
Can the Brain’s Thermostatic Mechanism Generate Sleep-Wake and NREM-REM Sleep Cycles? A Nested Doll Model of Sleep-Regulating Processes


Abstract
1. Introduction
2. Results
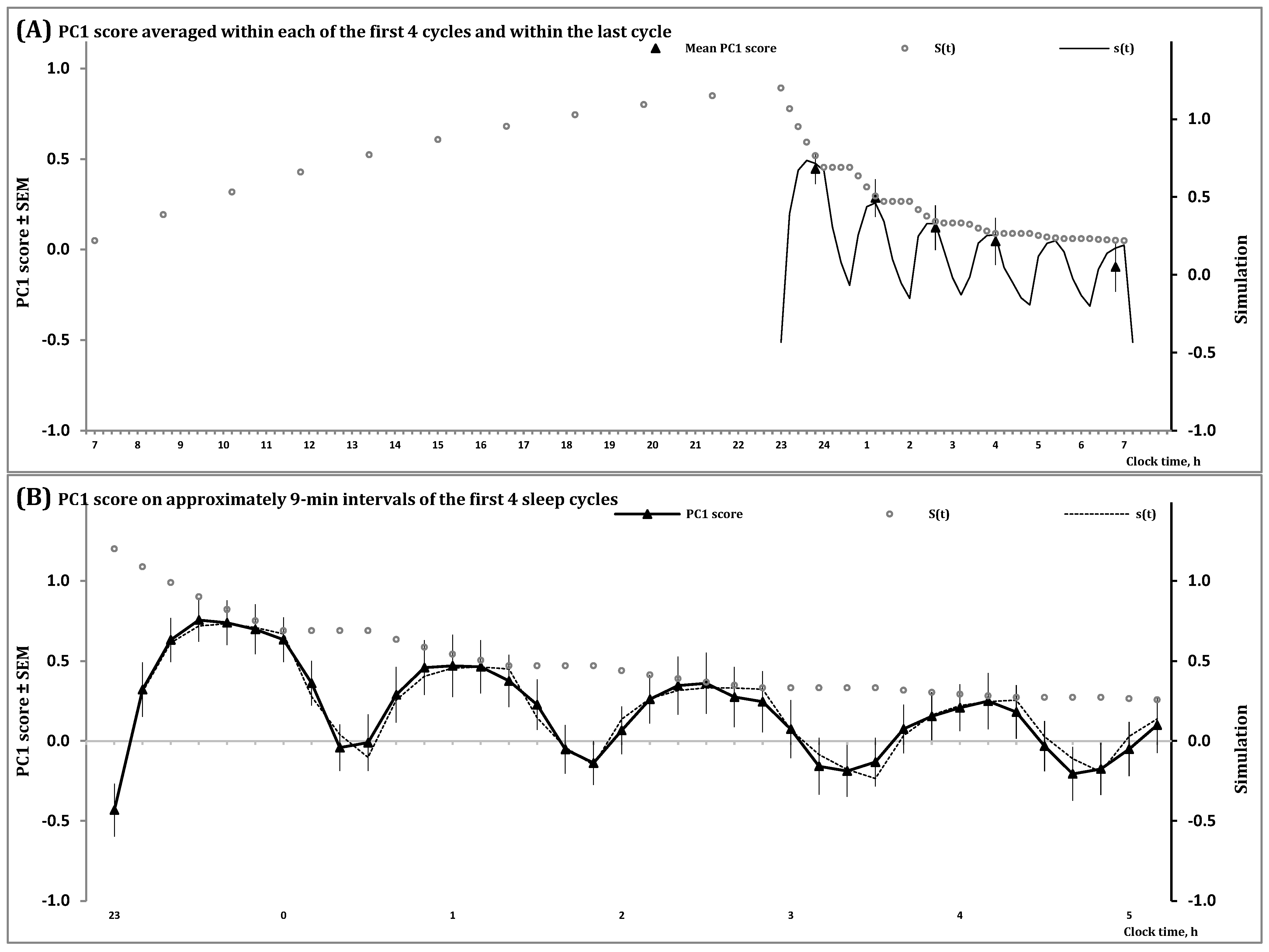
