
Causal Effects of Gut Microbiota on Sleep-Related Phenotypes: A Two-Sample Mendelian Randomization Study

 and
and
Abstract
:1. Introduction
2. Results
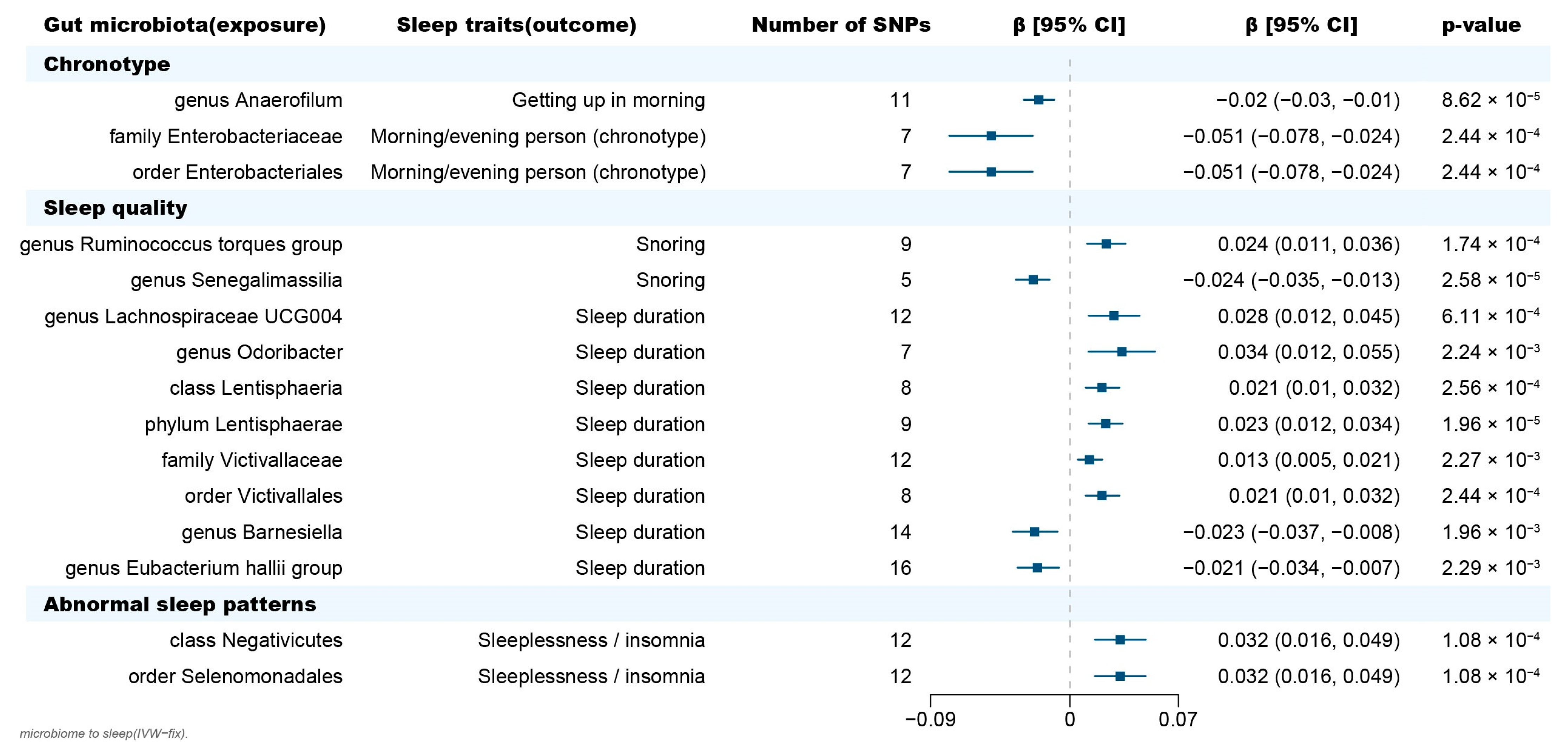
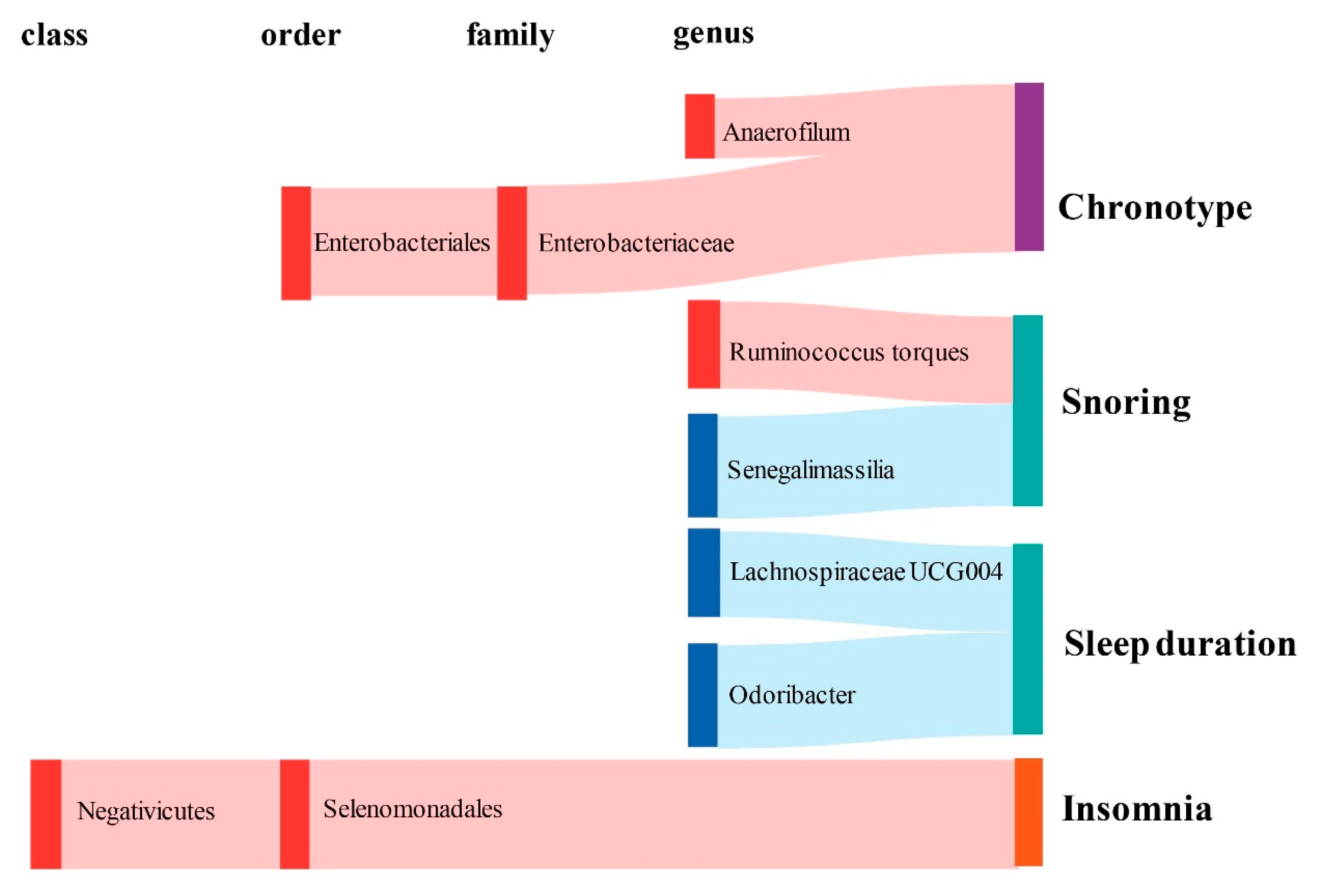
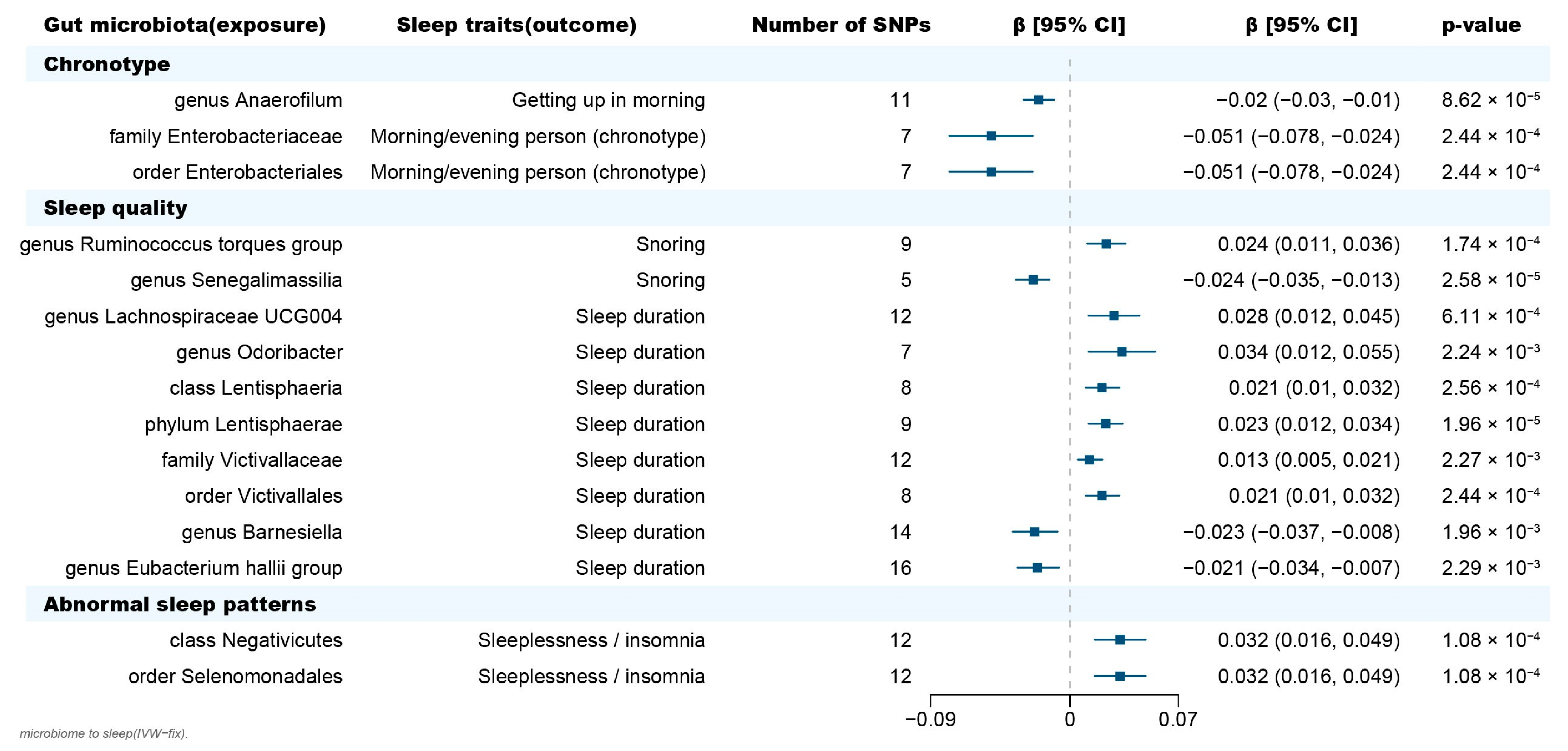
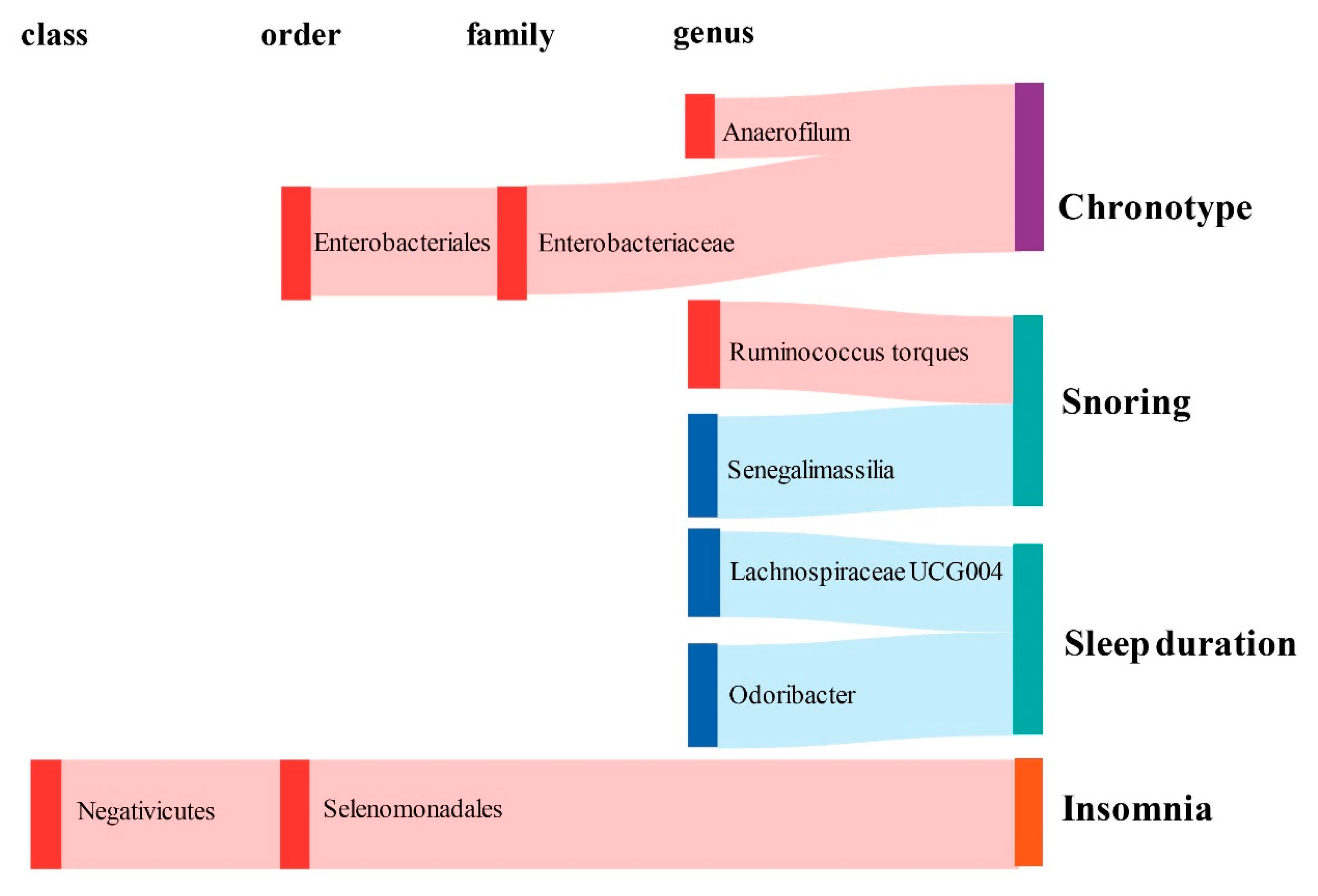
2.1. Causal Effect of Gut Microbiota on Chronotype
2.2. Causal Effect of Gut Microbiota on Sleep Quality
2.3. Causal Effect of Gut Microbiota on Abnormal Sleep Patterns
2.4. Bidirectional Causal Effects between Gut Microbiota and Sleep Phenotypes
3. Discussion
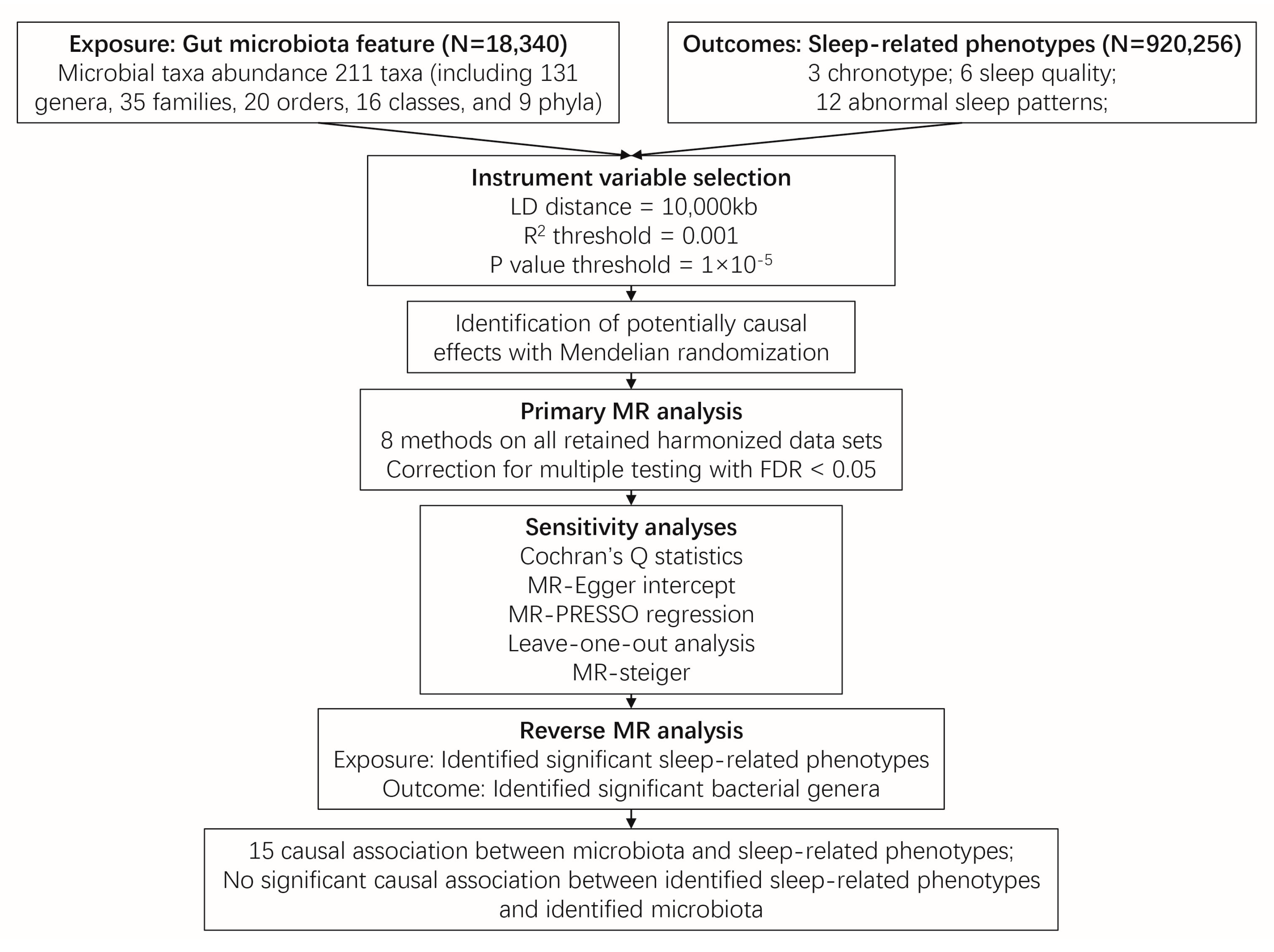
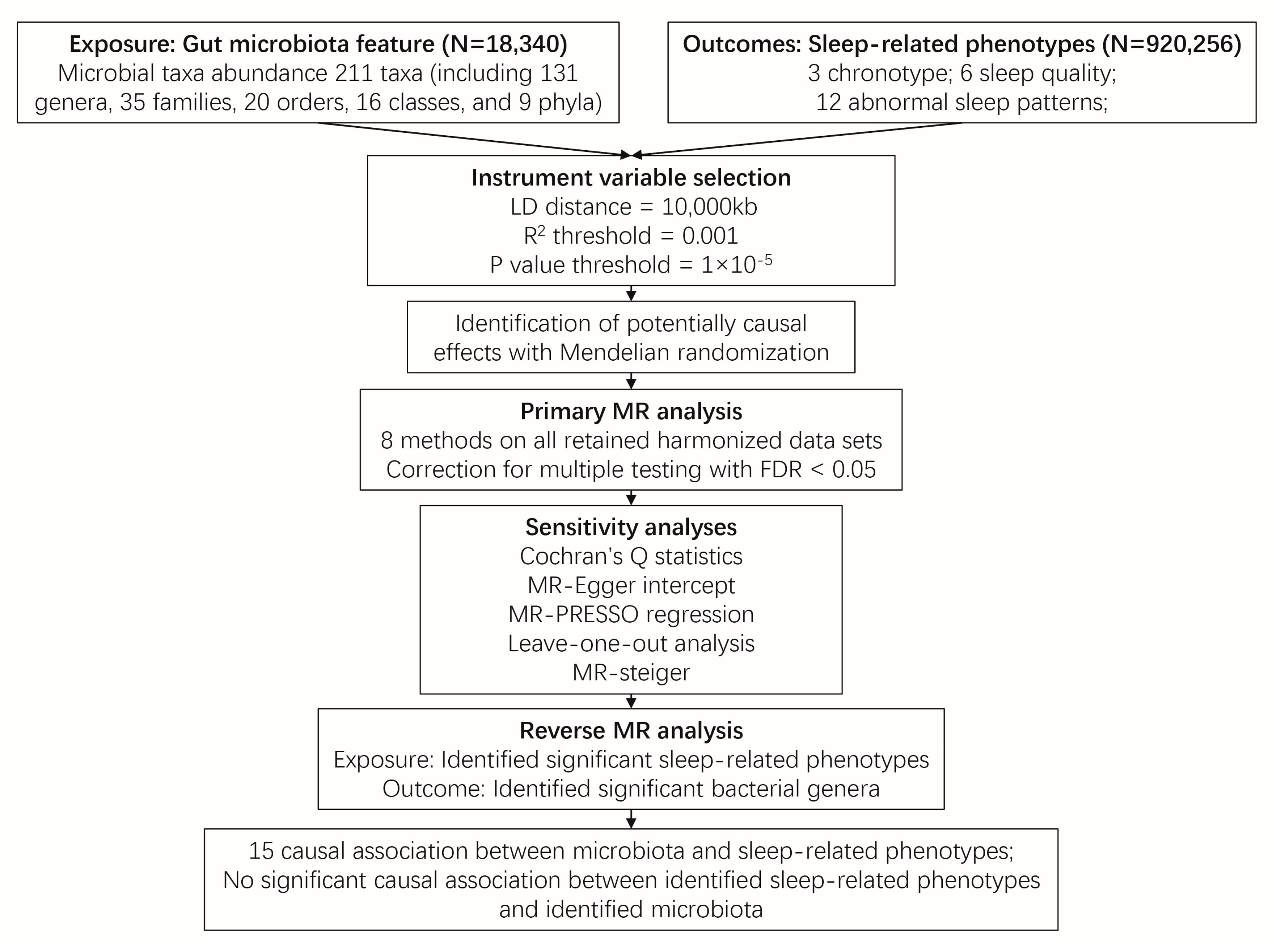
4. Materials and Methods
4.1. Exposure Data
4.2. Outcome Data
4.3. Instrumental Variable (IV) Selection
4.4. MR Analysis
4.5. Sensitivity Analyses
4.6. Reverse MR Analysis
4.7. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Chauhan, P.; Preetam, M. Brain waves and sleep science. Int. J. Eng. Sci. Adv. Res. 2016, 2, 33–36. [Google Scholar]
- Ali, T.; Orr, W.C. Sleep disturbances and inflammatory bowel disease. Inflamm. Bowel Dis. 2014, 20, 1986–1995. [Google Scholar] [CrossRef]
- Zheng, T. Sleep disturbance in heart failure: A concept analysis. In Nursing Forum 2021; Wiley Online Library: Hoboken, NJ, USA, 2021; pp. 710–716. [Google Scholar]
- Dyken, M.E.; Afifi, A.K.; Lin-Dyken, D.C. Sleep-related problems in neurologic diseases. Chest 2012, 141, 528–544. [Google Scholar] [CrossRef] [PubMed]
- Pavlova, M.K.; Latreille, V. Sleep disorders. Am. J. Med. 2019, 132, 292–299. [Google Scholar] [CrossRef]
- Troynikov, O.; Watson, C.G.; Nawaz, N. Sleep environments and sleep physiology: A review. J. Therm. Biol. 2018, 78, 192–203. [Google Scholar] [CrossRef]
- Li, C.; Shang, S. Relationship between Sleep and Hypertension: Findings from the NHANES (2007–2014). Int. J. Environ. Res. Public Health 2021, 18, 7867. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Liu, N.; Guo, D.; Li, B.; Liang, Y.; Huang, L.; Wang, X.; Su, Z.; Zhang, G.; Wang, P. Association between sleep quality and central obesity among southern Chinese reproductive-aged women. BMC Women’s Health 2021, 21, 280. [Google Scholar] [CrossRef]
- Li, F.; Li, Q.; Zhang, Y.; Zhou, X.; Yi, R.; Zhao, X. Effects of xylooligosaccharides on lipid metabolism, inflammation, and gut microbiota in C57BL/6J mice fed a high-fat diet. Front. Pharmacol. 2021, 12, 791614. [Google Scholar] [CrossRef] [PubMed]
- Samy, A.L.; Hairi, N.N.; Low, W.Y. Psychosocial stress, sleep deprivation, and its impact on type II diabetes mellitus: Policies, guidelines, and initiatives from Malaysia. FASEB Bioadv. 2021, 3, 593. [Google Scholar] [CrossRef]
- Golenkov, A.; Filonenko, V.; Filonenko, A. Sleep disorders as one of the indicators of postpartum depression. Zhurnal Nevrol. I Psikhiatrii Im. SS Korsakova 2019, 119, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Ge, L.; Guyatt, G.; Tian, J.; Pan, B.; Chang, Y.; Chen, Y.; Li, H.; Zhang j Li, Y.; Ling, J. Insomnia and risk of mortality from all-cause, cardiovascular disease, and cancer: Systematic review and meta-analysis of prospective cohort studies. Sleep Med. Rev. 2019, 48, 101215. [Google Scholar] [CrossRef]
