
Spatial Variability of Topsoil Properties on a Semi-Arid Floodplain

Abstract
1. Introduction
2. Materials and Methods
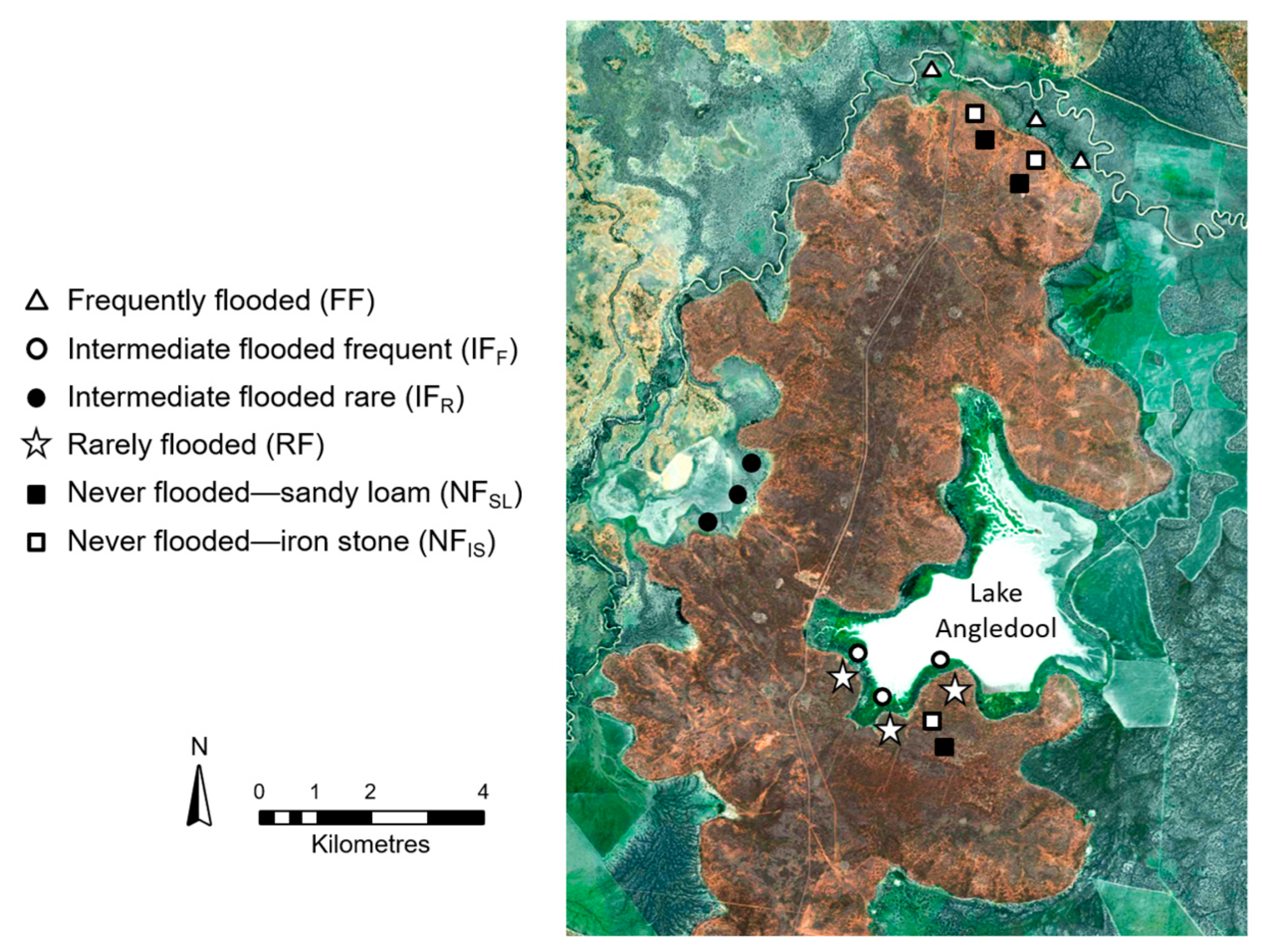
2.1. Study Region
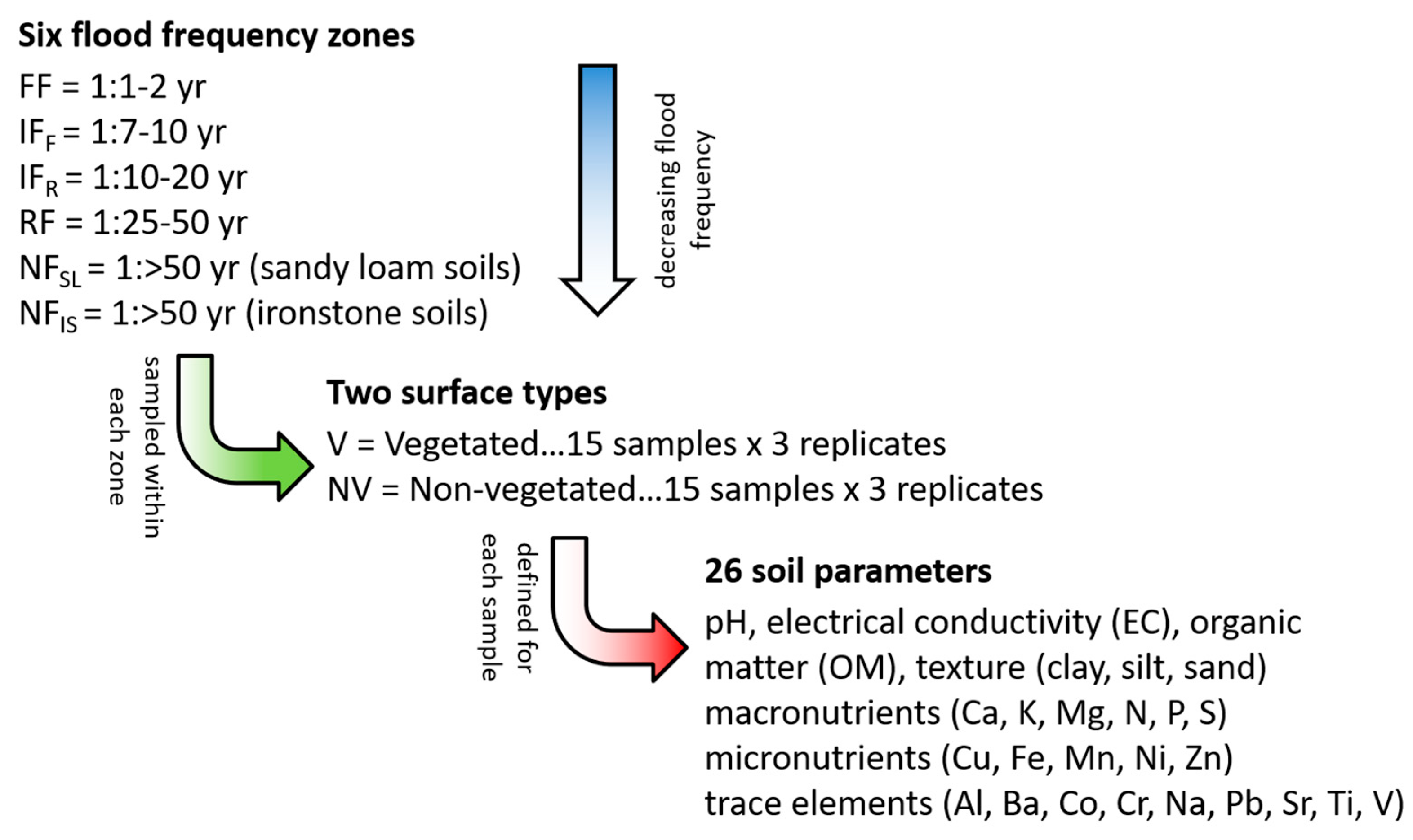
2.2. Field Sampling
2.3. Laboratory Analyses
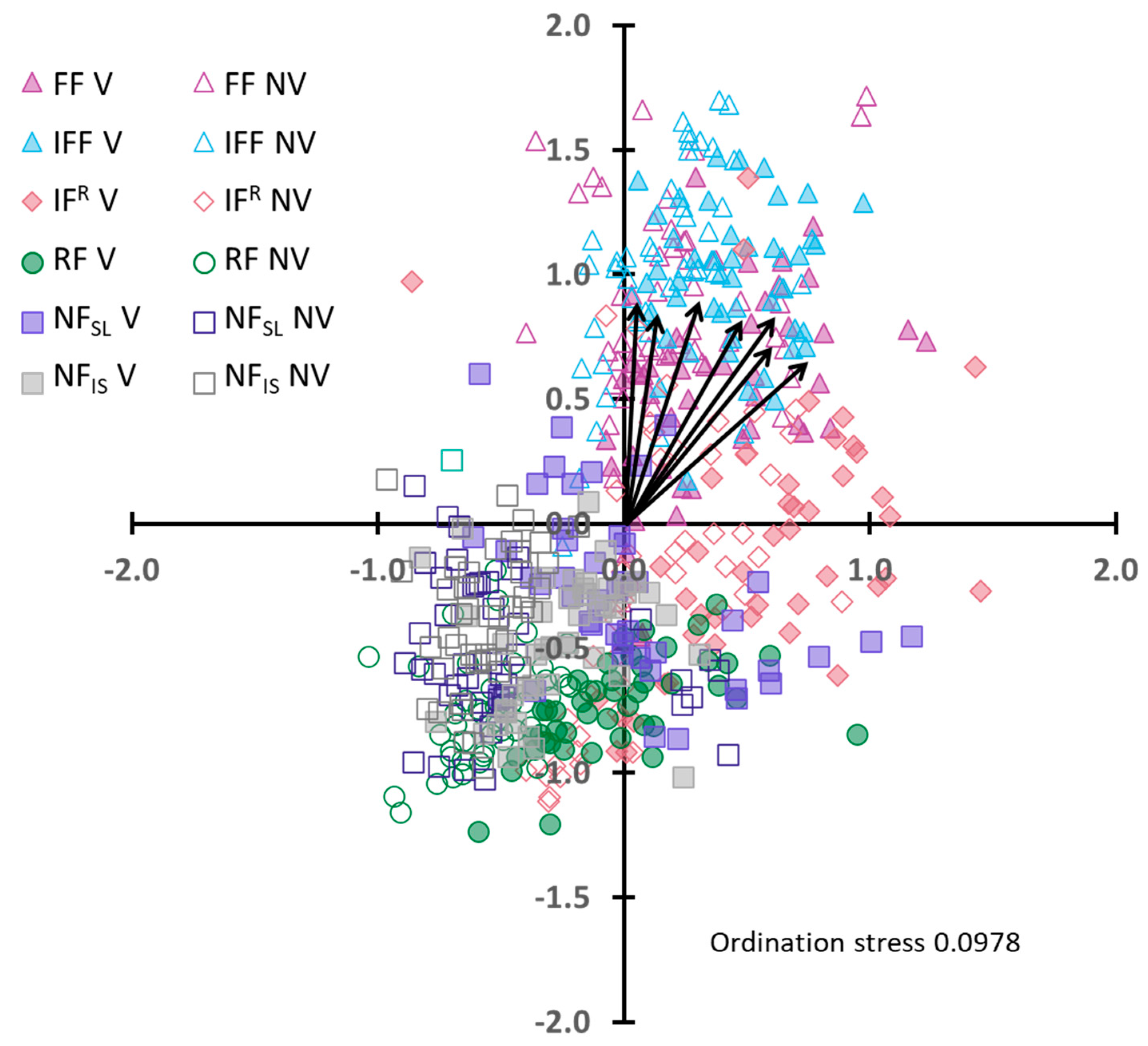
2.4. Statistical Analyses
3. Results
3.1. Whole Site Scale
3.2. Within Flood Frequency Zones
3.3. Between Flood Frequency Zones
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hillman, T.J. Billabongs. In Limnology in Australia; de Deckker, P., Williams, W.D., Eds.; CSIRO Publishing: Melbourne, Australia, 1986; pp. 457–470. [Google Scholar]
- Ogden, R.; Reid, M.; Thoms, M. Soil fertility in a large dryland floodplain: Patterns, processes and the implications of water resource development. Catena 2007, 70, 114–126. [Google Scholar] [CrossRef]
- Mac Nally, R.; Cunningham, S.C.; Baker, P.J.; Horner, G.J.; Thomson, J.R. Dynamics of Murray-Darling floodplain forests under multiple stressors: The past, present, and future of an Australian icon. Water Resour. Res. 2011, 47, W00G05. [Google Scholar] [CrossRef]
- Leauthaud, C.; Kergoat, L.; Hiernaux, P.; Grippa, M.; Musila, W.; Duvail, S.; Albergel, J. Modelling the growth of floodplain grasslands to explore the impact of changing hydrological conditions on vegetation productivity. Ecol. Model. 2018, 387, 220–237. [Google Scholar] [CrossRef]
- Boon, P.I.; Frankenberg, J.; Hillman, T.; Oliver, R.; Shiel, R. Billabongs. In The Murray; Mackay, N., Eastburn, D., Eds.; Murray Darling Basin Commission: Canberra, Australia, 1990; pp. 167–181. [Google Scholar]
- Hillman, T.J.; Shiel, R.J. Macro- and microinvertebrates in Australian billabongs. Int. Ver. Theor. Angew. Limnol. 1991, 24, 1581–1587. [Google Scholar] [CrossRef]
