
Self-Assembled Monolayers Derived from Positively Charged Adsorbates on Plasmonic Substrates for MicroRNA Delivery: A Review

 , , , , , ,
, , , , , ,  ,
, 
Abstract
1. Introduction
2. Roles of miRNA in Gene Regulation
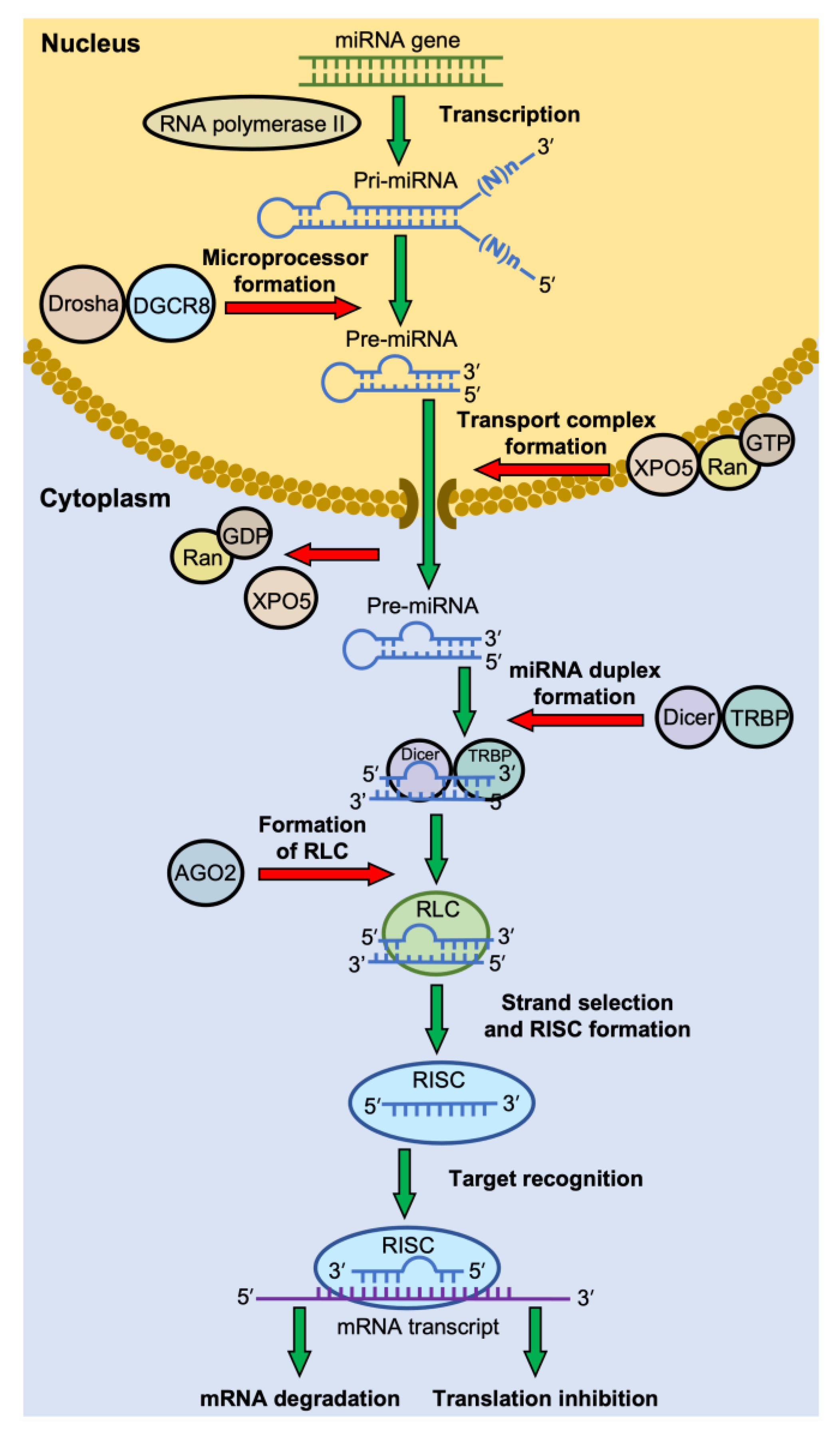
2.1. Biogenesis of miRNAs
2.2. RNA-Induced Silencing Complex (RISC) Formation
2.3. Target Recognition by RISC
2.4. Modes of Gene Silencing
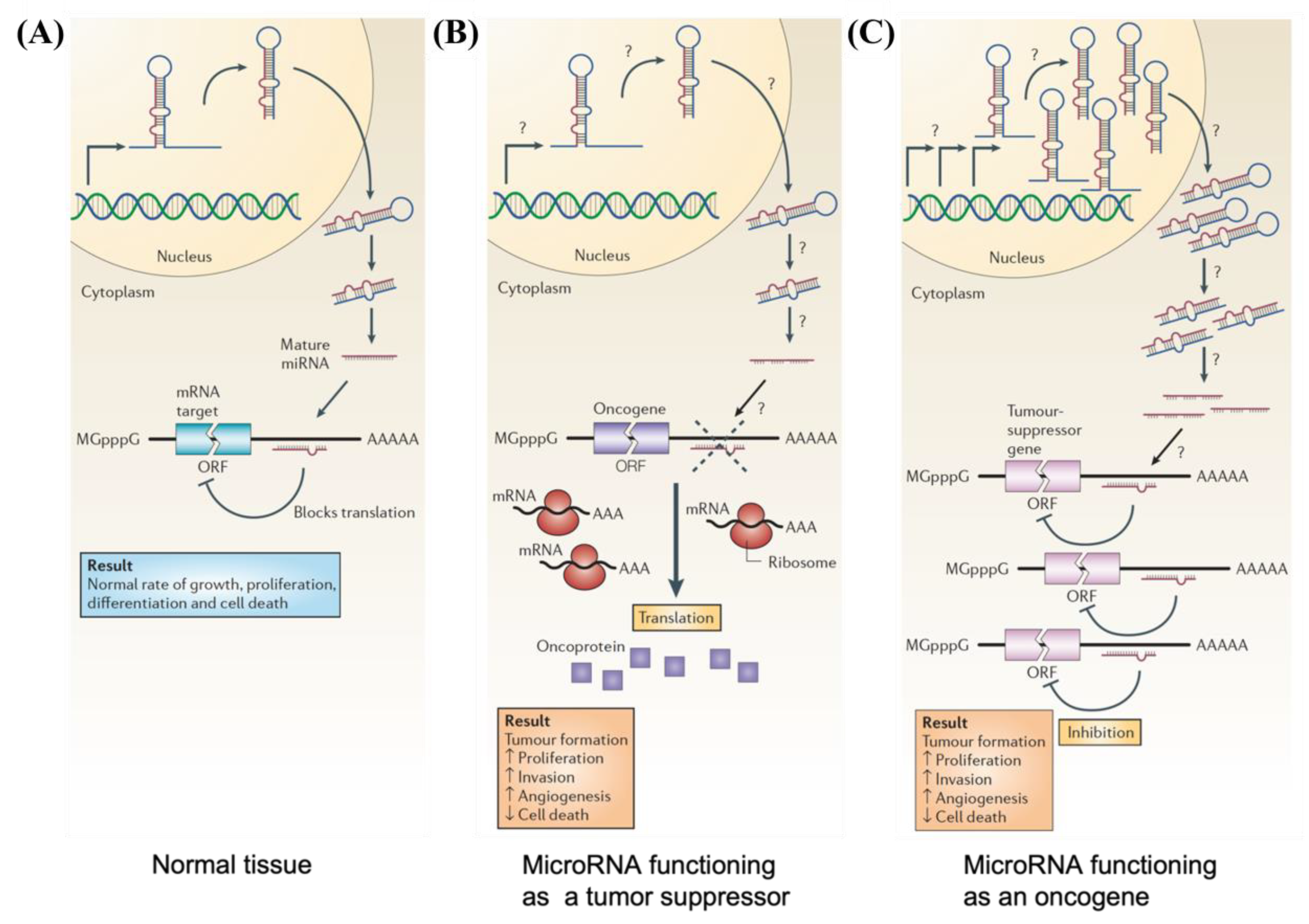
3. Role of MicroRNA in Cancer
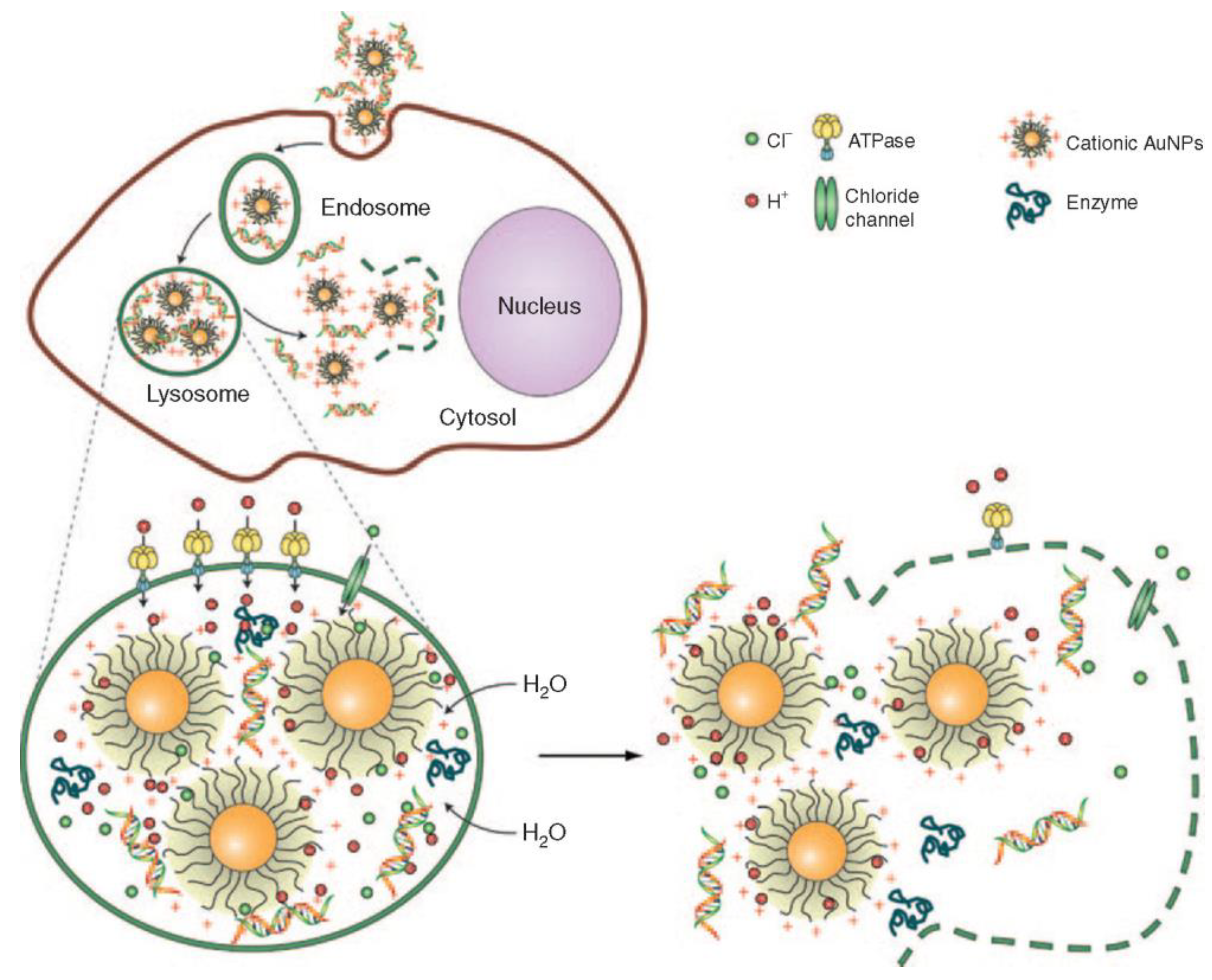
4. Endocytosis of Gold Nanoparticles
4.1. Gold Nanoparticles Entering the Lysosome
4.2. Gold Nanoparticles Exiting the Lysosome
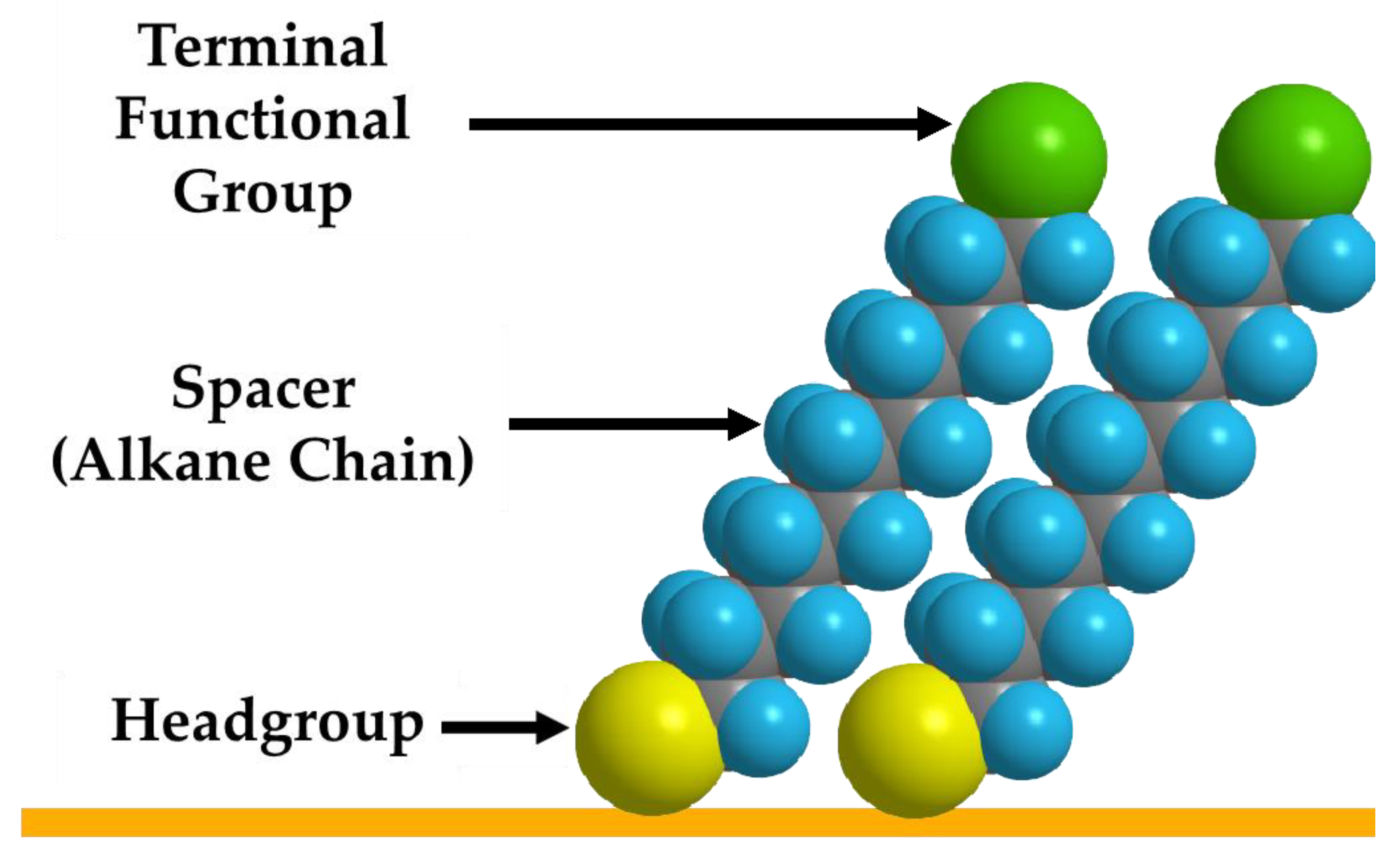
5. Using SAMs to Modify the Surfaces of Gold and Other Plasmonic Nanoparticles
6. Plasmonic Nanoparticles for miRNA Delivery/Detection
Nanoparticles for miRNA Delivery

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
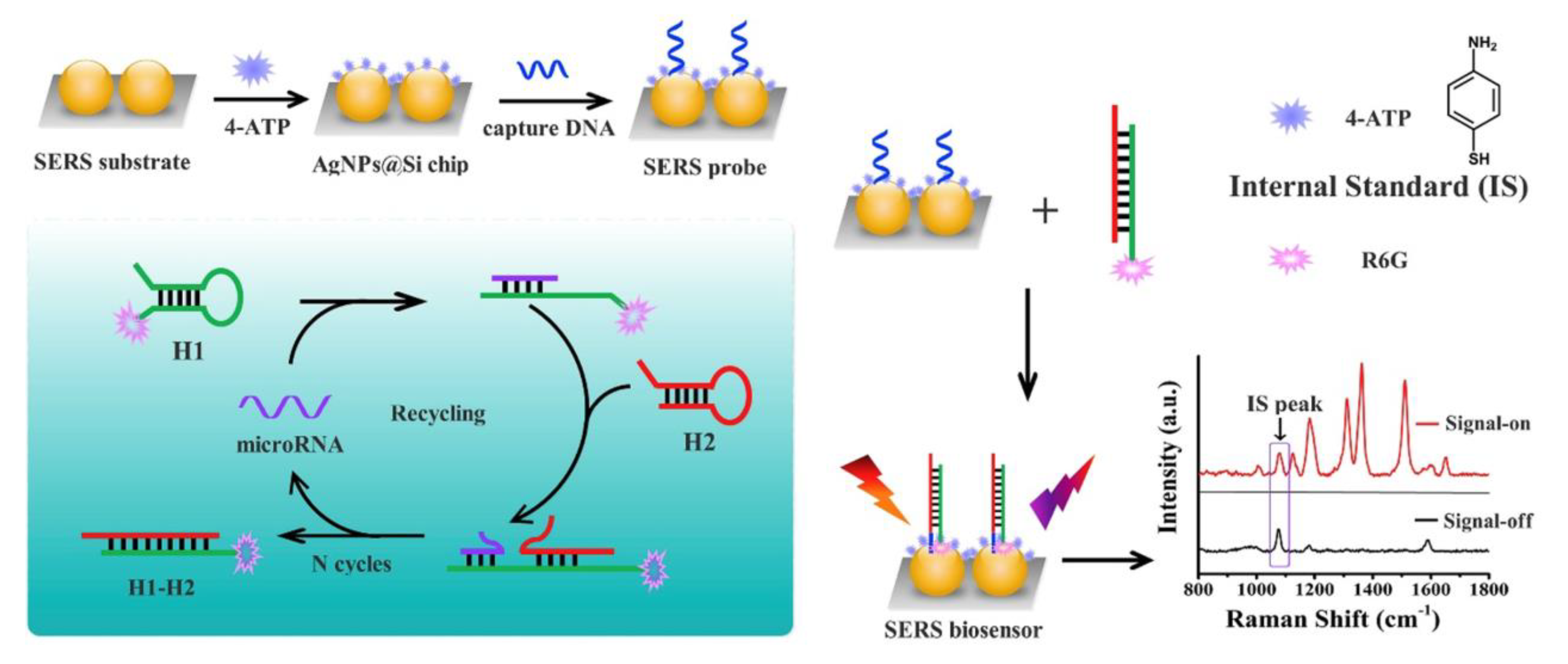{kind=link}
| Plasmonic Substrate | Morphology | Delivery Approach | Size (nm) | Conjugate | Target miRNA | Target Cell | Ref. |
|---|---|---|---|---|---|---|---|
| Au | Nanosphere (AuNPs) | miRNA inhibition | 2.6 | Terminated PAMAM | miR-21i | Cancer cells | [107] |
| miRNA inhibition | 5 | Carrier DNA | Anti-miRNA-712 | Endothelial cells | [108] | ||
| miRNA inhibition | 10 | Streptavidin | miRNA-491 | Breast cancer cells | [109] | ||
| miRNA inhibition | 13 | Thiol-modified miRNA | miRNA-182 | GBM cells | [110] | ||
| miRNA inhibition | 13 | Cargo DNA | MiR-29b | Myeloid cells | [111] | ||
| miRNA inhibition | 20 | Thiolated miRNA | MiR-214 | HEK293 cells | [112] | ||
| miRNA replacement | 20 | SH-PEG-NH2 | miRNA-206 | Breast cancer cells | [103] | ||
| miRNA replacement | 13 | Thiol-modified RNA | miRNA-205 and miRNA-20a | Prostate cancer cells | [106] | ||
| miRNA replacement | 13 | Thiolated miRNA | MiRNA-130b | Myeloma cells | [113] | ||
| miRNA replacement | 14 | Cysteamine | miRNA-31 and miRNA-1323 | Neuroblastoma cells and ovarian cancer cells | [114] | ||
| Hollow sphere | miRNA inhibition | 61 | Thiolated PAMAM | Anti-miR-21i | GBM cells | [115] | |
| miRNA inhibition | 150 | Thiolated miRNA | miRNA-34a | TNBC cells | [116] | ||
| Nanocage | miRNA inhibition | 30, 50, and 70 | SH-PEG-OMe | miRNA-26a | Cancer cells | [117] | |
| miRNA inhibition | 50 | Cationic polyethylene- imine | anti-miR-181b | HCC cells | [118] | ||
| miRNA replacement | 50 | Polyethylene-imine | miR-122 | HCC cells | [119] | ||
| Au-iron oxide | Nanostar | miRNA inhibition | 15 | β-cyclodextrin-chitosan hybrid polymers | AntimiR-21 and miR-100 | GBM cells | [120] |
| Au@Bi2Se3 | Sphere | miRNA inhibition | 11 | Thiol-modified ssDNA | antagomiRNA-152 | Neuroblastoma cells | [121] |
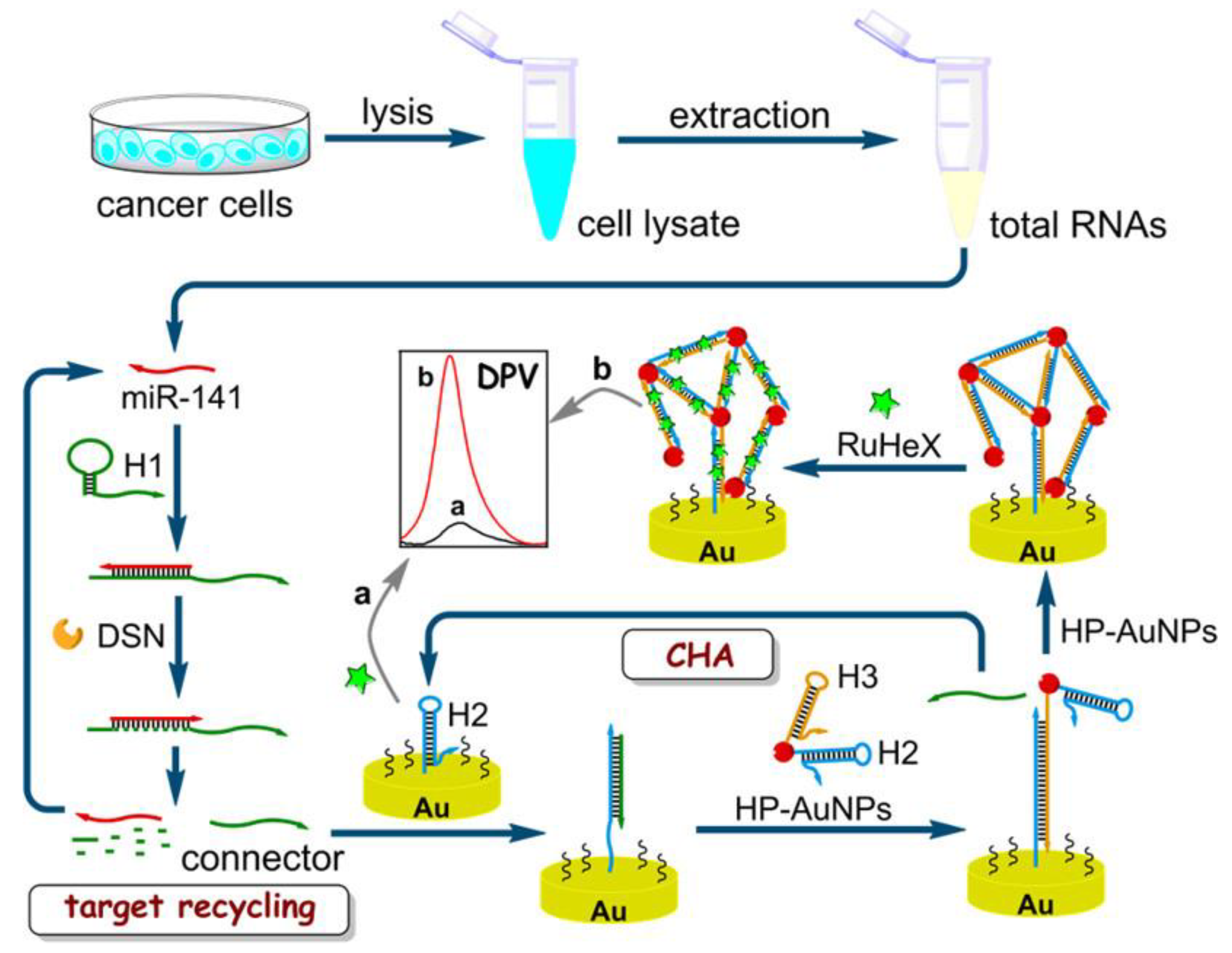
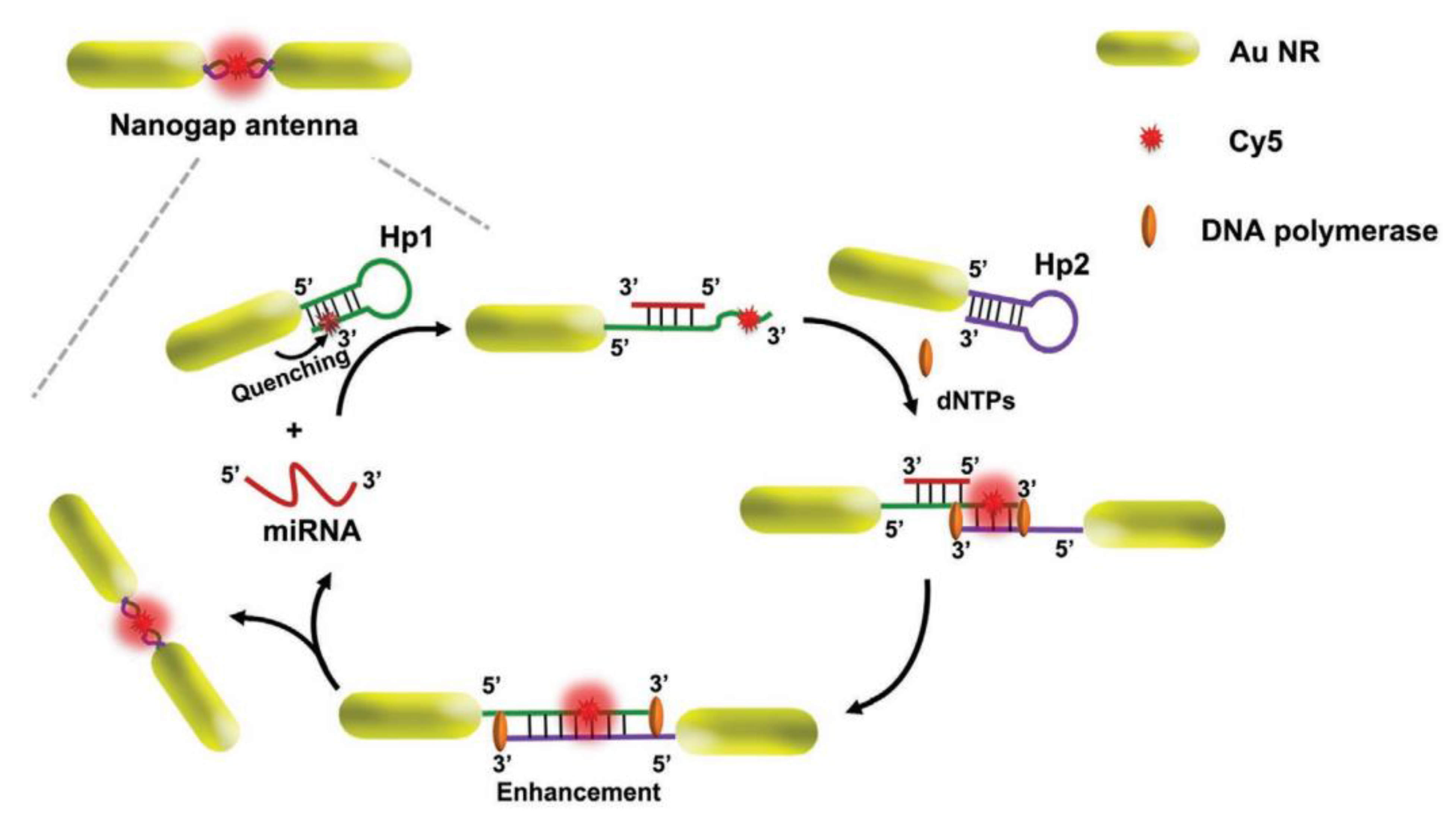
7. Nanoparticles for miRNA Diagnostics
| Plasmonic Substrate | Morphology | Type of Biosensor | Size (nm) | Conjugate | Target miRNA | Target Cell | Ref. |
|---|---|---|---|---|---|---|---|
| Au | Nanosphere (AuNPs) | Electrochemical | 13 | Thiol-labeled hairpin DNA | miR-141 | Breast cancer | [125] |
| Colorimetric | 19 | Thiolated probe | miRNA-155 | Cancer cells | [126] | ||
| Colorimetric | 22 | Thiolated RNA probes | miRNA-148a | Gastric cancer | [127] | ||
| Colorimetric | 20–30 | Thiol-modified DNA | miR-21 and miR-155 | Cancer cells | [128] | ||
| Colorimetric and FRET | 13 | Thiol-modified hairpin probe | miRNA-21 | Cancer cells | [124] | ||
| FRET | 13 | Thiol-modified oligonucleotides | miR-21 and miR-141 | Cancer cells | [129] | ||
| Thermophoretic | 13 | Thiol-modified single-stranded DNA | miR-375 | Breast cancer | [130] | ||
| Dark-field | 50 | Thiolated DNA | miRNA | - | [131] | ||
| Nanospike | PRAM | 100 | Thiolated DNA | miR-375 | Prostate cancer | [132] | |
| Nanorod (AuNRs) | PEF | 27 × 12 | Thiolated DNA | miRNA-21 | Cancer cells | [133] | |
| FRET | 60 | Thiolated DNA | miR-21 and miR-200b | Cancer cells | [134] | ||
| Nanocubes (AuNCs) | SPR | 50 | Thiol-modified single-strand DNA | miRNA-205 | Lung cancer | [135] | |
| Nanostripes | SPR | 200 × 100 | Thiolated DNA | miRNA-125b | - | [136] | |
| Mountaintop-shaped nanopillar | SERS | 800 × 200 | Thiol-modified DNA | miRNA-10b, miRNA-21, and miRNA-373 | Cancer Cells | [137] | |
| Ag | Nanosphere (AgNPs) | SERS | 110 | Thiolated DNA | miRNA-21 | Cancer cells | [87] |
| Nanoparticle films (AgNFs) | SERS | - | MBA-ssDNA, DSNB-ssDNA, and 6TG-ssDNA | miR-26a-5p, miR-223, and miR-27a-3p | Liver cancer | [138] | |
| Au-Ag | Nanosphere | SPR | 13 | Thiolated DNA | miRNA-21 | Cancer cells | [123] |
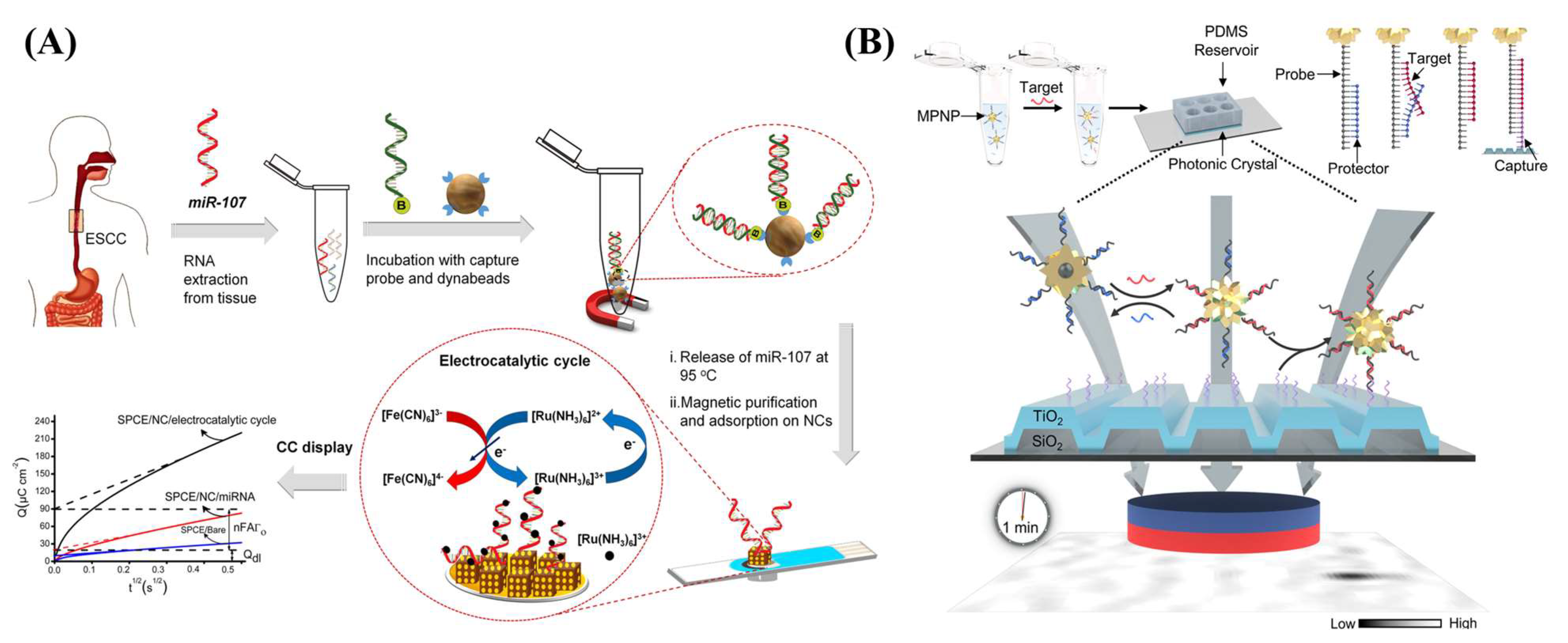
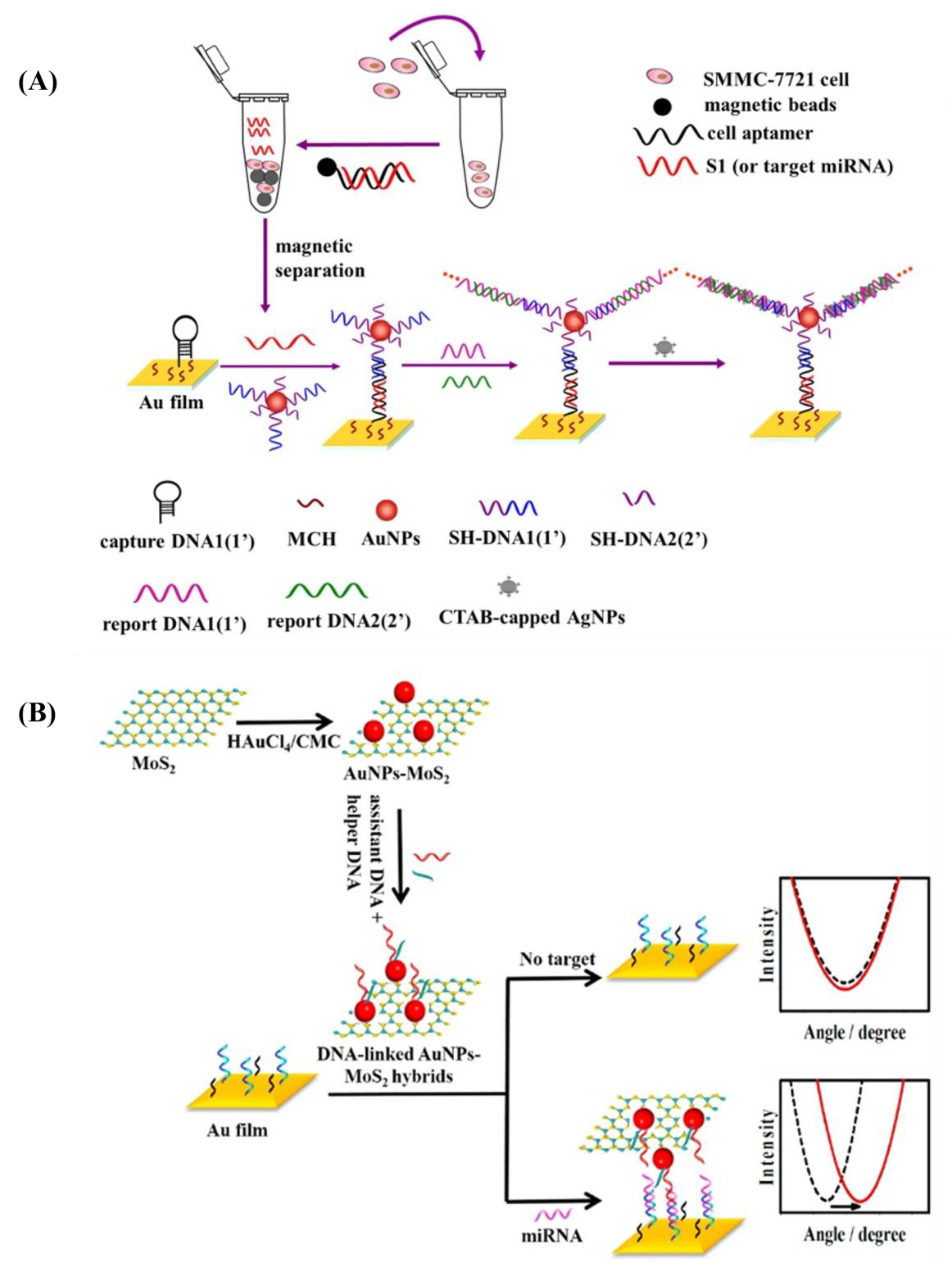
| Au@MoS2 | Nanosphere | SPR | 10 | Thiol-modified DNA | miRNA-141 | Cancer cells | [139] |
| Au@graph-ene oxide | Nanosphere (AuNPs-GO) | SPR | 18 | Thiolated capture DNA probe | miRNA-141 | Prostate cells | [140] |
| Fe3O4@Au | Nanospike | PRAM | 90 | SH-PEG-SA-B-DNA | miR-375 | Prostate cancer | [141] |
