studied grazing systems; thus, it could be suggested that MULTI have the potential to provide micronutrient levels similar to those of traditional pastures. Although PRG was identified to have the highest β-carotene level, MULTI was found to have enough levels of carotenoids to meet the nutrient requirement for ruminants. It is worth considering that MULTI has beneficial properties beyond fat composition. For instance, the herbs in MULTI contain phytochemicals that have the potential to positively impact the health of animals. In particular, various polyphenols have been associated with antioxidant, anti-inflammatory and anti-parasitic benefits [37]. Herbs contain high mineral contents, which are indispensable for optimal animal performance and productivity. Red clover is rich in polyphenol oxidase (PPO), an enzyme that can reduce proteolysis in the rumen, improve protein utilisation and lower nitrogen emissions. Finally, perennial ryegrass, plantain and timothy grow well during springtime, while white clover, red clover plantain and chicory grow well during summertime [53]. Hence, farmers establishing multispecies can benefit from available diversified pastures all year-round.
5. Conclusions
A multispecies pasture could be included in dairy production systems as an alternative to traditional pastures and provide an equivalent source of α-linolenic, linoleic acid, lutein and α-tocopherol concentrations in the ruminant diet. Although the monoculture ryegrass contained higher levels of β-carotene than the multispecies system in this study, the amounts were still above the recommended threshold. Farmers should consider implementing multispecies pastures because they contribute to the sustainability of dairy productions systems and contain non-fat nutrients that are linked to animal health and productivity.
Author Contributions
Conceptualisation, S.R. and N.A.-G.; methodology, S.R., G.O. and N.A.-G.; software, S.R.; validation, S.R., G.O. and N.A.-G.; formal analysis, S.R.; investigation, S.R.; data curation, S.R.; writing—original draft preparation, S.R.; writing—review and editing, G.O. and N.A.-G.; supervision, N.A.-G.; project administration, N.A.-G.; funding acquisition, N.A.-G. All authors have read and agreed to the published version of the manuscript.
Funding
The research reported herein was funded by the Irish Department of Agriculture, Food and the Marine’s competitive funding programmes (SMARTSWARD: “Future proofing Irish livestock sustainability” (17/S/267; Dublin Ireland)) in conjunction with the commercial industry.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data used to support the findings of this study are included within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bionaz, M.; Vargas-Bello-Pérez, E.; Busato, S. Advances in fatty acids nutrition in dairy cows: From gut to cells and effects on performance. J. Anim. Sci. Biotechnol. 2020, 11, 110. [Google Scholar] [CrossRef]
- Keenan, T.W.; Patton, S. The structure of milk. In Handbook of Milk Composition; Academic Press: Cambridge, MA, USA, 1995; pp. 5–50. [Google Scholar]
- Loften, J.R.; Linn, J.G.; Drackley, J.K.; Jenkins, T.C.; Soderholm, C.G.; Kertz, A.F. Invited review: Palmitic and stearic acid metabolism in lactating dairy cows. J. Dairy Sci. 2014, 97, 4661–4674. [Google Scholar] [CrossRef]
- Elgersma, A. Grazing increases the unsaturated fatty acid concentration of milk from grass-fed cows: A review of the contributing factors, challenges and future perspectives. Eur. J. Lipid Sci. Technol. 2015, 117, 1345–1369. [Google Scholar] [CrossRef]
- Teagasc. Grass Growth Curve. 2020. Available online: https://www.teagasc.ie/crops/grassland/pasturebase-ireland/grass-curve/ (accessed on 2 February 2023).