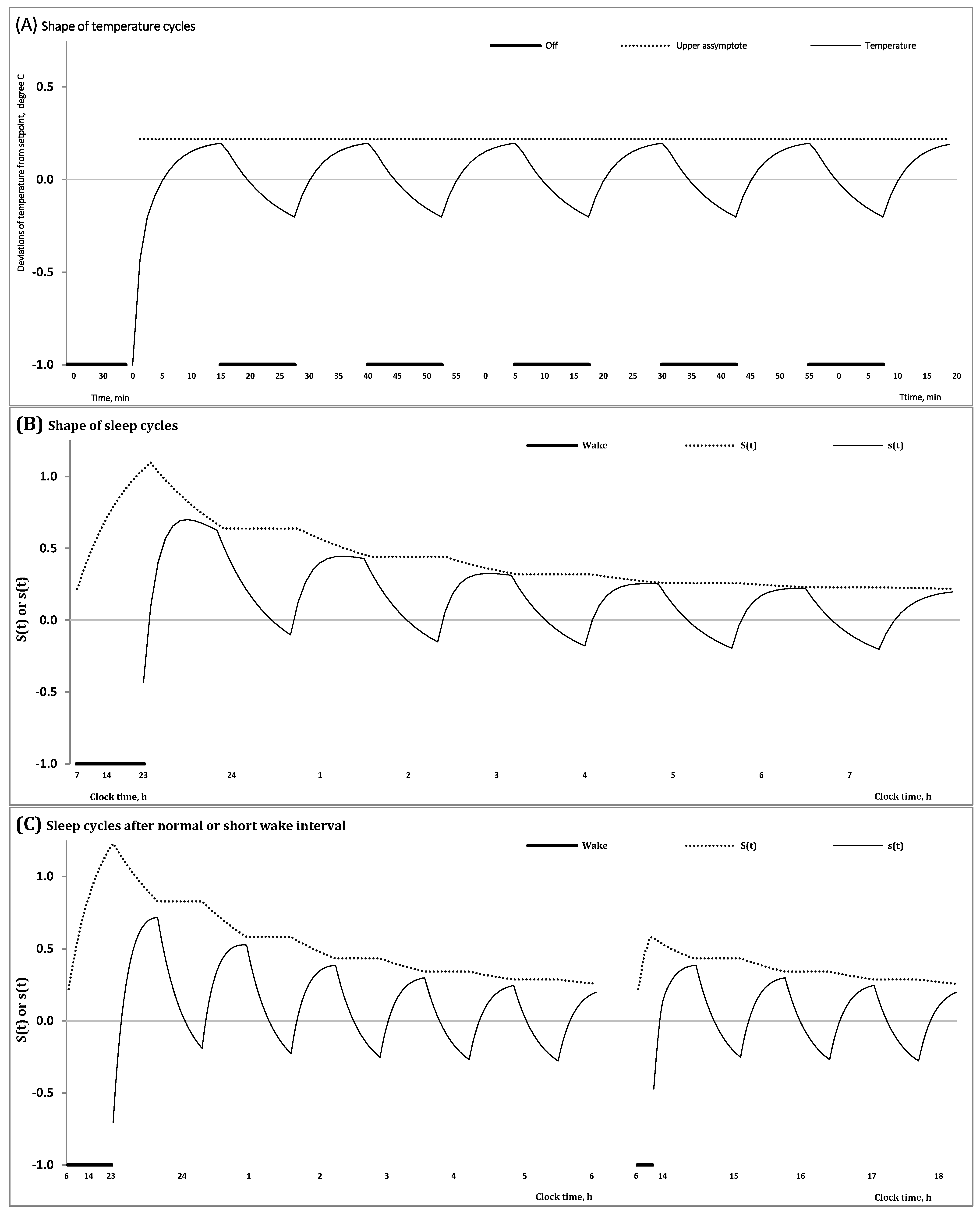
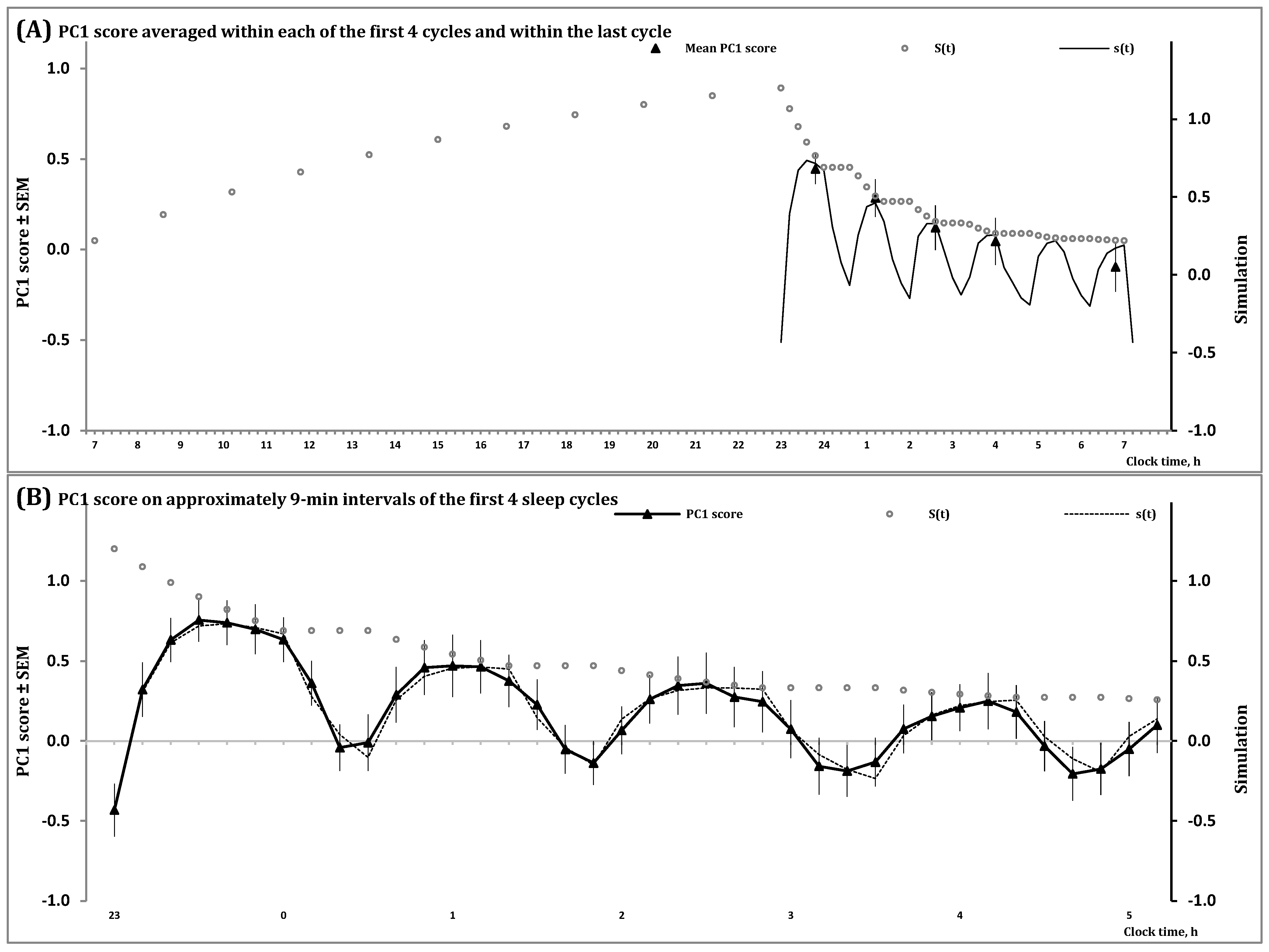
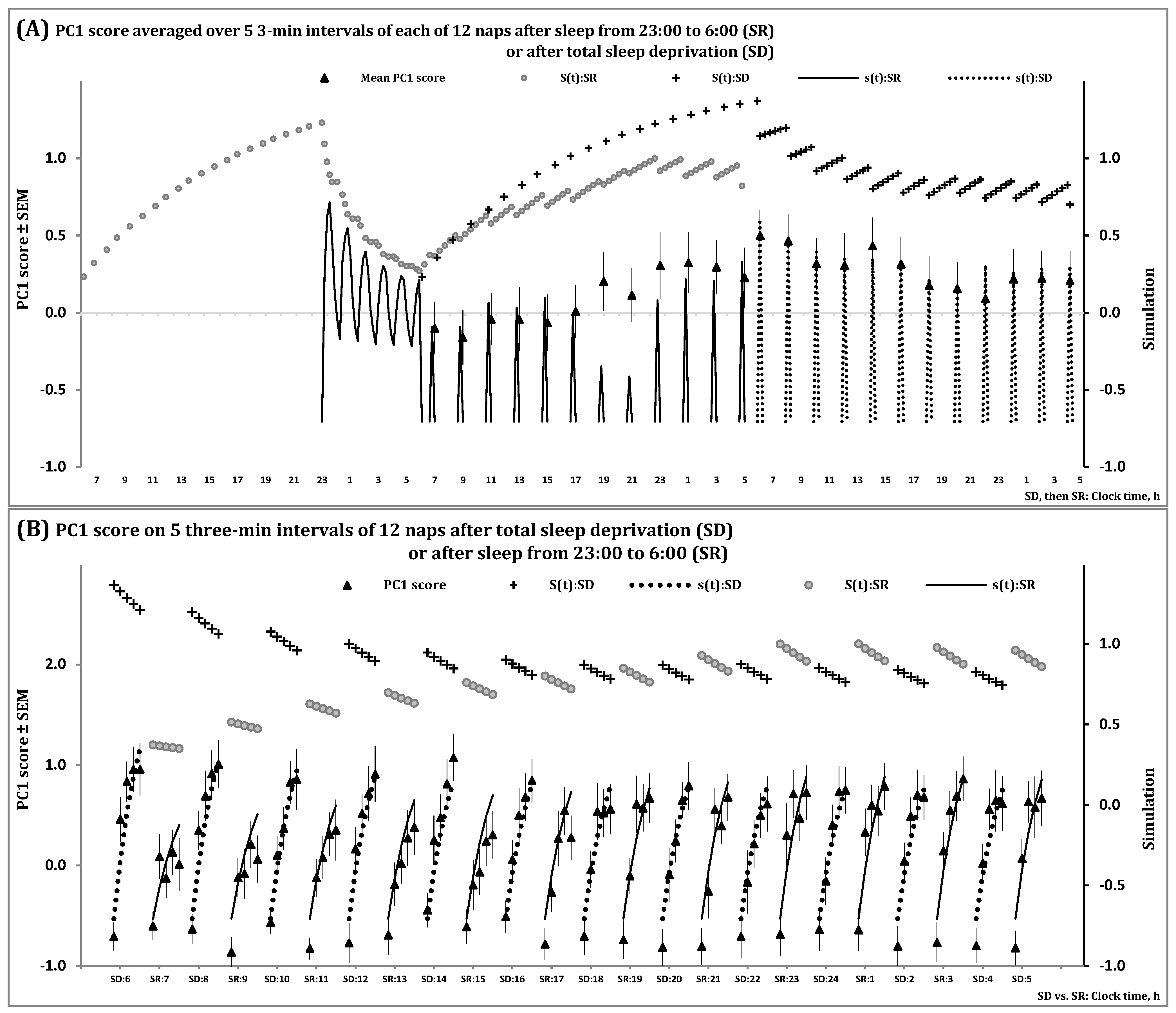
2.1. Results of In Silico Study
- − The duration of the previous wake phase determines the mean level, amplitude and shape of fluctuations of the regulated process on the interval of the first sleep cycles constituting the sleep phase of the sleep-wake cycle;
- − The shape of these cycles cannot closely resemble the alternations of simple exponential curves;
- − The interval with negative values of the regulated process gradually increases in a sequence of sleep cycles during an all-night sleep episode.