- Gominak, S.; Stumpf, W. The world epidemic of sleep disorders is linked to vitamin D deficiency. Med. Hypotheses 2012, 79, 132–135. [Google Scholar] [CrossRef]
- Wang, Q.; Chen, B.; Sheng, D.; Yang, J.; Fu, S.; Wang, J.; Zhao, C.; Wang, Y.; Gai, X.; Wang, J. Multiomics analysis reveals aberrant metabolism and immunity linked gut microbiota with insomnia. Microbiol. Spectr. 2022, 10, e00998-22. [Google Scholar] [CrossRef]
- Matenchuk, B.A.; Mandhane, P.J.; Kozyrskyj, A.L. Sleep, circadian rhythm, and gut microbiota. Sleep Med. Rev. 2020, 53, 101340. [Google Scholar] [CrossRef]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the healthy gut microbiota composition? A changing ecosystem across age, environment, diet, and diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef]
- Davey Smith, G.; Ebrahim, S. ‘Mendelian randomization’: Can genetic epidemiology contribute to understanding environmental determinants of disease? Int. J. Epidemiol. 2003, 32, 1–22. [Google Scholar] [CrossRef]
- Davey Smith, G.; Hemani, G. Mendelian randomization: Genetic anchors for causal inference in epidemiological studies. Hum. Mol. Genet. 2014, 23, R89–R98. [Google Scholar] [CrossRef]
- Sekula, P.; Fabiola Del Greco, M.; Pattaro, C.; Köttgen, A. Mendelian randomization as an approach to assess causality using observational data. J. Am. Soc. Nephrol. 2016, 27, 3253–3265. [Google Scholar] [CrossRef]
- Wang, J.; Kurilshikov, A.; Radjabzadeh, D.; Turpin, W.; Croitoru, K.; Bonder, M.J.; Jackson, M.A.; Medina-Gomez, C.; Frost, F.; Homuth, G. Meta-Analysis of Human Genome-Microbiome Association Studies: The MiBioGen Consortium Initiative; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Kurilshikov, A.; Medina-Gomez, C.; Bacigalupe, R.; Radjabzadeh, D.; Wang, J.; Demirkan, A.; Le Roy, C.I.; Raygoza Garay, J.A.; Finnicum, C.T.; Liu, X. Large-scale association analyses identify host factors influencing human gut microbiome composition. Nat. Genet. 2021, 53, 156–165. [Google Scholar] [CrossRef]
- Bowden, J.; Davey Smith, G.; Haycock, P.C.; Burgess, S. Consistent estimation in Mendelian randomization with some invalid instruments using a weighted median estimator. Genet. Epidemiol. 2016, 40, 304–314. [Google Scholar] [CrossRef]
- Agrawal, R.; Ajami, N.J.; Malhotra, S.; Chen, L.; White, D.L.; Sharafkhaneh, A.; Hoffman, K.L.; Graham, D.Y.; El-Serag, H.B.; Petrosino, J.F. Habitual sleep duration and the colonic mucosa-associated gut microbiota in humans—A pilot study. Clocks Sleep 2021, 3, 387–397. [Google Scholar] [CrossRef]
- Anderson, J.R.; Carroll, I.; Azcarate-Peril, M.A.; Rochette, A.D.; Heinberg, L.J.; Peat, C.; Steffen, K.; Manderino, L.M.; Mitchell, J.; Gunstad, J. A preliminary examination of gut microbiota, sleep, and cognitive flexibility in healthy older adults. Sleep Med. 2017, 38, 104–107. [Google Scholar] [CrossRef]
- El Aidy, S.; Bolsius, Y.G.; Raven, F.; Havekes, R. A brief period of sleep deprivation leads to subtle changes in mouse gut microbiota. J. Sleep Res. 2020, 29, e12920. [Google Scholar] [CrossRef]
- Liu, Z.; Wei, Z.-Y.; Chen, J.; Chen, K.; Mao, X.; Liu, Q.; Sun, Y.; Zhang, Z.; Zhang, Y.; Dan, Z. Acute sleep-wake cycle shift results in community alteration of human gut microbiome. Msphere 2020, 5, e00914-19. [Google Scholar] [CrossRef]