- Kingsford, R.T.; Johnson, W. The impact of water diversions on colonially nesting waterbirds in the Macquarie Marshes in arid Australia. Colon. Waterbirds 1999, 21, 159–170. [Google Scholar] [CrossRef]
- Jansen, A.; Healey, M. Frog communities and wetland condition: Relationships with grazing by domestic livestock along an Australian floodplain river. Biol. Conserv. 2003, 109, 207–219. [Google Scholar] [CrossRef]
- Jolly, I.D.; McEwan, K.L.; Holland, K.L. A review of groundwater–surface water interactions in arid/semi-arid wetlands and the consequences of salinity for wetland ecology. Ecohydrology 2008, 1, 43–58. [Google Scholar] [CrossRef]
- Gell, P.A.; Reid, M.A.; Wilby, R.L. Management pathways for the floodplain wetlands of the southern Murray–Darling Basin: Lessons from history. River Res. Appl. 2019, 2019, 1291–1301. [Google Scholar] [CrossRef]
- Ford, D.J.; Cookson, W.R.; Adams, M.A.; Grierson, P.F. Role of soil drying in nitrogen mineralization and microbial community function in semi-arid grasslands of north-west Australia. Soil Biol. Biochem. 2007, 39, 1557–1569. [Google Scholar] [CrossRef]
- McIntyre, R.E.S.; Adams, M.A.; Grierson, P.F. Nitrogen mineralization potential in rewetted soils from a semi-arid stream landscape, north-west Australia. J. Arid Environ. 2009, 73, 48–54. [Google Scholar] [CrossRef]
- Junk, W.J.; Bayley, P.B.; Sparks, R.E. The flood pulse concept in river-floodplain systems. Can. Spec. Publ. Fish. Aquat. Sci. 1989, 106, 110–127. [Google Scholar]
- Thoms, M.C. Floodplain–river ecosystems: Lateral connections and the implications of human interference. Geomorphology 2003, 56, 335–349. [Google Scholar] [CrossRef]
- Olde Venterink, H.; Vermaat, J.E.; Pronk, M.; Wiegman, F.; van der Lee, G.E.M.; van den Hoorn, M.W.; Higler, L.W.G.; Verhoeven, J.T.A. Importance of sediment deposition and denitrification for nutrient retention in floodplain wetlands. Appl. Veg. Sci. 2006, 9, 163–174. [Google Scholar] [CrossRef]