2.2. Results of Simulation Study
3. Discussion
3.1. A Model-Based Support for the Hypothesis of Brainstem Thermoregulation Function
3.2. Support from the Studies of Polyphasic Sleep in Laboratory Rodents and Infants
3.3. A Need for Further Support from Studies on Mammal Species with Monophasic Sleep
4. Materials and Methods
4.1. The Proposed Model of the Sleep-Wake Regulatory Processes S and s
4.2. Estimation of Spectral EEG Indicators of Processes S and s
5. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Daan, S.; Beersma, D.G.M.; Borbély, A.A. Timing of human sleep: Recovery process gated by a circadian pacemaker. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1984, 246, R161–R178. [Google Scholar] [CrossRef]
- Magnus, K. Schwingungen: Eine Einführung in Die Theoretische Behandlung von Schwingungsproblemen; BG Teubner Verlagsgesellschaft: Stuttgart, Germany, 1976. [Google Scholar]
- Putilov, A.A. The timing of sleep modelling: Circadian modulation of the homeostatic process. Biol. Rhythm Res. 1995, 26, 1–19. [Google Scholar] [CrossRef]
- Putilov, A.A. Reaction of the endogenous regulatory mechanisms to early weekday wake-ups: A review of its popular explanations in the light of model-based simulations. Front. Netw. Physiol. 2023, 3, 1285658. [Google Scholar] [CrossRef]
- McGinty, D.; Szymusiak, R. Keeping cool: A hypothesis about the mechanisms and functions of slow-wave sleep. Trends Neurosci. 1990, 13, 480–487. [Google Scholar] [CrossRef]
- Wehr, T.A. A brain-warming function for REM sleep. Neurosci. Biobehav. Rev. 1992, 16, 379–397. [Google Scholar] [CrossRef]
- Harding, E.C.; Franks, N.P.; Wisden, W. Sleep and thermoregulation. Curr. Opin. Physiol. 2020, 15, 7–13. [Google Scholar] [CrossRef]
- Parmeggiani, P.L. REM sleep related increase in brain temperature: A physiologic problem. Arch. Ital. Biol. 2007, 145, 13–21. [Google Scholar] [PubMed]
- Szymusiak, R.; McGinty, D. Hypothalamic regulation of sleep and arousal. Ann. N. Y. Acad. Sci. 2008, 1129, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Wisor, J.P.; Rempe, M.J.; Schmidt, M.A.; Moore, M.E.; Clegern, W.C. Sleep slow-wave activity regulates cerebral glycolytic metabolism. Cereb. Cortex 2013, 23, 1978–1987. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.H. The energy allocation function of sleep: A unifying theory of sleep, torpor, and continuous wakefulness. Neurosci. Biobehav. Rev. 2014, 47, 122–153. [Google Scholar] [CrossRef]
- Siegel, J.M. Sleep function: An evolutionary perspective. Lancet Neurol. 2022, 21, 937–946. [Google Scholar] [CrossRef] [PubMed]
- Iranzo, A. How and why do human beings sleep? Lancet Neurol. 2022, 21, 862–864. [Google Scholar] [CrossRef] [PubMed]
- Borbély, A.A. A two process model of sleep regulation. Hum. Neurobiol. 1982, 1, 195–204. [Google Scholar] [PubMed]
- Achermann, P.; Borbély, A.A. Simulation of human sleep: Ultradian dynamics of electroencephalographic slow-wave activity. J. Biol. Rhythms 1990, 5, 141–157. [Google Scholar] [CrossRef] [PubMed]
- Achermann, P.; Dijk, D.J.; Brunner, D.P.; Borbély, A.A. A model of human sleep homeostasis based on EEG slow-wave activity: Quantitative comparison of data and simulations. Brain Res. Bull. 1993, 31, 97–113. [Google Scholar] [CrossRef]