- Maki, K.A.; Burke, L.A.; Calik, M.W.; Watanabe-Chailland, M.; Sweeney, D.; Romick-Rosendale, L.E.; Green, S.J.; Fink, A.M. Sleep fragmentation increases blood pressure and is associated with alterations in the gut microbiome and fecal metabolome in rats. Physiol. Genom. 2020, 52, 280–292. [Google Scholar] [CrossRef]
- Valentini, F.; Evangelisti, M.; Arpinelli, M.; Di Nardo, G.; Borro, M.; Simmaco, M.; Villa, M.P. Gut microbiota composition in children with obstructive sleep apnoea syndrome: A pilot study. Sleep Med. 2020, 76, 140–147. [Google Scholar] [CrossRef]
- Moreno-Indias, I.; Torres, M.; Montserrat, J.M.; Sanchez-Alcoholado, L.; Cardona, F.; Tinahones, F.J.; Gozal, D.; Poroyko, V.A.; Navajas, D.; Queipo-Ortuño, M.I. Intermittent hypoxia alters gut microbiota diversity in a mouse model of sleep apnoea. Eur. Respir. J. 2015, 45, 1055–1065. [Google Scholar] [CrossRef]
- Kim, K.S.; Min, H.J. Correlations between the adenotonsillar microbiome and clinical characteristics of pediatric patients with snoring. Clin. Exp. Otorhinolaryngol. 2021, 14, 295–302. [Google Scholar] [CrossRef]
- Han, M.; Yuan, S.; Zhang, J. The interplay between sleep and gut microbiota. Brain Res. Bull. 2022, 180, 131–146. [Google Scholar] [CrossRef]
- Huang, J.; Zuber, V.; Matthews, P.M.; Elliott, P.; Tzoulaki, J.; Dehghan, A. Sleep, major depressive disorder, and Alzheimer disease: A Mendelian randomization study. Neurology 2020, 95, e1963–e1970. [Google Scholar] [CrossRef]
- Gao, R.-C.; Sang, N.; Jia, C.-Z.; Zhang, M.-Y.; Li, B.-H.; Wei, M.; Wu, G.-C. Association Between Sleep Traits and Rheumatoid Arthritis: A Mendelian Randomization Study. Front. Public Health 2022, 10, 940161. [Google Scholar] [CrossRef]
- Sang, N.; Gao, R.-C.; Zhang, M.-Y.; Wu, Z.-Z.; Wu, Z.-G.; Wu, G.-C. Causal relationship between sleep traits and risk of systemic lupus erythematosus: A two-sample Mendelian randomization study. Front. Immunol. 2022, 13, 918749. [Google Scholar] [CrossRef] [PubMed]
- Short, M.A.; Blunden, S.; Rigney, G.; Matricciani, L.; Coussens, S.; Reynolds, C.M.; Galland, B. Cognition and objectively measured sleep duration in children: A systematic review and meta-analysis. Sleep Health 2018, 4, 292–300. [Google Scholar] [CrossRef]
- Maski, K.P.; Kothare, S.V. Sleep deprivation and neurobehavioral functioning in children. Int. J. Psychophysiol. 2013, 89, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Chaput, J.-P.; Wong, S.L.; Michaud, I. Duration and quality of sleep among Canadians aged 18 to 79. Health Rep. 2017, 28, 28. [Google Scholar]
- Chen, Y.; Tan, F.; Wei, L.; Li, X.; Lyu, Z.; Feng, X.; Wen, Y.; Guo, L.; He, J.; Dai, M. Sleep duration and the risk of cancer: A systematic review and meta-analysis including dose–response relationship. BMC Cancer 2018, 18, 1149. [Google Scholar] [CrossRef]
- Engeda, J.; Mezuk, B.; Ratliff, S.; Ning, Y. Association between duration and quality of sleep and the risk of pre-diabetes: Evidence from NHANES. Diabet. Med. 2013, 30, 676–680. [Google Scholar] [CrossRef]
- Lou, P.; Chen, P.; Zhang, L.; Zhang, P.; Yu, J.; Zhang, N.; Wu, H.; Zhao, J. Relation of sleep quality and sleep duration to type 2 diabetes: A population-based cross-sectional survey. BMJ Open 2012, 2, e000956. [Google Scholar] [CrossRef]