- Danielsson, H.; Nadeau, E.; Gustavsson, A.M.; Jensen, S.K.; Soegaard, K.; Nilsdotter-Linde, N. Contents of alpha-tocopherol and beta-carotene in grasses and legumes harvested at different maturities. Grassl. Sci. 2008, 13, 432–434. [Google Scholar]
- Piantoni, P.; Lock, A.L.; Allen, M.S. Palmitic acid increased yields of milk and milk fat and nutrient digestibility across production level of lactating cows. J. Dairy Sci. 2013, 96, 7143–7154. [Google Scholar] [CrossRef]
- Moallem, U. Invited review: Roles of dietary n-3 fatty acids in performance, milk fat composition, and reproductive and immune systems in dairy cattle. J. Dairy Sci. 2018, 101, 8641–8661. [Google Scholar] [CrossRef] [PubMed]
- Basak, S.; Duttaroy, A. Conjugated Linoleic Acid and Its Beneficial Effects in Obesity, Cardiovascular Disease, and Cancer. Nutrients 2020, 12, 1913. [Google Scholar] [CrossRef]
- Swanson, D.; Block, R.; Mousa, S.A. Omega-3 Fatty Acids EPA and DHA: Health Benefits Throughout Life. Adv. Nutr. Int. Rev. J. 2012, 3, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Shnimizu, M.; Moriwaki, H. Cancer Chemoprevention by Carotenoids. Molecules 2012, 17, 3202–3242. [Google Scholar] [CrossRef]
- Strickland, J.M.; Wisnieski, L.; Herdt, T.H.; Sordillo, L.M. Serum retinol, β-carotene, and α-tocopherol as biomarkers for disease risk and milk production in periparturient dairy cows. J. Dairy Sci. 2021, 104, 915–927. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Wang, H.; Yang, J.; Wang, J.; Duan, Z.; Wang, C.; Liu, J.; Lao, Y. Effects of feeding lutein on production performance, antioxidative status, and milk quality of high-yielding dairy cows. J. Dairy Sci. 2014, 97, 7144–7150. [Google Scholar] [CrossRef] [PubMed]
- Moyo, N.; Nielen, M.; Kruitwagen, C. Vitamin E supplementation and udder health: A metaanalysis. Mastitis in dairy production: Current knowledge and future solutions. In Proceedings of the 4th IDF International Mastitis Conference, Maastricht, The Netherlands, 12–15 June 2005; pp. 159–165. [Google Scholar]
- Palladino, R.A.; O’Donovan, M.; Kennedy, E.; Murphy, J.J.; Boland, T.M.; Kenny, D.A. Fatty acid composition and nutritive value of twelve cultivars of perennial ryegrass. Grass Forage Sci. 2009, 64, 219–226. [Google Scholar] [CrossRef]
- Jäpelt, R.B.; Didion, T.; Smedsgaard, J.; Jakobsen, J. Seasonal Variation of Provitamin D2 and Vitamin D2 in Perennial Ryegrass (Lolium perenne L.). J. Agric. Food Chem. 2011, 59, 10907–10912. [Google Scholar] [CrossRef]
- Gould, I.J.; Quinton, J.N.; Weigelt, A.; De Deyn, G.B.; Bardgett, R.D. Plant diversity and root traits benefit physical properties key to soil function in grasslands. Ecol. Lett. 2016, 19, 1140–1149. [Google Scholar] [CrossRef] [PubMed]
- Burke, J.L.; Waghorn, G.C.; Brookes, I.M.; Kolver, E.S.; Attwood, G.T. New Zealand Society of Animal Production online archive. N. Z. Soc. Anim. Prod. 2000, 60, 9–14. [Google Scholar]
- Cranston, L.M.; Kenyon, P.R.; Morris, S.T.; Kemp, P.D. A review of the use of chicory, plantain, red clover and white clover in a sward mix for increased sheep and beef production. J. N. Z. Grassl. 2015, 77, 89–94. [Google Scholar] [CrossRef]
- Brunton, N.P.; Mason, C.; Collins, M.J. Rapid microwave assisted preparation of fatty acid methyl esters for the analysis of fatty acid profiles in foods. J. Anal. Chem. 2015, 70, 1218–1224. [Google Scholar] [CrossRef]