| Au@Pt | Nanosphere | FRET | 16 | Thiol-modified single-stranded DNA | miRNA-21 | Cancer cells | [89] |
| Au@Fe2O3 | Nanocube | Electrochemical | - | MB-SA-B-DNA | miR-107 | Cancer cells | [142] |
| Pd | Nanosphere (PdNPs) | Electrochemical | 30 | Thiolated miRNA-155 primer | miRNA-155 | Oncogenesis | [88] |
| Au-Ni-Au | Nanorod | MEF | 267 × 745 | Thiolated molecular beacon and anti-CD63 | miRNA-124 | Stem cell neurogenesis | [143] |
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guyon, N.; Garnier, D.; Briand, J.; Nadaradjane, A.; Bougras-Cartron, G.; Raimbourg, J.; Campone, M.; Heymann, D.; Vallette, F.M.; Frenel, J.-S.; et al. Anti-PD1 Therapy Induces Lymphocyte-Derived Exosomal MiRNA-4315 Release Inhibiting Bim-Mediated Apoptosis of Tumor Cells. Cell Death Dis. 2020, 11, 1048. [Google Scholar] [CrossRef]
- He, Z.; Li, W.; Zheng, T.; Liu, D.; Zhao, S. Human Umbilical Cord Mesenchymal Stem Cells-Derived Exosomes Deliver MicroRNA-375 to Downregulate ENAH and thus Retard Esophageal Squamous Cell Carcinoma Progression. J. Exp. Clin. Cancer Res. 2020, 39, 140. [Google Scholar] [CrossRef]
- Masoumi-Dehghi, S.; Babashah, S.; Sadeghizadeh, M. MicroRNA-141-3p-Containing Small Extracellular Vesicles Derived from Epithelial Ovarian Cancer Cells Promote Endothelial Cell Angiogenesis through Activating the JAK/STAT3 and NF-ΚB Signaling Pathways. J. Cell Commun. Signal. 2020, 14, 233–244. [Google Scholar] [CrossRef]
- Zhang, M.; Shi, Y.; Zhang, Y.; Wang, Y.; Alotaibi, F.; Qiu, L.; Wang, H.; Peng, S.; Liu, Y.; Li, Q.; et al. MiRNA-5119 Regulates Immune Checkpoints in Dendritic Cells to Enhance Breast Cancer Immunotherapy. Cancer Immunol. Immunother. 2020, 69, 951–967. [Google Scholar] [CrossRef]
- Hodge, J.; Wang, F.; Wang, J.; Liu, Q.; Saaoud, F.; Wang, Y.; Singh, U.P.; Chen, H.; Luo, M.; Ai, W.; et al. Overexpression of MicroRNA-155 Enhances the Efficacy of Dendritic Cell Vaccine against Breast Cancer. OncoImmunology 2020, 9, 1724761. [Google Scholar] [CrossRef]
- Zhao, H.; Li, T.; Yao, C.; Gu, Z.; Liu, C.; Li, J.; Yang, D. Dual Roles of Metal–Organic Frameworks as Nanocarriers for MiRNA Delivery and Adjuvants for Chemodynamic Therapy. ACS Appl. Mater. Interfaces 2021, 13, 6034–6042. [Google Scholar] [CrossRef]
- De Haan, P.; Van Diemen, F.R.; Toscano, M.G. Viral Gene Delivery Vectors: The next Generation Medicines for Immune-Related Diseases. Hum. Vaccines Immunother. 2021, 17, 14–21. [Google Scholar] [CrossRef]
- Wang, J.; Yu, L.; Zhou, A.; Liu, J.; Wang, K.; Luo, Y.; Wang, F. Non-Viral Gene Delivery Systems for Treatment of Myocardial Infarction: Targeting Strategies and Cardiac Cell Modulation. Pharmaceutics 2021, 13, 1520. [Google Scholar] [CrossRef]
- Tasset, A.; Bellamkonda, A.; Wang, W.; Pyatnitskiy, I.; Ward, D.; Peppas, N.; Wang, H. Overcoming Barriers in Non-Viral Gene Delivery for Neurological Applications. Nanoscale 2022, 14, 3698–3719. [Google Scholar] [CrossRef]
- Rodriguez, D.; Marquez, M.D.; Zenasni, O.; Han, L.T.; Baldelli, S.; Lee, T.R. Surface Dipoles Induce Uniform Orientation in Contacting Polar Liquids. Chem. Mater. 2020, 32, 7832–7841. [Google Scholar] [CrossRef]
- Bhardwaj, A.; Pandey, L.M. Design of Antibiofilm Surfaces by Immobilization of Biogenic Silver Nanoparticles on Amine Self-Assembled Monolayers. Mater. Lett. 2022, 311, 131574. [Google Scholar] [CrossRef]
- Durainatarajan, P.; Prabakaran, M.; Ramesh, S. Self-Assembled Monolayers of Novel Imidazole Derivative on Copper Surface for Anticorrosion Protection in Neutral Medium. J. Adhes. Sci. Technol. 2021, 35, 2580–2601. [Google Scholar] [CrossRef]
- Pathak, P.; Cho, H.J. Self-Assembled 1-Octadecanethiol Membrane on Pd/ZnO for a Selective Room Temperature Flexible Hydrogen Sensor. Micromachines 2022, 13, 26. [Google Scholar] [CrossRef] [PubMed]
- Singhana, B.; Jamison, A.C.; Hoang, J.; Lee, T.R. Self-Assembled Monolayer Films Derived from Tridentate Cyclohexyl Adsorbates with Alkyl Tailgroups of Increasing Chain Length. Langmuir 2013, 29, 14108–14116. [Google Scholar] [CrossRef] [PubMed]
- St. Hill, L.R.; Craft, J.W.; Chinwangso, P.; Tran, H.-V.; Marquez, M.D.; Lee, T.R. Antifouling Coatings Generated from Unsymmetrical Partially Fluorinated Spiroalkanedithiols. ACS Appl. Bio Mater. 2021, 4, 1563–1572. [Google Scholar] [CrossRef]
- St. Hill, L.R.; Tran, H.-V.; Chinwangso, P.; Lee, H.J.; Marquez, M.D.; Craft, J.W.; Lee, T.R. Antifouling Studies of Unsymmetrical Oligo(Ethylene Glycol) Spiroalkanedithiol Self-Assembled Monolayers. Micro 2021, 1, 151–163. [Google Scholar] [CrossRef]
- Tajalli, P.; Hernandez Rivera, J.M.; Omidiyan, M.; Tran, H.-V.; Lee, T.R. Carbonate-Terminated Self-Assembled Monolayers for Mimicking Nanoscale Polycarbonate Surfaces. ACS Appl. Nano Mater. 2023, 6, 2472–2477. [Google Scholar] [CrossRef]
- Wang, L.; Schubert, U.S.; Hoeppener, S. Surface Chemical Reactions on Self-Assembled Silane Based Monolayers. Chem. Soc. Rev. 2021, 50, 6507–6540. [Google Scholar] [CrossRef]