- Peter-Derex, L.; Yammine, P.; Bastuji, H.; Croisile, B. Sleep and Alzheimer’s disease. Sleep Med. Rev. 2015, 19, 29–38. [Google Scholar] [CrossRef]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The role of short-chain fatty acids from gut microbiota in gut-brain communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef]
- Morin, C.M.; Drake, C.L.; Harvey, A.G.; Krystal, A.D.; Manber, R.; Riemann, D.; Spiegelhalder, K. Insomnia disorder. Nat. Rev. Dis. Primers 2015, 1, 15026. [Google Scholar] [CrossRef]
- Szentirmai, É.; Millican, N.S.; Massie, A.R.; Kapás, L. Butyrate, a metabolite of intestinal bacteria, enhances sleep. Sci. Rep. 2019, 9, 7035. [Google Scholar] [CrossRef] [PubMed]
- Cornejo-Pareja, I.; Ruiz-Limón, P.; Gómez-Pérez, A.M.; Molina-Vega, M.; Moreno-Indias, I.; Tinahones, F.J. Differential microbial pattern description in subjects with autoimmune-based thyroid diseases: A pilot study. J. Pers. Med. 2020, 10, 192. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, W.; Liu, M.; Zhang, Q.; Lin, Z.; Jia, M.; Liu, D.; Lin, L. Imbalance of Autophagy and Apoptosis Induced by Oxidative Stress May Be Involved in Thyroid Damage Caused by Sleep Deprivation in Rats. Oxidative Med. Cell. Longev. 2021, 2021, 5645090. [Google Scholar] [CrossRef]
- Baglioni, C.; Spiegelhalder, K.; Lombardo, C.; Riemann, D. Sleep and emotions: A focus on insomnia. Sleep Med. Rev. 2010, 14, 227–238. [Google Scholar] [CrossRef]
- Cardinali, D.P.; Srinivasan, V.; Brzezinski, A.; Brown, G.M. Melatonin and its analogs in insomnia and depression. J. Pineal Res. 2012, 52, 365–375. [Google Scholar] [CrossRef]
- Xia, S.; Gao, W.; Li, Y.; Ma, J.; Gong, S.; Gao, Z.; Tang, W.; Tian, W.; Tang, S. Effects of melatonin on intestinal function and bacterial compositions in sucking piglets. J. Anim. Physiol. Anim. Nutr. 2022, 106, 1139–1148. [Google Scholar] [CrossRef] [PubMed]
- Fischer, D.; Lombardi, D.A.; Marucci-Wellman, H.; Roenneberg, T. Chronotypes in the US–influence of age and sex. PLoS ONE 2017, 12, e0178782. [Google Scholar] [CrossRef]
- Jones, S.E.; Lane, J.M.; Wood, A.R.; van Hees, V.T.; Tyrrell, J.; Beaumont, R.N.; Jeffries, A.R.; Dashti, H.S.; Hillsdon, M.; Ruth, K.S. Genome-wide association analyses of chronotype in 697,828 individuals provides insights into circadian rhythms. Nat. Commun. 2019, 10, 343. [Google Scholar] [CrossRef]
- Ekiz Erim, S.; Sert, H. The relationship between chronotype and obesity: A systematic review. Chronobiol. Int. 2023, 40, 529–541. [Google Scholar] [CrossRef]
- De Amicis, R.; Galasso, L.; Leone, A.; Vignati, L.; De Carlo, G.; Foppiani, A.; Montaruli, A.; Roveda, E.; Cè, E.; Esposito, F.; et al. Is Abdominal Fat Distribution Associated with Chronotype in Adults Independently of Lifestyle Factors? Nutrients 2020, 12, 592. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.; Zhang, J.; Wei, L.; Ma, X.; Zhang, W.; Nie, C. Cottonseed meal fermented by Candida tropical reduces the fat deposition in white-feather broilers through cecum bacteria-host metabolic cross-talk. Appl. Microbiol. Biotechnol. 2020, 104, 4345–4357. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L. The gut microbiota and obesity: From correlation to causality. Nat. Rev. Microbiol. 2013, 11, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Buxton, O.M.; Cain, S.W.; O’Connor, S.P.; Porter, J.H.; Duffy, J.F.; Wang, W.; Czeisler, C.A.; Shea, S.A. Adverse metabolic consequences in humans of prolonged sleep restriction combined with circadian disruption. Sci. Transl. Med. 2012, 4, 129ra143. [Google Scholar] [CrossRef]