- Prusty, B.A.K.; Chandra, R.; Azeez, P.A. Macronutrients along the sediment profile in a subtropical monsoonal wetland in India. Wetl. Ecol. Manag. 2010, 18, 91–105. [Google Scholar] [CrossRef]
- Steiger, J.; Gurnell, A.M. Spatial hydrogeomorphological influences on sediment and nutrient deposition in riparian zones: Observations from the Garonne River, France. Geomorphology 2002, 49, 1–23. [Google Scholar] [CrossRef]
- Ward, J.V.; Tockner, K.; Arscott, D.B.; Claret, C. Riverine landscape diversity. Freshw. Biol. 2002, 47, 517–539. [Google Scholar] [CrossRef]
- Reid, M.A.; Ogden, R.; Thoms, M.C. The influence of flood frequency, geomorphic setting and grazing on plant communities and plant biomass on a large dryland floodplain. J. Arid Environ. 2011, 75, 815–826. [Google Scholar] [CrossRef]
- Duarte, C.; Magurran, A.E.; Zuanon, J.; Deus, C.P. Trophic ecology of benthic fish assemblages in a lowland river in the Brazilian Amazon. Aquat. Ecol. 2019, 53, 707–718. [Google Scholar] [CrossRef]
- Sanchez-Carrillo, S.; Alvarez-Cobelas, M.; Angeler, D.G. Sedimentation in the semi-arid freshwater wetland Las Tablas De Daimiel (Spain). Wetlands 2001, 21, 112–124. [Google Scholar] [CrossRef]
- Baldwin, D.S.; Colloff, M.J.; Rees, G.N.; Chariton, A.A.; Watson, G.E.; Court, L.N.; Hartley, D.M.; Morgan, M.J.; King, A.J.; Wilson, J.S.; et al. Impacts of inundation and drought on eukaryote biodiversity in semi-arid floodplain soils. Mol. Ecol. 2013, 22, 1746–1758. [Google Scholar] [CrossRef]
- Higgisson, W.; Higgisson, B.; Powell, M.; Driver, P.; Dyer, F. Impacts of water resource development on hydrological connectivity of different floodplain habitats in a highly variable system. River Res. Appl. 2019, 36, 542–552. [Google Scholar] [CrossRef]