- Auzelle, T.; Ullrich, F.; Hietzschold, S.; Sinito, C.; Brackmann, S.; Kowalsky, W.; Mankel, E.; Brandt, O.; Lovrincic, R.; Fernández-Garrido, S. External Control of GaN Band Bending Using Phosphonate Self-Assembled Monolayers. ACS Appl. Mater. Interfaces 2021, 13, 4626–4635. [Google Scholar] [CrossRef]
- Jiang, C.; Huang, F.; Chen, Y.; Jiang, L. Highly Uniform Self-Assembled Monolayers of Silver Nanospheres for the Sensitive and Quantitative Detection of Glutathione by SERS. Dalton Trans. 2021, 50, 10436–10445. [Google Scholar] [CrossRef]
- Sakunkaewkasem, S.; Gonzalez, M.A.; Marquez, M.D.; Lee, T.R. Olefin-Bridged Bidentate Adsorbates for Generating Self-Assembled Monolayers on Gold. Langmuir 2020, 36, 10699–10707. [Google Scholar] [CrossRef]
- Hoang, J.; Park, C.S.; Marquez, M.D.; Gunaratne, P.H.; Lee, T.R. DNA Binding on Self-Assembled Monolayers Terminated with Mixtures of Ammonium and Trimethylammonium Groups: Toward a Gene-Delivery Platform. ACS Appl. Nano Mater. 2020, 3, 6621–6628. [Google Scholar] [CrossRef]
- Yu, T.; Marquez, M.D.; Tran, H.-V.; Lee, T.R. Crosslinked Organosulfur-Based Self-Assembled Monolayers: Formation and Applications. Soft Sci. 2022, 2, 5. [Google Scholar] [CrossRef]
- Sánchez-Paniagua, M.; Palenzuela-Batista, S.; Manzanares-Palenzuela, C.L.; López-Ruiz, B. Electrochemical Genosensor for Klotho Detection Based on Aliphatic and Aromatic Thiols Self-Assembled Monolayers. Talanta 2020, 212, 120735. [Google Scholar] [CrossRef] [PubMed]
- Hoang, J.; Park, C.S.; Lee, H.J.; Marquez, M.D.; Zenasni, O.; Gunaratne, P.H.; Lee, T.R. Quaternary Ammonium-Terminated Films Formed from Mixed Bidentate Adsorbates Provide a High-Capacity Platform for Oligonucleotide Delivery. ACS Appl. Mater. Interfaces 2018, 10, 40890–40900. [Google Scholar] [CrossRef] [PubMed]
- Diener, C.; Keller, A.; Meese, E. Emerging Concepts of MiRNA Therapeutics: From Cells to Clinic. Trends Genet. 2022, 38, 613–626. [Google Scholar] [CrossRef] [PubMed]
- Friedman, R.C.; Farh, K.K.-H.; Burge, C.B.; Bartel, D.P. Most Mammalian MRNAs Are Conserved Targets of MicroRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef]
- Liu, B.; Shyr, Y.; Cai, J.; Liu, Q. Interplay between MiRNAs and Host Genes and Their Role in Cancer. Brief. Funct. Genom. 2019, 18, 255–266. [Google Scholar] [CrossRef]
- Roberts, T.C. The MicroRNA Biology of the Mammalian Nucleus. Mol. Ther. Nucleic Acids 2014, 3, e188. [Google Scholar] [CrossRef]
- Kim, Y.-K.; Kim, V.N. Processing of Intronic MicroRNAs. EMBO J. 2007, 26, 775–783. [Google Scholar] [CrossRef]
- He, C.; Li, Z.; Chen, P.; Huang, H.; Hurst, L.D.; Chen, J. Young Intragenic MiRNAs Are Less Coexpressed with Host Genes than Old Ones: Implications of MiRNA–Host Gene Coevolution. Nucleic Acids Res. 2012, 40, 4002–4012. [Google Scholar] [CrossRef] [PubMed]
- Marsico, A.; Huska, M.R.; Lasserre, J.; Hu, H.; Vucicevic, D.; Musahl, A.; Orom, U.A.; Vingron, M. PROmiRNA: A New MiRNA Promoter Recognition Method Uncovers the Complex Regulation of Intronic MiRNAs. Genome Biol. 2013, 14, R84. [Google Scholar] [CrossRef] [PubMed]
- Borchert, G.M.; Lanier, W.; Davidson, B.L. RNA Polymerase III Transcribes Human MicroRNAs. Nat. Struct. Mol. Biol. 2006, 13, 1097–1101. [Google Scholar] [CrossRef]
- Diebel, K.W.; Smith, A.L.; van Dyk, L.F. Mature and Functional Viral MiRNAs Transcribed from Novel RNA Polymerase III Promoters. RNA 2010, 16, 170–185. [Google Scholar] [CrossRef] [PubMed]
- Saini, H.K.; Griffiths-Jones, S.; Enright, A.J. Genomic Analysis of Human MicroRNA Transcripts. Proc. Natl. Acad. Sci. USA 2007, 104, 17719–17724. [Google Scholar] [CrossRef] [PubMed]
- Altuvia, Y.; Landgraf, P.; Lithwick, G.; Elefant, N.; Pfeffer, S.; Aravin, A.; Brownstein, M.J.; Tuschl, T.; Margalit, H. Clustering and Conservation Patterns of Human MicroRNAs. Nucleic Acids Res. 2005, 33, 2697–2706. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef]
- Ha, M.; Kim, V.N. Regulation of MicroRNA Biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Roden, C.; Gaillard, J.; Kanoria, S.; Rennie, W.; Barish, S.; Cheng, J.; Pan, W.; Liu, J.; Cotsapas, C.; Ding, Y.; et al. Novel Determinants of Mammalian Primary MicroRNA Processing Revealed by Systematic Evaluation of Hairpin-Containing Transcripts and Human Genetic Variation. Genome Res. 2017, 27, 374–384. [Google Scholar] [CrossRef]
- Yoshida, T.; Asano, Y.; Ui-Tei, K. Modulation of MicroRNA Processing by Dicer via Its Associated DsRNA Binding Proteins. Non-Coding RNA 2021, 7, 57. [Google Scholar] [CrossRef]
- Sheng, P.; Fields, C.; Aadland, K.; Wei, T.; Kolaczkowski, O.; Gu, T.; Kolaczkowski, B.; Xie, M. Dicer Cleaves 5′-Extended MicroRNA Precursors Originating from RNA Polymerase II Transcription Start Sites. Nucleic Acids Res. 2018, 46, 5737–5752. [Google Scholar] [CrossRef] [PubMed]
- Iwakawa, H.; Tomari, Y. Life of RISC: Formation, Action, and Degradation of RNA-Induced Silencing Complex. Mol. Cell 2022, 82, 30–43. [Google Scholar] [CrossRef] [PubMed]