- Scheer, F.A.; Hilton, M.F.; Mantzoros, C.S.; Shea, S.A. Adverse metabolic and cardiovascular consequences of circadian misalignment. Proc. Natl. Acad. Sci. USA 2009, 106, 4453–4458. [Google Scholar] [CrossRef]
- Verhulst, S.L.; Aerts, L.; Jacobs, S.; Schrauwen, N.; Haentjens, D.; Claes, R.; Vaerenberg, H.; Van Gaal, L.F.; De Backer, W.A.; Desager, K.N. Sleep-disordered breathing, obesity, and airway inflammation in children and adolescents. Chest 2008, 134, 1169–1175. [Google Scholar] [CrossRef]
- Sakamoto, N.; Gozal, D.; Smith, D.L.; Yang, L.; Morimoto, N.; Wada, H.; Maruyama, K.; Ikeda, A.; Suzuki, Y.; Nakayama, M.; et al. Sleep Duration, Snoring Prevalence, Obesity, and Behavioral Problems in a Large Cohort of Primary School Students in Japan. Sleep 2017, 40, zsw082. [Google Scholar] [CrossRef]
- Ma, Y.; Peng, L.; Kou, C.; Hua, S.; Yuan, H. Associations of Overweight, Obesity and Related Factors with Sleep-Related Breathing Disorders and Snoring in Adolescents: A Cross-Sectional Survey. Int. J. Environ. Res. Public Health 2017, 14, 194. [Google Scholar] [CrossRef]
- Liu, R.; Hong, J.; Xu, X.; Feng, Q.; Zhang, D.; Gu, Y.; Shi, J.; Zhao, S.; Liu, W.; Wang, X. Gut microbiome and serum metabolome alterations in obesity and after weight-loss intervention. Nat. Med. 2017, 23, 859–868. [Google Scholar] [CrossRef]
- Le Chatelier, E.; Nielsen, T.; Qin, J.; Prifti, E.; Hildebrand, F.; Falony, G.; Almeida, M.; Arumugam, M.; Batto, J.-M.; Kennedy, S. Richness of human gut microbiome correlates with metabolic markers. Nature 2013, 500, 541–546. [Google Scholar] [CrossRef]
- Peterson, C.T.; Sharma, V.; Elmén, L.; Peterson, S.N. Immune homeostasis, dysbiosis and therapeutic modulation of the gut microbiota. Clin. Exp. Immunol. 2015, 179, 363–377. [Google Scholar] [CrossRef]
- Adamberg, K.; Adamberg, S.; Ernits, K.; Larionova, A.; Voor, T.; Jaagura, M.; Visnapuu, T.; Alamäe, T. Composition and metabolism of fecal microbiota from normal and overweight children are differentially affected by melibiose, raffinose and raffinose-derived fructans. Anaerobe 2018, 52, 100–110. [Google Scholar] [CrossRef]
- Sudlow, C.; Gallacher, J.; Allen, N.; Beral, V.; Burton, P.; Danesh, J.; Downey, P.; Elliott, P.; Green, J.; Landray, M. UK biobank: An open access resource for identifying the causes of a wide range of complex diseases of middle and old age. PLoS Med. 2015, 12, e1001779. [Google Scholar] [CrossRef]
- Battram, T.; Yousefi, P.; Crawford, G.; Prince, C.; Babaei, M.S.; Sharp, G.; Hatcher, C.; Vega-Salas, M.J.; Khodabakhsh, S.; Whitehurst, O. The EWAS Catalog: A database of epigenome-wide association studies. Wellcome Open Res. 2022, 7, 41. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.E.; Tyrrell, J.; Wood, A.R.; Beaumont, R.N.; Ruth, K.S.; Tuke, M.A.; Yaghootkar, H.; Hu, Y.; Teder-Laving, M.; Hayward, C. Genome-wide association analyses in 128,266 individuals identifies new morningness and sleep duration loci. PLoS Genet. 2016, 12, e1006125. [Google Scholar] [CrossRef] [PubMed]