- Putilov, A.A.; Kovalzon, V.M.; Dorokhov, V.B. A relay model of human sleep stages. Eur. Phys. J. Spec. Top. 2023. [Google Scholar] [CrossRef]
- Franken, P.; Tobler, I.; Borbély, A.A. Sleep and waking have a major effect on the 24-hr rhythm of cortical temperature in the rat. J. Biol. Rhythms 1992, 7, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Sela, Y.; Hoekstra, M.M.; Franken, P. Sub-minute prediction of brain temperature based on sleep-wake state in the mouse. elife 2021, 10, e62073. [Google Scholar] [CrossRef]
- Athanasouli, C.; Kalmbach, K.; Booth, V.; Diniz Behn, C.G. NREM-REM alternation complicates transitions from napping to non-napping behavior in a three-state model of sleep-wake regulation. Math. Biosci. 2023, 355, 108929. [Google Scholar] [CrossRef]
- Jenni, O.G.; Borbély, A.A.; Achermann, P. Development of the nocturnal sleep electroencephalogram in human infants. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 286, R528–R538. [Google Scholar] [CrossRef]
- Strughold, H. Rhythmostasis—A fundamental life characteristic aerospace medical aspect. Riv. Med. Aeronaut. Spaz. 1971, 34, 168–175. [Google Scholar]
- Dijk, D.-J.; Brunner, D.P.; Borbély, A.A. Time course of EEG power density during long sleep in humans. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1990, 258, R650–R661. [Google Scholar] [CrossRef]
- Dijk, D.-J.; Brunner, D.P.; Borbély, A.A. EEG power density during recovery sleep in the morning. Electroencephalogr. Clin. Neurophysiol. 1991, 78, 203–214. [Google Scholar] [CrossRef]
- Dijk, D.-J.; Beersma, D.G.M.; Daan, S. EEG power density during nap sleep: Reflection of an hourglass measuring the duration of prior wakefulness. J. Biol. Rhythms 1987, 2, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Greene, R.W.; Frank, M.G. Slow wave activity during sleep: Functional and therapeutic implications. Neuroscientist 2010, 16, 618–633. [Google Scholar] [CrossRef] [PubMed]
- Putilov, A.A. Principal components of electroencephalographic spectrum as markers of opponent processes underlying ultradian sleep cycles. Chronobiol. Int. 2011, 28, 287–299. [Google Scholar] [CrossRef]
- Putilov, A.A. Principal component structure of wake-sleep transition: Quantitative de-scription in multiple sleep latency tests. Somnologie 2010, 14, 234–243. [Google Scholar] [CrossRef]
- Putilov, A.A. Prospects of using electroencephalographic signatures of the chronoregula-tory processes for meaningful, parsimonious and quantitative description of the sleep-wake sub-states. Biol. Rhythm Res. 2011, 42, 181–207. [Google Scholar] [CrossRef]
- Putilov, A.A. The EEG indicators of the dynamic properties of sleep-wake regulating pro-cesses: Comparison of the changes occurring across wake-sleep transition with the effects of prolonged wakefulness. Biol. Rhythm Res. 2013, 44, 621–643. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| # | Process | s(t) | S(t) or su(t) | C(t) |
|---|---|---|---|---|
| 1 | Formula | (4) | (1); (3,5) after exclusion of (2) | (2) in (1) |
| 2 | Name of regulatory mechanism | Ultradian (internal) somnostat | Circadian (external) somnostat | Circadian (clocks) rhythmostat |