- Santhekadur, P.K.; Kumar, D.P. RISC Assembly and Post-Transcriptional Gene Regulation in Hepatocellular Carcinoma. Genes Dis. 2020, 7, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Dana, H.; Chalbatani, G.M.; Mahmoodzadeh, H.; Karimloo, R.; Rezaiean, O.; Moradzadeh, A.; Mehmandoost, N.; Moazzen, F.; Mazraeh, A.; Marmari, V.; et al. Molecular Mechanisms and Biological Functions of SiRNA. Int. J. Biomed. Sci. 2017, 13, 48–57. [Google Scholar]
- Lambeth, L.S.; Smith, C.A. Short Hairpin RNA-Mediated Gene Silencing. In Methods in Molecular Biology; Clifton, N.J., Ed.; Humana Press: Totowa, NJ, USA, 2013; pp. 205–232. ISBN 978-1-62703-118-9. [Google Scholar]
- Zhang, J.; Chen, S.; Liu, K. Structural Insights into PiRNA Biogenesis. Biochim. Biophys. Acta Gene Regul. Mech. 2022, 1865, 194799. [Google Scholar] [CrossRef]
- Wu, J.; Yang, J.; Cho, W.C.; Zheng, Y. Argonaute Proteins: Structural Features, Functions and Emerging Roles. J. Adv. Res. 2020, 24, 317–324. [Google Scholar] [CrossRef]
- Nakanishi, K. Anatomy of Four Human Argonaute Proteins. Nucleic Acids Res. 2022, 50, 6618–6638. [Google Scholar] [CrossRef]
- Rouya, C.; Siddiqui, N.; Morita, M.; Duchaine, T.F.; Fabian, M.R.; Sonenberg, N. Human DDX6 Effects MiRNA-Mediated Gene Silencing via Direct Binding to CNOT1. RNA 2014, 20, 1398–1409. [Google Scholar] [CrossRef]
- Hwang, H.-W.; Mendell, J.T. MicroRNAs in Cell Proliferation, Cell Death, and Tumorigenesis. Br. J. Cancer 2006, 94, 776–780. [Google Scholar] [CrossRef]
- Esquela-Kerscher, A.; Slack, F.J. Oncomirs—MicroRNAs with a Role in Cancer. Nat. Rev. Cancer 2006, 6, 259–269. [Google Scholar] [CrossRef]
- Fu, Z.; Wang, L.; Li, S.; Chen, F.; Au-Yeung, K.K.-W.; Shi, C. MicroRNA as an Important Target for Anticancer Drug Development. Front. Pharmacol. 2021, 12, 736323. [Google Scholar] [CrossRef]
- Forterre, A.; Komuro, H.; Aminova, S.; Harada, M. A Comprehensive Review of Cancer MicroRNA Therapeutic Delivery Strategies. Cancers 2020, 12, 1852. [Google Scholar] [CrossRef] [PubMed]
- Jansson, M.D.; Lund, A.H. MicroRNA and Cancer. Mol. Oncol. 2012, 6, 590–610. [Google Scholar] [CrossRef]
- Zhang, X.; Dong, H.; Tian, Y. MiRNA Biology in Pathological Processes. In MicroRNA Detection and Pathological Functions; Springer : Berlin/Heidelberg, Germany, 2015; pp. 7–22. ISBN 978-3-662-47293-4. [Google Scholar]
- Fuziwara, C.S.; Kimura, E.T. Insights into Regulation of the MiR-17-92 Cluster of MiRNAs in Cancer. Front. Med. 2015, 2, 64. [Google Scholar] [CrossRef] [PubMed]
- Saiyed, A.N.; Vasavada, A.R.; Johar, S.R.K. Recent Trends in MiRNA Therapeutics and the Application of Plant MiRNA for Prevention and Treatment of Human Diseases. Future J. Pharm. Sci. 2022, 8, 24. [Google Scholar] [CrossRef]
- He, B.; Zhao, Z.; Cai, Q.; Zhang, Y.; Zhang, P.; Shi, S.; Xie, H.; Peng, X.; Yin, W.; Tao, Y.; et al. MiRNA-Based Biomarkers, Therapies, and Resistance in Cancer. Int. J. Biol. Sci. 2020, 16, 2628–2647. [Google Scholar] [CrossRef]
- Tagliaferri, P.; Rossi, M.; Di Martino, M.T.; Amodio, N.; Leone, E.; Gulla, A.; Neri, A.; Tassone, P. Promises and Challenges of MicroRNA-Based Treatment of Multiple Myeloma. Curr. Cancer Drug Targets 2012, 12, 838–846. [Google Scholar] [CrossRef] [PubMed]
- De Jong, W.H.; Borm, P.J.A. Drug Delivery and Nanoparticles:Applications and Hazards. Int. J. Nanomed. 2008, 3, 133–149. [Google Scholar] [CrossRef] [PubMed]
- Yue, J.; Feliciano, T.J.; Li, W.; Lee, A.; Odom, T.W. Gold Nanoparticle Size and Shape Effects on Cellular Uptake and Intracellular Distribution of SiRNA Nanoconstructs. Bioconjug. Chem. 2017, 28, 1791–1800. [Google Scholar] [CrossRef]
- He, C.; Hu, Y.; Yin, L.; Tang, C.; Yin, C. Effects of Particle Size and Surface Charge on Cellular Uptake and Biodistribution of Polymeric Nanoparticles. Biomaterials 2010, 31, 3657–3666. [Google Scholar] [CrossRef]
- Shang, L.; Nienhaus, K.; Nienhaus, G.U. Engineered Nanoparticles Interacting with Cells: Size Matters. J. Nanobiotechnol. 2014, 12, 5. [Google Scholar] [CrossRef] [PubMed]
- Yameen, B.; Choi, W.I.; Vilos, C.; Swami, A.; Shi, J.; Farokhzad, O.C. Insight into Nanoparticle Cellular Uptake and Intracellular Targeting. J. Control. Release 2014, 190, 485–499. [Google Scholar] [CrossRef] [PubMed]
- Fröhlich, E. The Role of Surface Charge in Cellular Uptake and Cytotoxicity of Medical Nanoparticles. Int. J. Nanomed. 2012, 7, 5577–5591. [Google Scholar] [CrossRef] [PubMed]
- Noël, C.; Simard, J.-C.; Girard, D. Gold Nanoparticles Induce Apoptosis, Endoplasmic Reticulum Stress Events and Cleavage of Cytoskeletal Proteins in Human Neutrophils. Toxicol. Vitr. 2016, 31, 12–22. [Google Scholar] [CrossRef]
- Chithrani, B.D.; Ghazani, A.A.; Chan, W.C.W. Determining the Size and Shape Dependence of Gold Nanoparticle Uptake into Mammalian Cells. Nano Lett. 2006, 6, 662–668. [Google Scholar] [CrossRef]
- Nambara, K.; Niikura, K.; Mitomo, H.; Ninomiya, T.; Takeuchi, C.; Wei, J.; Matsuo, Y.; Ijiro, K. Reverse Size Dependences of the Cellular Uptake of Triangular and Spherical Gold Nanoparticles. Langmuir 2016, 32, 12559–12567. [Google Scholar] [CrossRef]