- Richmond, R.C.; Anderson, E.L.; Dashti, H.S.; Jones, S.E.; Lane, J.M.; Strand, L.B.; Brumpton, B.; Rutter, M.K.; Wood, A.R.; Straif, K. Investigating causal relations between sleep traits and risk of breast cancer in women: Mendelian randomisation study. BMJ 2019, 365, l2327. [Google Scholar] [CrossRef] [PubMed]
- Jansen, P.R.; Watanabe, K.; Stringer, S.; Skene, N.; Bryois, J.; Hammerschlag, A.R.; de Leeuw, C.A.; Benjamins, J.S.; Muñoz-Manchado, A.B.; Nagel, M. Genome-wide analysis of insomnia in 1,331,010 individuals identifies new risk loci and functional pathways. Nat. Genet. 2019, 51, 394–403. [Google Scholar] [CrossRef]
- Sateia, M.J. International classification of sleep disorders. Chest 2014, 146, 1387–1394. [Google Scholar] [CrossRef] [PubMed]
- Strausz, S.; Ruotsalainen, S.; Ollila, H.M.; Karjalainen, J.; Kiiskinen, T.; Reeve, M.; Kurki, M.; Mars, N.; Havulinna, A.S.; Luonsi, E. Genetic analysis of obstructive sleep apnoea discovers a strong association with cardiometabolic health. Eur. Respir. J. 2021, 57, 2003091. [Google Scholar] [CrossRef]
- Burgess, S.; Thompson, S.G. Bias in causal estimates from Mendelian randomization studies with weak instruments. Stat. Med. 2011, 30, 1312–1323. [Google Scholar] [CrossRef]
- Burgess, S.; Dudbridge, F.; Thompson, S.G. Combining information on multiple instrumental variables in Mendelian randomization: Comparison of allele score and summarized data methods. Stat. Med. 2016, 35, 1880–1906. [Google Scholar] [CrossRef]
- Pierce, B.L.; Burgess, S. Efficient design for Mendelian randomization studies: Subsample and 2-sample instrumental variable estimators. Am. J. Epidemiol. 2013, 178, 1177–1184. [Google Scholar] [CrossRef] [PubMed]
- Bowden, J.; Davey Smith, G.; Burgess, S. Mendelian randomization with invalid instruments: Effect estimation and bias detection through Egger regression. Int. J. Epidemiol. 2015, 44, 512–525. [Google Scholar] [CrossRef] [PubMed]
- Hartwig, F.P.; Davey Smith, G.; Bowden, J. Robust inference in summary data Mendelian randomization via the zero modal pleiotropy assumption. Int. J. Epidemiol. 2017, 46, 1985–1998. [Google Scholar] [CrossRef]
- Verbanck, M.; Chen, C.-Y.; Neale, B.; Do, R. Detection of widespread horizontal pleiotropy in causal relationships inferred from Mendelian randomization between complex traits and diseases. Nat. Genet. 2018, 50, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Hemani, G.; Tilling, K.; Davey Smith, G. Orienting the causal relationship between imprecisely measured traits using GWAS summary data. PLoS Genet. 2017, 13, e1007081. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
| Exposure | Outcome | Exposure ID | Outcome ID | Q | Q df | Q Pval | I2 | Sample Size | Intercepts [95% CI] | Pval | Global Test RSSobs | Global Test Pvalue | Distortion Test Pvalue | Distortion Coefficient | Correct Causal Direction | Steiger Test |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| class Negativicutes id.2164 | Sleeplessness/insomnia || id:ukb-b-3957 | ebi-a-GCST90016922 | ukb-b-3957 | 8.09 | 11.0 | 0.71 | −0.36 | 14,306 | −0.001 (−0.004, 0.003) | 0.717 | 9.73 | 0.768 | NA | NA | TRUE | 3.77 × 10−56 |
| order Selenomonadales id.2165 | Sleeplessness/insomnia || id:ukb-b-3957 | ebi-a-GCST90017107 | ukb-b-3957 | 8.09 | 11.0 | 0.71 | −0.36 | 14,306 | −0.001 (−0.004, 0.003) | 0.717 | 9.73 | 0.768 | NA | NA | TRUE | 3.77 × 10−56 |