- Angers, D.A.; Caron, J. Plant-induced changes in soil structure: Processes and feedbacks. Biogeochemistry 1998, 42, 55–72. [Google Scholar] [CrossRef]
- Ehrenfeld, J.G.; Ravit, B.; Elgersma, K. Feedback in the plant-soil system. Annu. Rev. Environ. Resour. 2005, 30, 75–115. [Google Scholar] [CrossRef]
- Schlesinger, W.H.; Raikes, J.A.; Hartely, A.E.; Cross, A.F. On the spatial pattern of soil nutrients in desert ecosystems. Ecology 1996, 77, 364–375. [Google Scholar] [CrossRef]
- Schlesinger, W.H.; Pilmanis, A.M. Plant-soil interactions in deserts. Biogeochemistry 1998, 42, 169–187. [Google Scholar] [CrossRef]
- Thompson, D.B.; Walker, L.R.; Landau, F.H.; Stark, L.R. The influence of elevation, shrub species and biological soil crust on fertile islands in the Mojave Desert, USA. J. Arid Environ. 2005, 61, 609–629. [Google Scholar] [CrossRef]
- Butterfield, B.J.; Briggs, J.M. Patch dynamics of soil biotic feedbacks in the Sonoran Desert. J. Arid Environ. 2009, 73, 96–102. [Google Scholar] [CrossRef]
- Segoli, M.; Ungar, E.D.; Shachak, M. Fine-scale spatial heterogeneity of resource modulation in semi-arid “Islands of Fertility”. Arid Land Res. Manag. 2012, 26, 344–354. [Google Scholar] [CrossRef]
- Azcón-Aguilar, C.; Palenzuela, J.; Roldán, A. Analysis of the mycorrhizal potential in the rhizosphere of representative plant species from desertification-threatened Mediterranean shrublands. Appl. Soil Ecol. 2003, 22, 29–37. [Google Scholar] [CrossRef]
- Zaady, E.; Groffman, P.M.; Shachak, M. Litter as a regulator of N and C dynamics in macrophytic patches in Negev desert soils. Soil Biol. Biochem. 1996, 28, 39–46. [Google Scholar] [CrossRef]