- Gentili, A.; Caretti, F.; Bellante, S.; Ventura, S.; Canepari, S.; Curini, R. Comprehensive Profiling of Carotenoids and Fat-Soluble Vitamins in Milk from Different Animal Species by LC-DAD-MS/MS Hyphenation. J. Agric. Food Chem. 2013, 61, 1628–1639. [Google Scholar] [CrossRef]
- Kusch, P. (Ed.) Gas Chromatography—Derivatization, Sample Preparation, Application Derivatization Methods in GC and GC/MS; BoD–Books on Demand: Nordestedt, Germany, 2019; pp. 118–119. [Google Scholar]
- Clapham, W.M.; Foster, J.G.; Neel, J.P.S.; Fedders, J.M. Fatty Acid Composition of Traditional and Novel Forages. J. Agric. Food Chem. 2005, 53, 10068–10073. [Google Scholar] [CrossRef]
- Nozière, P.; Graulet, B.; Lucas, A.; Martin, B.; Grolier, P.; Doreau, M. Carotenoids for ruminants: From forages to dairy products. Anim. Feed. Sci. Technol. 2006, 131, 418–450. [Google Scholar] [CrossRef]
- Kalač, P. Carotenoids, ergosterol and tocopherols in fresh and preserved herbage and their transfer to bovine milk fat and adipose tissues: A review. J. Agrobiol. 2013, 29, 1–13. [Google Scholar] [CrossRef]
- Dewhurst, R.J.; Moorby, J.M.; Scollan, N.D.; Tweed, J.K.S.; Humphrey, M.O. Effect of a stay-green trait on the concentrations and stability of fatty acids perennial ryegrass. Grass Forage Sci. 2002, 57, 360–366. [Google Scholar] [CrossRef]
- Mc Clements, D.J.; Decker, E.A. Lipids. In Fennema’s Food Chemistry; CRC Press: Boca Raton, FL, USA, 2008; pp. 155–216. [Google Scholar] [CrossRef]
- Van Ranst, G.; Lee, M.R.F.; Fievez, V. Red clover polyphenol oxidase and lipid metabolism. Animal 2011, 5, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. An increase in the Omega-6/Omega-3 fatty acid ratio increases the risk for obesity. Nutrients 2016, 8, 128. [Google Scholar] [CrossRef]
- Marei, W.F.A.; De Bie, J.; Mohey-Elsaeed, O.; Wydooghe, E.; Bols, P.E.J.; Leroy, J.L.M.R. Alpha-linolenic acid protects the developmental capacity of bovine cumulus-oocyte complexes matured under lipotoxic conditions in vitro. Biol. Reprod. 2017, 96, 1181–1196. [Google Scholar] [CrossRef]
- Van Hoeck, V.; Sturmey, R.G.; Bermejo-Alvarez, P.; Rizos, D.; Gutierrez-Adan, A.; Leese, H.J.; Bols, P.E.J.; Leroy, J.L.M.R.; Kim, S. Elevated Non-Esterified Fatty Acid Concentrations during Bovine Oocyte Maturation Compromise Early Embryo Physiology. PLoS ONE 2011, 6, e23183. [Google Scholar] [CrossRef]
- Senger, P. Pathways to Pregnancy and Parturition, 2nd ed.; Current Conceptions, Inc.: Redmond, OR, USA, 2003. [Google Scholar]
- Khiaosa-Ard, R.; Klevenhusen, F.; Soliva, C.R.; Kreuzer, M.; Leiber, F. Transfer of linoleic and linolenic acid from feed to milk in cows fed isoenergetic diets differing in proportion and origin of concentrates and roughages. J. Dairy Res. 2010, 77, 331–336. [Google Scholar] [CrossRef]
- Rapisarda, S.; Abu-Ghannam, N. Polyphenol Characterization and Antioxidant Capacity of Multi-Species Swards Grown in Ireland—Environmental Sustainability and Nutraceutical Potential. Sustainability 2023, 15, 634. [Google Scholar] [CrossRef]
- Lee, M.R.F. Forage polyphenol oxidase and ruminant livestock nutrition. Front. Plant Sci. 2014, 5, 694. [Google Scholar] [CrossRef]
- Cabiddu, A.; Salis, L.; Tweed, J.K.S.; Molle, G.; Decandia, M.; Lee, M.R.F. The influence of plant polyphenols on lipolysis and biohydrogenation in dried forages at different phenological stages: In vitro study. J. Sci. Food Agric. 2010, 90, 829–835. [Google Scholar] [CrossRef]
- Vasta, V.; Makkar, H.P.S.; Mele, M.; Priolo, A. Ruminal biohydrogenation as affected by tannins in vitro. Br. J. Nutr. 2009, 102, 82–92. [Google Scholar] [CrossRef]