| class Lentisphaeria id.2250 | Sleep duration || id:ukb-b-4424 | ebi-a-GCST90016918 | ukb-b-4424 | 10.40 | 7.0 | 0.17 | 0.33 | 14,306 | 0.003 (−0.004, 0.01) | 0.368 | 13.60 | 0.273 | NA | NA | TRUE | 4.47 × 10−42 |
| family Victivallaceae id.2255 | Sleep duration || id:ukb-b-4424 | ebi-a-GCST90016958 | ukb-b-4424 | 15.80 | 11.0 | 0.15 | 0.30 | 14,306 | 0.002 (−0.004, 0.009) | 0.524 | 18.90 | 0.212 | NA | NA | TRUE | 4.91 × 10−62 |
| genus Barnesiella id.944 | Sleep duration || id:ukb-b-4424 | ebi-a-GCST90016969 | ukb-b-4424 | 11.10 | 13.0 | 0.60 | −0.17 | 14,306 | −0.003 (−0.007, 0.001) | 0.155 | 12.90 | 0.605 | NA | NA | TRUE | 2.55 × 10−73 |
| genus Eubacterium hallii group id.11338 | Sleep duration || id:ukb-b-4424 | ebi-a-GCST90017000 | ukb-b-4424 | 11.80 | 15.0 | 0.70 | −0.27 | 14,306 | 0.001 (−0.001, 0.003) | 0.344 | 13.20 | 0.738 | NA | NA | TRUE | 1.30 × 10−67 |
| genus Lachnospiraceae UCG004 id.11324 | Sleep duration || id:ukb-b-4424 | ebi-a-GCST90017026 | ukb-b-4424 | 15.40 | 11.0 | 0.17 | 0.28 | 14,306 | 0.004 (0, 0.008) | 0.08 | 18.30 | 0.228 | NA | NA | TRUE | 1.27 × 10−54 |
| genus Odoribacter id.952 | Sleep duration || id:ukb-b-4424 | ebi-a-GCST90017034 | ukb-b-4424 | 4.57 | 6.0 | 0.60 | −0.31 | 14,306 | 0.002 (−0.004, 0.007) | 0.55 | 6.36 | 0.610 | NA | NA | TRUE | 1.53 × 10−39 |
| order Victivallales id.2254 | Sleep duration || id:ukb-b-4424 | ebi-a-GCST90017109 | ukb-b-4424 | 10.40 | 7.0 | 0.17 | 0.33 | 14,306 | 0.003 (−0.004, 0.01) | 0.37 | 13.60 | 0.273 | NA | NA | TRUE | 4.47 × 10−42 |
| phylum Lentisphaerae id.2238 | Sleep duration || id:ukb-b-4424 | ebi-a-GCST90017115 | ukb-b-4424 | 11.90 | 8.0 | 0.15 | 0.33 | 14,306 | 0.003 (−0.004, 0.01) | 0.44 | 15.10 | 0.293 | NA | NA | TRUE | 2.42 × 10−46 |
| genus Ruminococcus torques group id.14377 | Snoring || id:ukb-b-17400 | ebi-a-GCST90017066 | ukb-b-17400 | 12.40 | 8.0 | 0.13 | 0.36 | 14,306 | 0.002 (−0.001, 0.004) | 0.25 | 15.30 | 0.210 | NA | NA | TRUE | 2.18 × 10−54 |
| genus Senegalimassilia id.11160 | Snoring || id:ukb-b-17400 | ebi-a-GCST90017068 | ukb-b-17400 | 4.70 | 4.0 | 0.32 | 0.15 | 14,306 | 0 (−0.004, 0.005) | 0.89 | 7.18 | 0.465 | NA | NA | TRUE | 1.11 × 10−32 |
| genus Anaerofilum id.2053 | Getting up in morning || id:ukb-b-2772 | ebi-a-GCST90016965 | ukb-b-2772 | 13.90 | 1.0 | 0.18 | 0.28 | 14,306 | −0.002 (−0.009, 0.005) | 0.52 | 16.80 | 0.217 | NA | NA | TRUE | 5.87 × 10−53 |
| family Enterobacteriaceae id.3469 | Morning/evening person (chronotype) || id:ukb-b-4956 | ebi-a-GCST90016936 | ukb-b-4956 | 9.82 | 6.0 | 0.13 | 0.39 | 14,306 | −0.008 (−0.023, 0.007) | 0.29 | 13.40 | 0.163 | NA | NA | TRUE | 1.33 × 10−44 |
| order Enterobacteriales id.3468 | Morning/evening person (chronotype) || id:ukb-b-4956 | ebi-a-GCST90017098 | ukb-b-4956 | 9.82 | 6.0 | 0.13 | 0.39 | 14,306 | −0.008 (−0.023, 0.007) | 0.29 | 13.40 | 0.163 | NA | NA | TRUE | 1.33 × 10−44 |
| Phenotype | Consortium | Date | Sample Size (Case) | nSNPs | Population | Gender | GWAS_ID |
|---|---|---|---|---|---|---|---|
| Disorder of the sleep-wake schedule | FinnGen | 2021 | 216,354 (190) | 16,380,459 | European | Mixed | finn-b-F5_SLEEPWAKE |