- Xie, X.; Liao, J.; Shao, X.; Li, Q.; Lin, Y. The Effect of Shape on Cellular Uptake of Gold Nanoparticles in the Forms of Stars, Rods, and Triangles. Sci. Rep. 2017, 7, 3827. [Google Scholar] [CrossRef]
- Kumal, R.R.; Landry, C.R.; Abu-Laban, M.; Hayes, D.J.; Haber, L.H. Monitoring the Photocleaving Dynamics of Colloidal MicroRNA-Functionalized Gold Nanoparticles Using Second Harmonic Generation. Langmuir 2015, 31, 9983–9990. [Google Scholar] [CrossRef]
- Verderio, P.; Avvakumova, S.; Alessio, G.; Bellini, M.; Colombo, M.; Galbiati, E.; Mazzucchelli, S.; Avila, J.P.; Santini, B.; Prosperi, D. Delivering Colloidal Nanoparticles to Mammalian Cells: A Nano–Bio Interface Perspective. Adv. Healthc. Mater. 2014, 3, 957–976. [Google Scholar] [CrossRef]
- Creusat, G.; Rinaldi, A.-S.; Weiss, E.; Elbaghdadi, R.; Remy, J.-S.; Mulherkar, R.; Zuber, G. Proton Sponge Trick for PH-Sensitive Disassembly of Polyethylenimine-Based SiRNA Delivery Systems. Bioconjug. Chem. 2010, 21, 994–1002. [Google Scholar] [CrossRef]
- Guo, S.; Huang, L. Nanoparticles Escaping RES and Endosome: Challenges for SiRNA Delivery for Cancer Therapy. J. Nanomater. 2011, 2011, 742895. [Google Scholar] [CrossRef]
- Ding, Y.; Jiang, Z.; Saha, K.; Kim, C.S.; Kim, S.T.; Landis, R.F.; Rotello, V.M. Gold Nanoparticles for Nucleic Acid Delivery. Mol. Ther. 2014, 22, 1075–1083. [Google Scholar] [CrossRef] [PubMed]
- Nel, A.E.; Mädler, L.; Velegol, D.; Xia, T.; Hoek, E.M.V.; Somasundaran, P.; Klaessig, F.; Castranova, V.; Thompson, M. Understanding Biophysicochemical Interactions at the Nano–Bio Interface. Nat. Mater. 2009, 8, 543–557. [Google Scholar] [CrossRef] [PubMed]
- Benjaminsen, R.V.; Mattebjerg, M.A.; Henriksen, J.R.; Moghimi, S.M.; Andresen, T.L. The Possible “Proton Sponge” Effect of Polyethylenimine (PEI) Does Not Include Change in Lysosomal PH. Mol. Ther. 2013, 21, 149–157. [Google Scholar] [CrossRef]
- Freeman, E.C.; Weiland, L.M.; Meng, W.S. Modeling the Proton Sponge Hypothesis: Examining Proton Sponge Effectiveness for Enhancing Intracellular Gene Delivery through Multiscale Modeling. J. Biomater. Sci. Polym. Ed. 2013, 24, 398–416. [Google Scholar] [CrossRef] [PubMed]
- Colangelo, E.; Comenge, J.; Paramelle, D.; Volk, M.; Chen, Q.; Lévy, R. Characterizing Self-Assembled Monolayers on Gold Nanoparticles. Bioconjug. Chem. 2017, 28, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Love, J.C.; Estroff, L.A.; Kriebel, J.K.; Nuzzo, R.G.; Whitesides, G.M. Self-Assembled Monolayers of Thiolates on Metals as a Form of Nanotechnology. Chem. Rev. 2005, 105, 1103–1170. [Google Scholar] [CrossRef] [PubMed]
- Vakarelski, I.U.; McNamee, C.E.; Higashitani, K. Deposition of Silica Nanoparticles on a Gold Surface via a Self-Assembled Monolayer of (3-Mercaptopropyl)Trimethoxysilane. Colloids Surf. A Physicochem. Eng. Asp. 2007, 295, 16–20. [Google Scholar] [CrossRef]
- Ahmed, S.R.; Oh, S.; Baba, R.; Zhou, H.; Hwang, S.; Lee, J.; Park, E.Y. Synthesis of Gold Nanoparticles with Buffer-Dependent Variations of Size and Morphology in Biological Buffers. Nanoscale Res. Lett. 2016, 11, 65. [Google Scholar] [CrossRef]
- Heddle, J.G. Gold Nanoparticle-Biological Molecule Interactions and Catalysis. Catalysts 2013, 3, 683–708. [Google Scholar] [CrossRef]
- Sperling, R.A.; García-Fernández, L.; Ojea-Jiménez, I.; Piella, J.; Bastús, N.G.; Puntes, V. One-Pot Synthesis of Cationic Gold Nanoparticles by Differential Reduction. Z. Phys. Chem. 2017, 231, 7–18. [Google Scholar] [CrossRef]
- Vericat, C.; Vela, M.E.; Corthey, G.; Pensa, E.; Cortés, E.; Fonticelli, M.H.; Ibañez, F.; Benitez, G.E.; Carro, P.; Salvarezza, R.C. Self-Assembled Monolayers of Thiolates on Metals: A Review Article on Sulfur-Metal Chemistry and Surface Structures. RSC Adv. 2014, 4, 27730–27754. [Google Scholar] [CrossRef]
- Smith, A.M.; Marbella, L.E.; Johnston, K.A.; Hartmann, M.J.; Crawford, S.E.; Kozycz, L.M.; Seferos, D.S.; Millstone, J.E. Quantitative Analysis of Thiolated Ligand Exchange on Gold Nanoparticles Monitored by 1H NMR Spectroscopy. Anal. Chem. 2015, 87, 2771–2778. [Google Scholar] [CrossRef]
- Smith, A.M.; Millstone, J.E. Ligand Exchange and 1H NMR Quantification of Single- and Mixed-Moiety Thiolated Ligand Shells on Gold Nanoparticles. In Biomedical Nanotechnology: Methods and Protocols; Petrosko, S.H., Day, E.S., Eds.; Springer: New York, NY, USA, 2017; pp. 17–29. ISBN 978-1-4939-6840-4. [Google Scholar]
- Chen, J.; Wu, Y.; Fu, C.; Cao, H.; Tan, X.; Shi, W.; Wu, Z. Ratiometric SERS Biosensor for Sensitive and Reproducible Detection of MicroRNA Based on Mismatched Catalytic Hairpin Assembly. Biosens. Bioelectron. 2019, 143, 111619. [Google Scholar] [CrossRef]
- Wu, X.; Chai, Y.; Yuan, R.; Su, H.; Han, J. A Novel Label-Free Electrochemical MicroRNA Biosensor Using Pd Nanoparticles as Enhancer and Linker. Analyst 2013, 138, 1060–1066. [Google Scholar] [CrossRef]
- Qing, Z.; Luo, G.; Xing, S.; Zou, Z.; Lei, Y.; Liu, J.; Yang, R. Pt–S Bond-Mediated Nanoflares for High-Fidelity Intracellular Applications by Avoiding Thiol Cleavage. Angew. Chem. Int. Ed. 2020, 59, 14044–14048. [Google Scholar] [CrossRef]