- Lindqvist, H.; Nadeau, E.; Jensen, S.K. Alpha-tocopherol and β-carotene in legume-grass mixtures as influenced by wilting, ensiling and type of silage additive. Grass Forage Sci. 2012, 67, 119–128. [Google Scholar] [CrossRef]
- Chew, B.P. Role of carotenoids in the immune response. J. Dairy Sci. 1993, 76, 2804–2811. [Google Scholar] [CrossRef]
- Martin, K.R.; Failla, M.L.; Smith, J.C. β-Carotene and Lutein Protect HepG2 Human Liver Cells against Oxidant-Induced Damage. J. Nutr. 1996, 126, 2098–2106. [Google Scholar] [CrossRef] [PubMed]
- Calsamiglia, S.; Rodríguez, M. Optimum Vitamin Nutrition in Dairy cattle. In Optimum Vitamin Nutrition in the Production of Quality Animal Foods; Publishing Benchmark House: Sheffield, UK, 2012; pp. 335–373. [Google Scholar]
- Kavanagh, S. Dairy Manual. 2016; Section 6, 34. Available online: https://www.teagasc.ie/media/website/animals/dairy/FeedingDiaryCow.pdf (accessed on 2 February 2023).
- Beeckman, A.; Vicca, J.; Van Ranst, G.; Janssens, G.P.J.; Fievez, V. Monitoring of vitamin E status of dry, early and mid-late lactating organic dairy cows fed conserved roughages during the indoor period and factors influencing forage vitamin E levels. J. Anim. Physiol. Anim. Nutr. 2010, 94, 736–746. [Google Scholar] [CrossRef]
- Lynch, A.; Kerry, J.P.; Buckley, D.J.; Morrissey, P.A. Lopez-Bote, C. Use of high pressure liquid chromatography (HPLC) for the determination of cda-tocopherol levels in forage (silage/grass) samples collected from different regions in Ireland. Food Chem. 2001, 72, 521–524. [Google Scholar] [CrossRef]
- Sickel, H. High level of alpha-tocopherol in Norwegian alpine grazing plants. J. Agric. Food Chem. 2012, 60, 7573–7580. [Google Scholar] [CrossRef] [PubMed]
- Munné-Bosch, S. The role of alpha-tocopherol in plant stress tolerance. J. Plant Physiol. 2005, 162, 743–748. [Google Scholar] [CrossRef]
- Met Eireann. Monthly Data, The Irish Meteorological Service. 2019. Available online: https://www.met.ie/climate/available-data/monthly-data (accessed on 1 February 2023).
- National Research Council (NRC). Nutrient Requirements of Dairy Cattle, 7th ed.; National Academy Press: Washington, DC, USA, 2001. [Google Scholar]
- Bernes, G.; Waller, K.P.; Jensen, S.K. Hay and silage as vitamin sources in organic sheep production. Grassl. Sci. Eur. 2008, 12, 251–255. [Google Scholar]
- Peña-Espinoza, M.; Thamsborg, S.M.; Desrues, O.; Hansen, T.V.A.; Enemark, H.L. Anthelmintic effects of forage chicory (Cichorium intybus) against gastrointestinal nematode parasites in experimentally infected cattle. Parasitology 2016, 143, 1279–1293. [Google Scholar] [CrossRef] [PubMed]
- Nkomboni, D.; Bryant, R.H.; Grant, E.R. Effect of increasing dietary proportion of plantain on milk production and nitrogen use of grazing dairy cows in late lactation. Anim. Prod. Sci. 2021, 61, 770–779. [Google Scholar] [CrossRef]
- Stampa, K.; Schipmann-Schwarze, C.; Hamm, U. Consumer perceptions, preferences, and behaviour regarding pasture-raised livestock products: A review. Food Qual. Prefer. 2020, 82, 103872. [Google Scholar] [CrossRef]
- Teagasc. Grassland Re-Seeding: How to Establish Multispecies Swards. 2020. Available online: https://www.teagasc.ie/publications/2020/grassland-re-seeding-how-to-establish-multispecies-swards.php (accessed on 2 February 2023).




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).