- Whitford, W.G. Animal feedbacks in desertification: An overview. Rev. Chil. Hist. Nat. 1993, 66, 243–251. [Google Scholar]
- Ridolfi, L.; Laio, F.; D’Odorico, P. Fertility island formation and evolution in dryland ecosystems. Ecol. Soc. 2008, 13, 5. Available online: http://www.ecologyandsociety.org/vol13/iss1/art5/ (accessed on 3 June 2022). [CrossRef]
- Neave, M.; Abrahams, A.D. Impact of small mammal disturbances on sediment yield from grassland and shrubland ecosystems in the Chihuahuan Desert. Catena 2001, 44, 285–303. [Google Scholar] [CrossRef]
- Neave, M.; Rayburg, S. Nonlinear biofluvial responses to vegetation change in a semiarid environment. Geomorphology 2007, 89, 217–239. [Google Scholar] [CrossRef]
- Cheng, F.-S.; Zeng, D.-H.; He, X.-Y. Small-Scale spatial variability of soil nutrients and vegetation properties in semi-arid Northern China. Pedosphere 2006, 16, 778–787. [Google Scholar] [CrossRef]
- Dean, W.R.J.; Milton, S.J.; Jeltsch, F. Large trees, fertile islands and birds in arid savanna. J. Arid Environ. 1999, 41, 61–78. [Google Scholar] [CrossRef]
- Maestre, F.T.; Reynolds, J.F. Small-scale spatial heterogeneity in the vertical distribution of soil nutrients has limited effects on the growth and development of Prosopis glandulosa seedlings. Plant Ecol. 2006, 183, 65–75. [Google Scholar] [CrossRef]
- Neave, M.; Rayburg, S. A field investigation into the effects of progressive rainfall-induced soil seal and crust development on runoff and erosion rates: The impact of surface cover. Geomorphology 2007, 87, 378–390. [Google Scholar] [CrossRef]
- Ludwig, J.A.; Tongway, D.J. Spatial organisation of landscapes and its function in semi-arid woodlands, Australia. Landsc. Ecol. 1995, 10, 51–63. [Google Scholar] [CrossRef]
- Collins, B.; Wein, G. Soil Heterogeneity effects on canopy structure and composition during early succession. Plant Ecol. 1998, 138, 217–230. [Google Scholar] [CrossRef]