- Huang, D.; Zhou, H.; Liu, H.; Gao, J. The Cytotoxicity of Gold Nanoparticles Is Dispersity-Dependent. Dalton Trans. 2015, 44, 17911–17915. [Google Scholar] [CrossRef]
- Min, Y.; Akbulut, M.; Kristiansen, K.; Golan, Y.; Israelachvili, J. The Role of Interparticle and External Forces in Nanoparticle Assembly. Nat. Mater. 2008, 7, 527–538. [Google Scholar] [CrossRef]
- Rosická, D.; Šembera, J. Changes in the Nanoparticle Aggregation Rate Due to the Additional Effect of Electrostatic and Magnetic Forces on Mass Transport Coefficients. Nanoscale Res. Lett. 2013, 8, 20. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Huang, X.; Liu, H.; Zan, F.; Ren, J. Colloidal Stability of Gold Nanoparticles Modified with Thiol Compounds: Bioconjugation and Application in Cancer Cell Imaging. Langmuir 2012, 28, 4464–4471. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.; Shin, M.; Lee, J.-Y.; Lee, S.-N.; Choi, J.-H.; Choi, J.-W. RNA Interference (RNAi)-Based Plasmonic Nanomaterials for Cancer Diagnosis and Therapy. J. Control. Release 2022, 342, 228–240. [Google Scholar] [CrossRef] [PubMed]
- Moraes, F.C.; Pichon, C.; Letourneur, D.; Chaubet, F. miRNA Delivery by Nanosystems: State of the Art and Perspectives. Pharmaceutics 2021, 13, 1901. [Google Scholar] [CrossRef] [PubMed]
- Nahalka, J. The Role of the Protein–RNA Recognition Code in Neurodegeneration. Cell. Mol. Life Sci. 2019, 76, 2043–2058. [Google Scholar] [CrossRef] [PubMed]
- Cristina Caroleo, M.; De Sarro, G. Chapter 24—Overview of MicroRNA-Based Therapeutics. In MicroRNA; Xiao, J., Ed.; Academic Press: Cambridge, MA, USA, 2022; pp. 493–502. ISBN 978-0-323-89774-7. [Google Scholar]
- Muthiah, M.; Park, I.-K.; Cho, C.-S. Nanoparticle-Mediated Delivery of Therapeutic Genes: Focus on MiRNA Therapeutics. Expert Opin. Drug Deliv. 2013, 10, 1259–1273. [Google Scholar] [CrossRef] [PubMed]
- Arghiani, N.; Shah, K. Modulating MicroRNAs in Cancer: Next-Generation Therapies. Cancer Biol. Med. 2022, 19, 289. [Google Scholar] [CrossRef] [PubMed]
- Mansouri, S.; Lavigne, P.; Corsi, K.; Benderdour, M.; Beaumont, E.; Fernandes, J.C. Chitosan-DNA Nanoparticles as Non-Viral Vectors in Gene Therapy: Strategies to Improve Transfection Efficacy. Eur. J. Pharm. Biopharm. 2004, 57, 1–8. [Google Scholar] [CrossRef]
- Thomas, T.J.; Tajmir-Riahi, H.A.; Pillai, C.K.S. Biodegradable Polymers for Gene Delivery. Molecules 2019, 24, 3744. [Google Scholar] [CrossRef]
- Uchida, S.; Perche, F.; Pichon, C.; Cabral, H. Nanomedicine-Based Approaches for MRNA Delivery. Mol. Pharm. 2020, 17, 3654–3684. [Google Scholar] [CrossRef]
- Chaudhari, R.; Nasra, S.; Meghani, N.; Kumar, A. MiR-206 Conjugated Gold Nanoparticle Based Targeted Therapy in Breast Cancer Cells. Sci. Rep. 2022, 12, 4713. [Google Scholar] [CrossRef]
- Boca, S.; Gulei, D.; Zimta, A.-A.; Onaciu, A.; Magdo, L.; Tigu, A.B.; Ionescu, C.; Irimie, A.; Buiga, R.; Berindan-Neagoe, I. Nanoscale Delivery Systems for MicroRNAs in Cancer Therapy. Cell. Mol. Life Sci. 2020, 77, 1059–1086. [Google Scholar] [CrossRef]
- Lee, S.W.L.; Paoletti, C.; Campisi, M.; Osaki, T.; Adriani, G.; Kamm, R.D.; Mattu, C.; Chiono, V. MicroRNA Delivery through Nanoparticles. J. Control. Release 2019, 313, 80–95. [Google Scholar] [CrossRef] [PubMed]
- Hao, L.; Patel, P.C.; Alhasan, A.H.; Giljohann, D.A.; Mirkin, C.A. Nucleic Acid–Gold Nanoparticle Conjugates as Mimics of MicroRNA. Small 2011, 7, 3158–3162. [Google Scholar] [CrossRef] [PubMed]
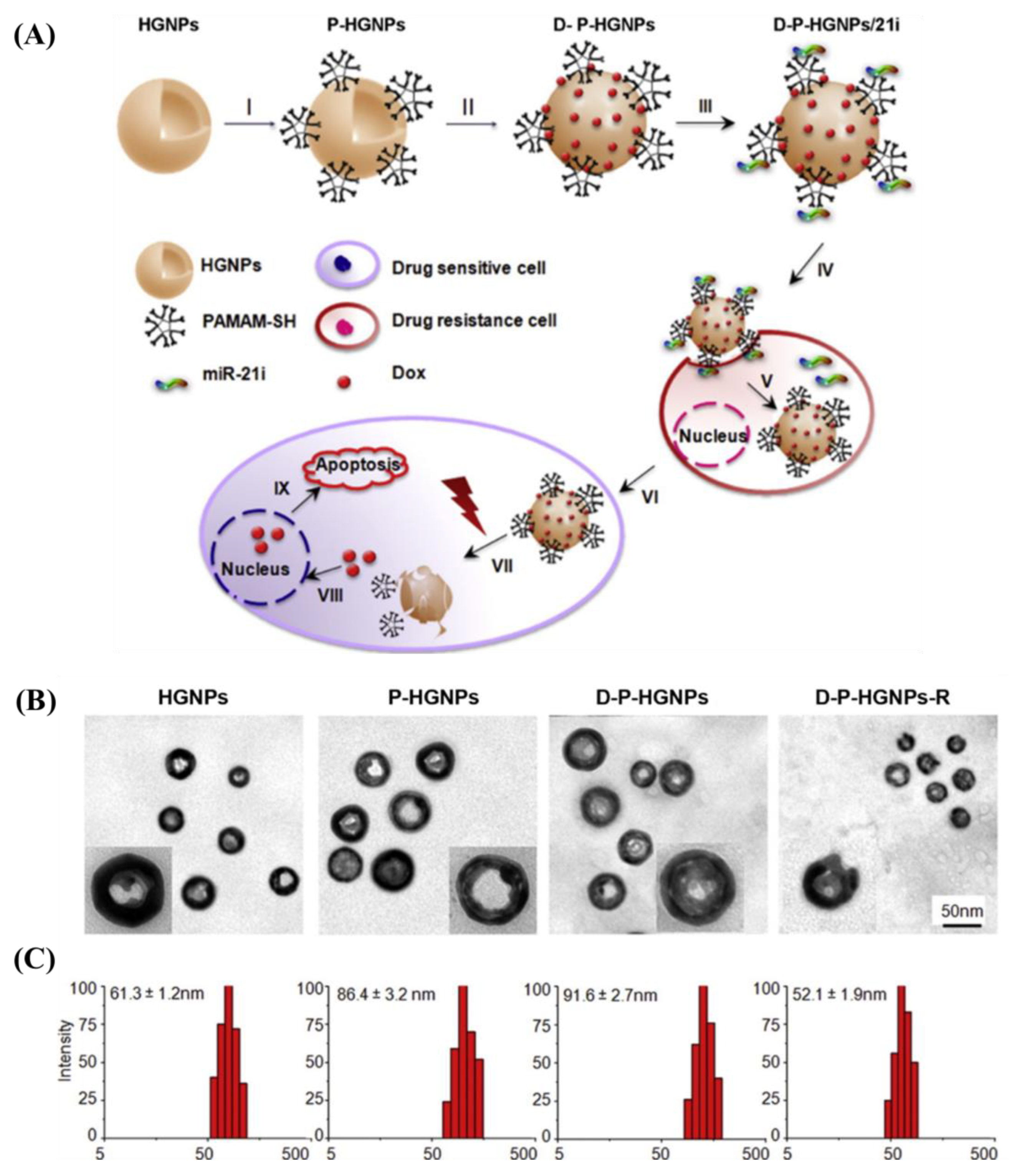
- Lin, L.; Fan, Y.; Gao, F.; Jin, L.; Li, D.; Sun, W.; Li, F.; Qin, P.; Shi, Q.; Shi, X.; et al. UTMD-Promoted Co-Delivery of Gemcitabine and MiR-21 Inhibitor by Dendrimer-Entrapped Gold Nanoparticles for Pancreatic Cancer Therapy. Theranostics 2018, 8, 1923–1939. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Simmons, R.; Huo, D.; Pang, B.; Zhao, X.; Kim, C.W.; Jo, H.; Xia, Y. Targeted Delivery of Anti-MiR-712 by VCAM1-Binding Au Nanospheres for Atherosclerosis Therapy. ChemNanoMat 2016, 2, 400–406. [Google Scholar] [CrossRef]
- Natarajan, A.; Venugopal, S.K.; DeNardo, S.J.; Zern, M.A. Breast Cancer Targeting Novel MicroRNA-Nanoparticles for Imaging. In Multimodal Biomedical Imaging IV; SPIE: Washington, DC, USA, 2009; Volume 7171. [Google Scholar]
- Kouri, F.M.; Hurley, L.A.; Daniel, W.L.; Day, E.S.; Hua, Y.; Hao, L.; Peng, C.-Y.; Merkel, T.J.; Queisser, M.A.; Ritner, C.; et al. MiR-182 Integrates Apoptosis, Growth, and Differentiation Programs in Glioblastoma. Genes Dev. 2015, 29, 732–745. [Google Scholar] [CrossRef]
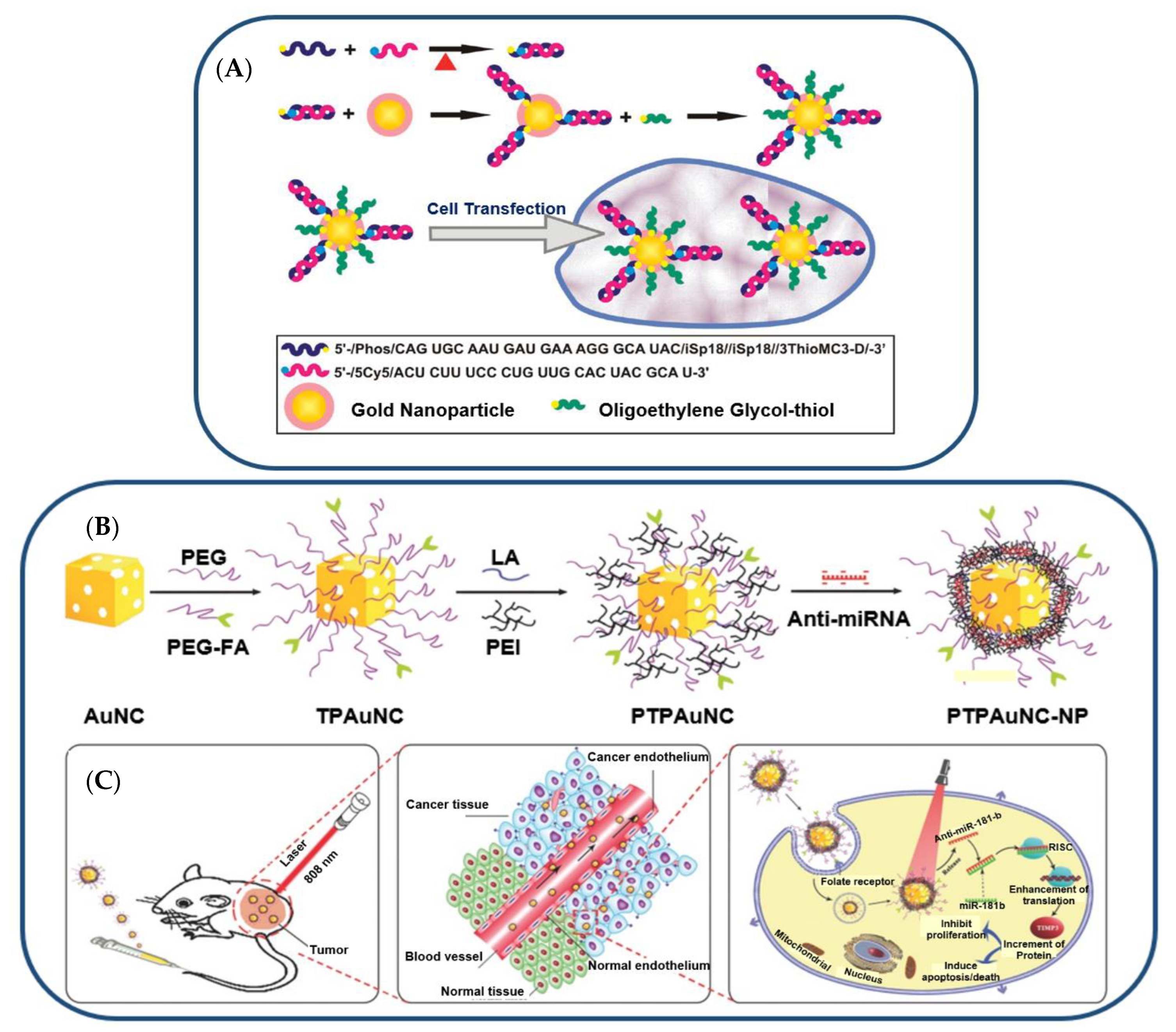
- Kim, J.-H.; Yeom, J.-H.; Ko, J.-J.; Han, M.S.; Lee, K.; Na, S.-Y.; Bae, J. Effective Delivery of Anti-MiRNA DNA Oligonucleotides by Functionalized Gold Nanoparticles. J. Biotechnol. 2011, 155, 287–292. [Google Scholar] [CrossRef]
- van der Ven, C.F.T.; Tibbitt, M.W.; Conde, J.; van Mil, A.; Hjortnaes, J.; Doevendans, P.A.; Sluijter, J.P.G.; Aikawa, E.; Langer, R.S. Controlled Delivery of Gold Nanoparticle-Coupled MiRNA Therapeutics via an Injectable Self-Healing Hydrogel. Nanoscale 2021, 13, 20451–20461. [Google Scholar] [CrossRef]