| 3 | Purpose of regulation | Control of deviations above and below a setpoint | Tuning this setpoint depending upon a previous wake duration | Entrainment and control of period of S(t) |
| 4 | Period of cycle | Approximately 90 min | Approximately 24 h | Entrained to 24 h |
| 5 | Phases of cycle | Two: one buildup and one decay | Two: one buildup and either decay or unchanged level | Rising and falling limbs of sinusoid |
| 6 | Shape of fluctuations | Inverse exponential function and exponential function | Inverse exponential function and alternation of exponential function with horizontal line | Sine function |
| Process | Parameters | Sleep | Naps | |
|---|---|---|---|---|
| Circadian (external) somnostat, S(t) | Inverse exponential buildup during wake phase (1a,3a) and exponential decay during some of intervals of sleep phase (1b,3b) of circadian (external) somnostat, S(t) (1,3) | Sb (lowest allowed decay) at t1 | 0.22 | 0.23 |
| Sd (highest allowed buildup) at t2 | 1.10 | 1.10 | ||
| Sl (lower asymptote) | 0.20 | 0.20 | ||
| Su (upper asymptote) | 1.41 | 1.41 | ||
| Td (phase constant for decay), h | 1.40 | 1.40 | ||
| Tb (phase constant for buildup), h | 12.01 | 12.01 | ||
| Circadian (clocks) rhythmostat, C(t) | 24 h sine shape modulation C(t) (2) of parameters of buildup (1a) and decay phases (1b) of S(t) (1) | φmax (circadian peak), clock h | 0.00 | 0.00 |
| A (circadian amplitude) | 0.00 | 0.00 | ||
| τ (entrained circadian period), h | 24.00 | 24.00 | ||
| k (twofold impact of circadian term) | 2.00 | 2.00 | ||
| Ultradian (internal) somnostat, s(t) | Inverse exponential buildup during the first (NREM) phases (4a) and exponential decay during the second (REM) phases (4b) of ultradian (internal) somnostat, s(t) (4) | sb (lowest allowed decay) | −0.30 | −0.30 |
| sd (highest allowed buildup) | 0.22 | 0.22 | ||
| sl (lower asymptote) | −0.34 | −0.34 | ||
| su (upper asymptote) = S(t) (3) | 0.70 | 0.70 | ||
| td (phase constant for decay), h | 0.36 | 0.36 | ||
| tb (phase constant for buildup), h | 0.22 | 0.22 | ||
| Initial times for buildup (1a,3a) and decay phases (1b,3b) of S(t) | t2 (sleep onset), clock h | 23.00 | 23.00 | |
| t1 (sleep offset), clock h | 7.00 | 6.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Putilov, A.A. Can the Brain’s Thermostatic Mechanism Generate Sleep-Wake and NREM-REM Sleep Cycles? A Nested Doll Model of Sleep-Regulating Processes. Clocks & Sleep 2024, 6, 97-113. https://doi.org/10.3390/clockssleep6010008
Putilov AA. Can the Brain’s Thermostatic Mechanism Generate Sleep-Wake and NREM-REM Sleep Cycles? A Nested Doll Model of Sleep-Regulating Processes. Clocks & Sleep. 2024; 6(1):97-113. https://doi.org/10.3390/clockssleep6010008
Chicago/Turabian StylePutilov, Arcady A. 2024. "Can the Brain’s Thermostatic Mechanism Generate Sleep-Wake and NREM-REM Sleep Cycles? A Nested Doll Model of Sleep-Regulating Processes" Clocks & Sleep 6, no. 1: 97-113. https://doi.org/10.3390/clockssleep6010008
APA StylePutilov, A. A. (2024). Can the Brain’s Thermostatic Mechanism Generate Sleep-Wake and NREM-REM Sleep Cycles? A Nested Doll Model of Sleep-Regulating Processes. Clocks & Sleep, 6(1), 97-113. https://doi.org/10.3390/clockssleep6010008