| Sleep apnoea | FinnGen | 2021 | 217,965 (16,761) | 16,380,465 | European | Mixed | finn-b-G6_SLEEPAPNO |
| Sleep disorders | FinnGen | 2021 | 216,454 (2628) | 16,380,458 | European | Mixed | finn-b-F5_SLEEP |
| Sleep disorders, other/unspecified | FinnGen | 2021 | 216,496 (332) | 16,380,458 | European | Mixed | finn-b-F5_SLEEP_NOS |
| Other sleep disorders | FinnGen | 2021 | 177,660 (1553) | 16,380,337 | European | Mixed | finn-b-G6_SLEEPDISOTH |
| Nonorganic sleeping disorders | FinnGen | 2021 | 218,792 (2214) | 16,380,466 | European | Mixed | finn-b-KRA_PSY_SLEEP_NONORG |
| Sleep disorders (combined) | FinnGen | 2021 | 216,700 (19,155) | 16,380,458 | European | Mixed | finn-b-SLEEP |
| Sleep duration | MRC-IEU | 2018 | 460,099 | 9,851,867 | European | Mixed | ukb-b-4424 |
| Snoring | MRC-IEU | 2018 | 430,438 (270,007) | 9,851,867 | European | Mixed | ukb-b-17400 |
| Nap during day | MRC-IEU | 2018 | 462,400 | 9,851,867 | European | Mixed | ukb-b-4616 |
| Sleeplessness/insomnia | MRC-IEU | 2018 | 462,341 | 9,851,867 | European | Mixed | ukb-b-3957 |
| Diagnoses—main ICD10: G47.3 Sleep apnoea | MRC-IEU | 2018 | 463,010 (2320) | 9,851,867 | European | Mixed | ukb-b-16781 |
| Daytime dozing/sleeping (narcolepsy) | MRC-IEU | 2018 | 460,913 | 9,851,867 | European | Mixed | ukb-b-5776 |
| Diagnoses—secondary ICD10: G47.3 Sleep apnoea | MRC-IEU | 2018 | 463,010 (1385) | 9,851,867 | European | Mixed | ukb-b-7853 |
| Non-cancer illness code, self-reported: sleep apnoea | MRC-IEU | 2018 | 462,933 (1510) | 9,851,867 | European | Mixed | ukb-b-9155 |
| Getting up in the morning | MRC-IEU | 2018 | 461,658 | 9,851,867 | European | Mixed | ukb-b-2772 |
| Morning/evening person (chronotype) | MRC-IEU | 2018 | 413,343 | 9,851,867 | European | Mixed | ukb-b-4956 |
| Sleep duration (over sleepers) | Jones SE | 2016 | 91,306 (10,102) | 16,563,303 | European | Mixed | ebi-a-GCST006685 |
| Sleep duration (under sleepers) | Jones SE | 2016 | 110,188 (28,980) | 16,561,726 | European | Mixed | ebi-a-GCST006686 |
| sleep duration | UK Biobank | 2016 | 128,266 | 16,761,226 | European | Mixed | ieu-a-1088 |
| Chronotype | UK Biobank | 2016 | 128,266 | 17,032,431 | European | Mixed | ieu-a-1087 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yue, M.; Jin, C.; Jiang, X.; Xue, X.; Wu, N.; Li, Z.; Zhang, L. Causal Effects of Gut Microbiota on Sleep-Related Phenotypes: A Two-Sample Mendelian Randomization Study. Clocks & Sleep 2023, 5, 566-580. https://doi.org/10.3390/clockssleep5030037
Yue M, Jin C, Jiang X, Xue X, Wu N, Li Z, Zhang L. Causal Effects of Gut Microbiota on Sleep-Related Phenotypes: A Two-Sample Mendelian Randomization Study. Clocks & Sleep. 2023; 5(3):566-580. https://doi.org/10.3390/clockssleep5030037
Chicago/Turabian StyleYue, Min, Chuandi Jin, Xin Jiang, Xinxin Xue, Nan Wu, Ziyun Li, and Lei Zhang. 2023. "Causal Effects of Gut Microbiota on Sleep-Related Phenotypes: A Two-Sample Mendelian Randomization Study" Clocks & Sleep 5, no. 3: 566-580. https://doi.org/10.3390/clockssleep5030037
APA StyleYue, M., Jin, C., Jiang, X., Xue, X., Wu, N., Li, Z., & Zhang, L. (2023). Causal Effects of Gut Microbiota on Sleep-Related Phenotypes: A Two-Sample Mendelian Randomization Study. Clocks & Sleep, 5(3), 566-580. https://doi.org/10.3390/clockssleep5030037