- Crew, E.; Rahman, S.; Razzak-Jaffar, A.; Mott, D.; Kamundi, M.; Yu, G.; Tchah, N.; Lee, J.; Bellavia, M.; Zhong, C.-J. MicroRNA Conjugated Gold Nanoparticles and Cell Transfection. Anal. Chem. 2012, 84, 26–29. [Google Scholar] [CrossRef]
- Ghosh, R.; Singh, L.C.; Shohet, J.M.; Gunaratne, P.H. A Gold Nanoparticle Platform for the Delivery of Functional MicroRNAs into Cancer Cells. Biomaterials 2013, 34, 807–816. [Google Scholar] [CrossRef]
- Ren, Y.; Wang, R.; Gao, L.; Li, K.; Zhou, X.; Guo, H.; Liu, C.; Han, D.; Tian, J.; Ye, Q.; et al. Sequential Co-Delivery of MiR-21 Inhibitor Followed by Burst Release Doxorubicin Using NIR-Responsive Hollow Gold Nanoparticle to Enhance Anticancer Efficacy. J. Control. Release 2016, 228, 74–86. [Google Scholar] [CrossRef]
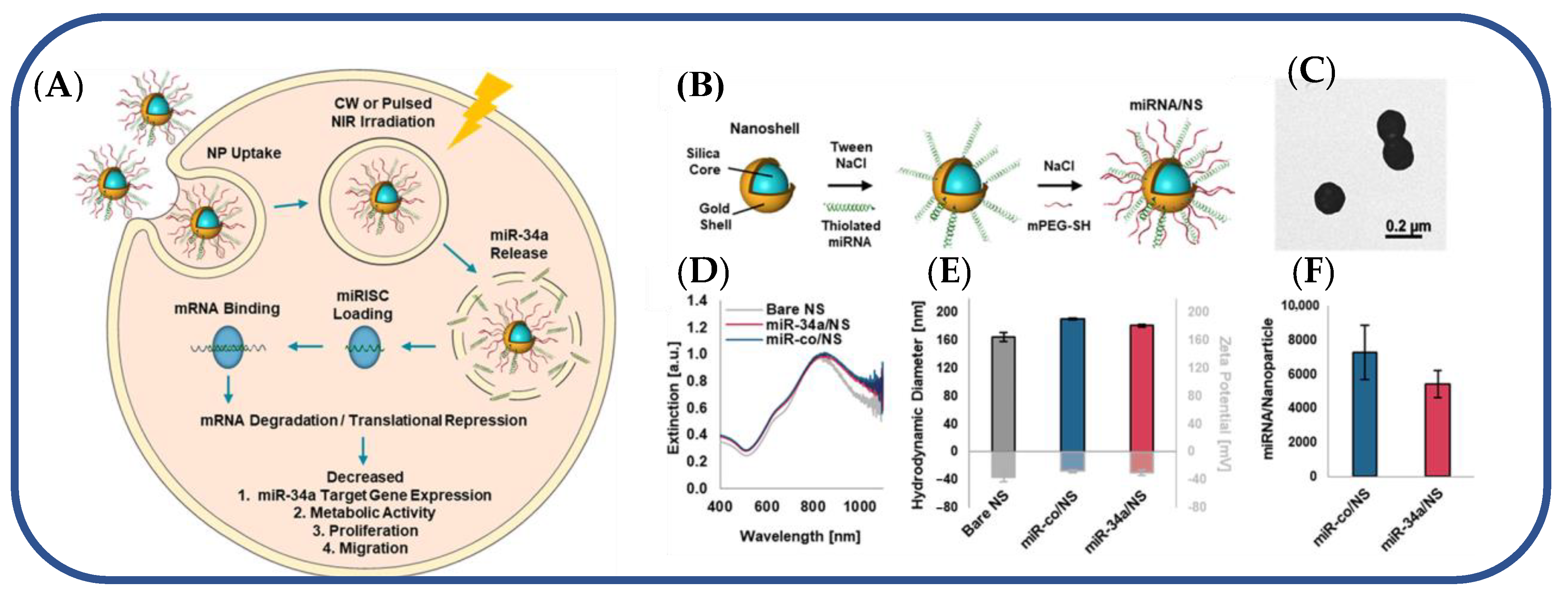
- Dang, M.N.; Gomez Casas, C.; Day, E.S. Photoresponsive MiR-34a/Nanoshell Conjugates Enable Light-Triggered Gene Regulation to Impair the Function of Triple-Negative Breast Cancer Cells. Nano Lett. 2021, 21, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Bao, S.; Huang, S.; Liu, Y.; Hu, Y.; Wang, W.; Ji, M.; Li, H.; Zhang, N.X.; Song, C.; Duan, S. Gold Nanocages with Dual Modality for Image-Guided Therapeutics. Nanoscale 2017, 9, 7284–7296. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Duan, S.; Wang, J.; Bao, S.; Qiu, X.; Li, C.; Liu, Y.; Yan, L.; Zhang, Z.; Hu, Y. Folic-Acid-Mediated Functionalized Gold Nanocages for Targeted Delivery of Anti-MiR-181b in Combination of Gene Therapy and Photothermal Therapy against Hepatocellular Carcinoma. Adv. Funct. Mater. 2016, 26, 2532–2544. [Google Scholar] [CrossRef]
- Huang, S.; Liu, Y.; Xu, X.; Ji, M.; Li, Y.; Song, C.; Duan, S.; Hu, Y. Triple Therapy of Hepatocellular Carcinoma with MicroRNA-122 and Doxorubicin Co-Loaded Functionalized Gold Nanocages. J. Mater. Chem. B 2018, 6, 2217–2229. [Google Scholar] [CrossRef]
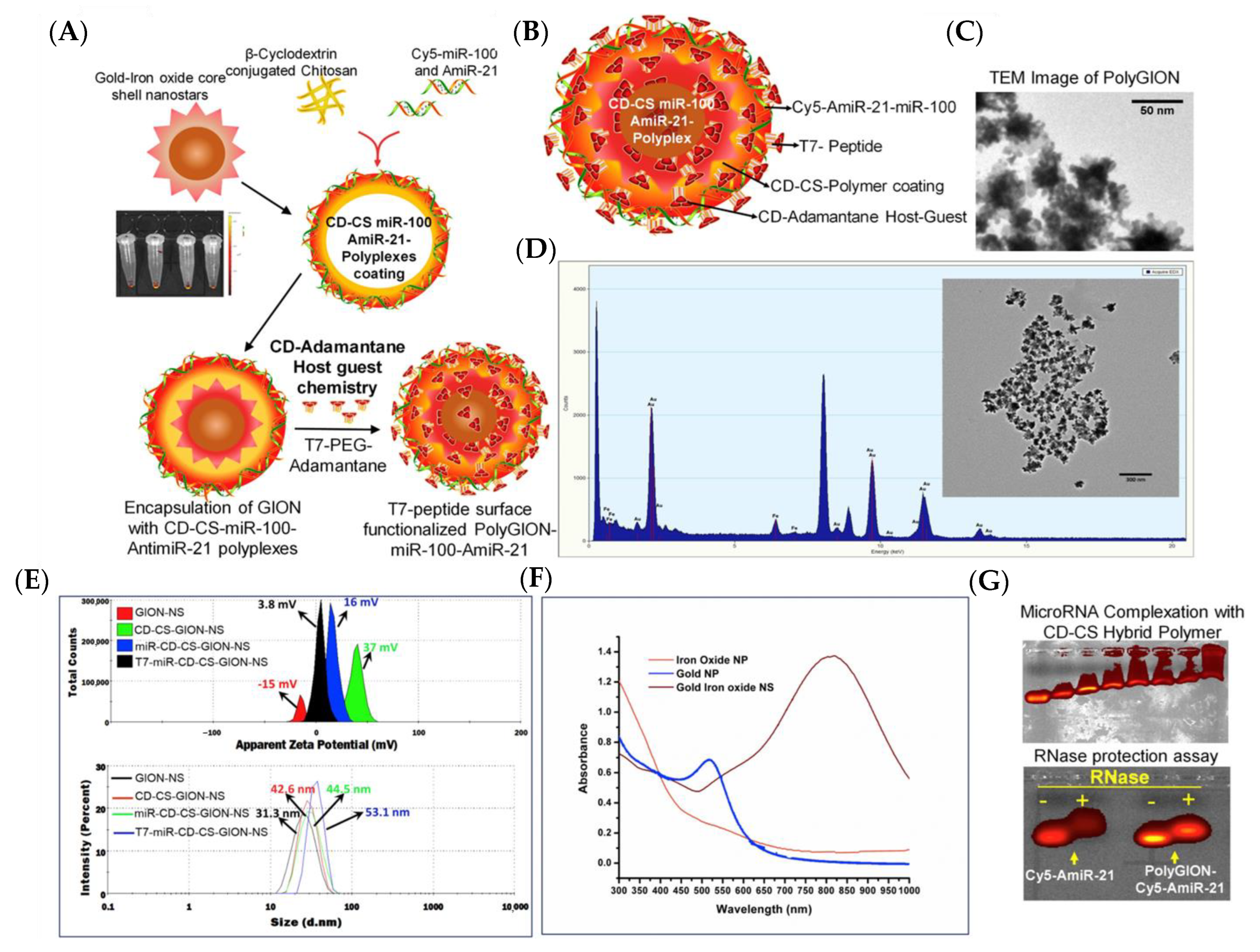
- Sukumar, U.K.; Bose, R.J.C.; Malhotra, M.; Babikir, H.A.; Afjei, R.; Robinson, E.; Zeng, Y.; Chang, E.; Habte, F.; Sinclair, R.; et al. Intranasal Delivery of Targeted Polyfunctional Gold–Iron Oxide Nanoparticles Loaded with Therapeutic MicroRNAs for Combined Theranostic Multimodality Imaging and Presensitization of Glioblastoma to Temozolomide. Biomaterials 2019, 218, 119342. [Google Scholar] [CrossRef]
- Mohammadniaei, M.; Lee, T.; Bharate, B.G.; Yoon, J.; Choi, H.K.; Park, S.; Kim, J.; Kim, J.; Choi, J.-W. Bifunctional Au@Bi2Se3 Core–Shell Nanoparticle for Synergetic Therapy by SERS-Traceable AntagomiR Delivery and Photothermal Treatment. Small 2018, 14, 1802934. [Google Scholar] [CrossRef]
- Duff, D.G.; Baiker, A.; Edwards, P.P. A New Hydrosol of Gold Clusters. 1. Formation and Particle Size Variation. Langmuir 1993, 9, 2301–2309. [Google Scholar] [CrossRef]
- Liu, R.; Wang, Q.; Li, Q.; Yang, X.; Wang, K.; Nie, W. Surface Plasmon Resonance Biosensor for Sensitive Detection of MicroRNA and Cancer Cell Using Multiple Signal Amplification Strategy. Biosens. Bioelectron. 2017, 87, 433–438. [Google Scholar] [CrossRef]
- Huang, J.; Shangguan, J.; Guo, Q.; Ma, W.; Wang, H.; Jia, R.; Ye, Z.; He, X.; Wang, K. Colorimetric and Fluorescent Dual-Mode Detection of MicroRNA Based on Duplex-Specific Nuclease Assisted Gold Nanoparticle Amplification. Analyst 2019, 144, 4917–4924. [Google Scholar] [CrossRef]
- Yu, S.; Wang, Y.; Jiang, L.-P.; Bi, S.; Zhu, J.-J. Cascade Amplification-Mediated In Situ Hot-Spot Assembly for MicroRNA Detection and Molecular Logic Gate Operations. Anal. Chem. 2018, 90, 4544–4551. [Google Scholar] [CrossRef]
- Esmaeili-bandboni, A.; Amini, S.M.; Faridi-majidi, R.; Bagheri, J.; Mohammadnejad, J.; Sadroddiny, E. Cross-Linking Gold Nanoparticles Aggregation Method Based on Localised Surface Plasmon Resonance for Quantitative Detection of MiR-155. IET Nanobiotechnol. 2018, 12, 453–458. [Google Scholar] [CrossRef]
- Cai, J.; Ding, L.; Gong, P.; Huang, J. A Colorimetric Detection of MicroRNA-148a in Gastric Cancer by Gold Nanoparticle–RNA Conjugates. Nanotechnology 2019, 31, 095501. [Google Scholar] [CrossRef] [PubMed]
- Mollasalehi, H.; Shajari, E. A Colorimetric Nano-Biosensor for Simultaneous Detection of Prevalent Cancers Using Unamplified Cell-Free Ribonucleic Acid Biomarkers. Bioorg. Chem. 2021, 107, 104605. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Huang, J.; Yang, X.; Yang, Y.; Quan, K.; Xie, N.; Wu, Y.; Ma, C.; Wang, K. Two-Color-Based Nanoflares for Multiplexed MicroRNAs Imaging in Live Cells. Nanotheranostics 2018, 2, 96–105. [Google Scholar] [CrossRef]
- Zhao, J.; Liu, C.; Li, Y.; Ma, Y.; Deng, J.; Li, L.; Sun, J. Thermophoretic Detection of Exosomal MicroRNAs by Nanoflares. J. Am. Chem. Soc. 2020, 142, 4996–5001. [Google Scholar] [CrossRef]
- Hwu, S.; Blickenstorfer, Y.; Tiefenauer, R.F.; Gonnelli, C.; Schmidheini, L.; Lüchtefeld, I.; Hoogenberg, B.-J.; Gisiger, A.B.; Vörös, J. Dark-Field Microwells toward High-Throughput Direct MiRNA Sensing with Gold Nanoparticles. ACS Sens. 2019, 4, 1950–1956. [Google Scholar] [CrossRef] [PubMed]
- Canady, T.D.; Li, N.; Smith, L.D.; Lu, Y.; Kohli, M.; Smith, A.M.; Cunningham, B.T. Digital-Resolution Detection of MicroRNA with Single-Base Selectivity by Photonic Resonator Absorption Microscopy. Proc. Natl. Acad. Sci. USA 2019, 116, 19362–19367. [Google Scholar] [CrossRef]
- Peng, M.; Sun, F.; Na, N.; Ouyang, J. Target-Triggered Assembly of Nanogap Antennas to Enhance the Fluorescence of Single Molecules and Their Application in MicroRNA Detection. Small 2020, 16, 2000460. [Google Scholar] [CrossRef] [PubMed]
- Qu, A.; Sun, M.; Xu, L.; Hao, C.; Wu, X.; Xu, C.; Kotov, N.A.; Kuang, H. Quantitative Zeptomolar Imaging of MiRNA Cancer Markers with Nanoparticle Assemblies. Proc. Natl. Acad. Sci. USA 2019, 116, 3391–3400. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, J.; Zhang, J.; Liu, Y.; Wu, L.; Shen, J.; Zhang, Y.; Hu, Y.; Fan, Q.; Huang, W.; et al. Individual Au-Nanocube Based Plasmonic Nanoprobe for Cancer Relevant MicroRNA Biomarker Detection. ACS Sens. 2017, 2, 1435–1440. [Google Scholar] [CrossRef]
- Slabý, J.; Bocková, M.; Homola, J. Plasmonic Biosensor Based on a Gold Nanostripe Array for Detection of MicroRNA Related to Myelodysplastic Syndromes. Sens. Actuators B Chem. 2021, 347, 130629. [Google Scholar] [CrossRef]
- Kim, W.H.; Lee, J.U.; Song, S.; Kim, S.; Choi, Y.J.; Sim, S.J. A Label-Free, Ultra-Highly Sensitive and Multiplexed SERS Nanoplasmonic Biosensor for MiRNA Detection Using a Head-Flocked Gold Nanopillar. Analyst 2019, 144, 1768–1776. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.-F.; Cheng, L.-X.; Li, M.; Zuo, D.; Zhang, N.; Zhuang, H.-J.; Xie, D.; Zeng, Q.-D.; Hutchison, J.A.; Zhao, Y.-L. Frequency Shift Raman-Based Sensing of Serum MicroRNAs for Early Diagnosis and Discrimination of Primary Liver Cancers. Anal. Chem. 2018, 90, 10144–10151. [Google Scholar] [CrossRef] [PubMed]
- Nie, W.; Wang, Q.; Yang, X.; Zhang, H.; Li, Z.; Gao, L.; Zheng, Y.; Liu, X.; Wang, K. High Sensitivity Surface Plasmon Resonance Biosensor for Detection of MicroRNA Based on Gold Nanoparticles-Decorated Molybdenum Sulfide. Anal. Chim. Acta 2017, 993, 55–62. [Google Scholar] [CrossRef]
- Wang, Q.; Li, Q.; Yang, X.; Wang, K.; Du, S.; Zhang, H.; Nie, Y. Graphene Oxide–Gold Nanoparticles Hybrids-Based Surface Plasmon Resonance for Sensitive Detection of MicroRNA. Biosens. Bioelectron. 2016, 77, 1001–1007. [Google Scholar] [CrossRef]
- Che, C.; Xue, R.; Li, N.; Gupta, P.; Wang, X.; Zhao, B.; Singamaneni, S.; Nie, S.; Cunningham, B.T. Accelerated Digital Biodetection Using Magneto-Plasmonic Nanoparticle-Coupled Photonic Resonator Absorption Microscopy. ACS Nano 2022, 16, 2345–2354. [Google Scholar] [CrossRef]
- Islam, M.N.; Masud, M.K.; Nguyen, N.T.; Gopalan, V.; Alamri, H.R.; Alothman, Z.A.; Al Hossain, M.S.; Yamauchi, Y.; Lamd, A.K.; Shiddiky, M.J. Gold-Loaded Nanoporous Ferric Oxide Nanocubes for Electrocatalytic Detection of MicroRNA at Attomolar Level. Biosens. Bioelectron. 2018, 101, 275–281. [Google Scholar] [CrossRef]
- Lee, J.-H.; Choi, J.-H.; Chueng, S.-T.D.; Pongkulapa, T.; Yang, L.; Cho, H.-Y.; Choi, J.-W.; Lee, K.-B. Nondestructive Characterization of Stem Cell Neurogenesis by a Magneto-Plasmonic Nanomaterial-Based Exosomal MiRNA Detection. ACS Nano 2019, 13, 8793–8803. [Google Scholar] [CrossRef]














Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoang, J.; Tajalli, P.; Omidiyan, M.; Marquez, M.D.; Khantamat, O.; Tuntiwechapikul, W.; Li, C.-H.; Kohlhatkar, A.; Tran, H.-V.; Gunaratne, P.H.; et al. Self-Assembled Monolayers Derived from Positively Charged Adsorbates on Plasmonic Substrates for MicroRNA Delivery: A Review. J. Nanotheranostics 2023, 4, 171-200. https://doi.org/10.3390/jnt4020009
Hoang J, Tajalli P, Omidiyan M, Marquez MD, Khantamat O, Tuntiwechapikul W, Li C-H, Kohlhatkar A, Tran H-V, Gunaratne PH, et al. Self-Assembled Monolayers Derived from Positively Charged Adsorbates on Plasmonic Substrates for MicroRNA Delivery: A Review. Journal of Nanotheranostics. 2023; 4(2):171-200. https://doi.org/10.3390/jnt4020009
Chicago/Turabian StyleHoang, Johnson, Pooria Tajalli, Mina Omidiyan, Maria D. Marquez, Orawan Khantamat, Wirote Tuntiwechapikul, Chien-Hung Li, Arati Kohlhatkar, Hung-Vu Tran, Preethi H. Gunaratne, and et al. 2023. "Self-Assembled Monolayers Derived from Positively Charged Adsorbates on Plasmonic Substrates for MicroRNA Delivery: A Review" Journal of Nanotheranostics 4, no. 2: 171-200. https://doi.org/10.3390/jnt4020009
APA StyleHoang, J., Tajalli, P., Omidiyan, M., Marquez, M. D., Khantamat, O., Tuntiwechapikul, W., Li, C.-H., Kohlhatkar, A., Tran, H.-V., Gunaratne, P. H., & Lee, T. R. (2023). Self-Assembled Monolayers Derived from Positively Charged Adsorbates on Plasmonic Substrates for MicroRNA Delivery: A Review. Journal of Nanotheranostics, 4(2), 171-200. https://doi.org/10.3390/jnt4020009