- Kroes, D.E.; Brinson, M.M. Occurrence of riverine wetlands of floodplains along a climatic gradient. Wetlands 2004, 24, 167–177. [Google Scholar] [CrossRef]
- James, C.S.; Thoms, M.C.; Quinn, G.P. Zooplankton dynamics from inundation to drying in a complex ephemeral floodplain-wetland. Aquat. Sci. 2008, 70, 259–271. [Google Scholar] [CrossRef]
- Gergel, S. Assessing cumulative impacts of levees and dams on floodplain ponds: A neutral-terrain model approach. Ecol. Appl. 2002, 12, 1740–1754. [Google Scholar] [CrossRef]
- Kirby, M.; Evans, R.; Walker, G.; Cresswell, R.; Coram, J.; Khan, S.; Paydar, Z.; Mainuddin, M.; McKenzie, N.; Ryan, S. The Shared Water Resources of the Murray-Darling Basin. Part I in a Two Part Series on the Shared Water Resources of the Murray-Darling Basin Prepared for the Murray-Darling Basin Commission; MDBC Publication: Canberra, Australia, 2006. [Google Scholar]
- Kingsford, R.T. Ecological impacts of dams, water diversions and river management on floodplain wetlands in Australia. Austral Ecol. 2000, 25, 109–127. [Google Scholar] [CrossRef]
- Semeniuk, V.; Semeniuk, C.A. A geomorphic approach to global classification for natural inland wetlands and rationalization of the system used by the Ramsar Convention—A discussion. Wetl. Ecol. Manag. 1997, 5, 145–158. [Google Scholar] [CrossRef]
- Castañeda, C.; Herrero, J.; Auxiliadora Casterad, M. Landsat monitoring of laya-lakes in the Spanish monegros desert. J. Arid Environ. 2005, 63, 497–516. [Google Scholar] [CrossRef]
- Minckley, T.A.; Turner, D.S.; Weinstein, S.R. The relevance of wetland conservation in arid regions: A re-examination of vanishing communities in the American Southwest. J. Arid Environ. 2013, 88, 213–221. [Google Scholar] [CrossRef]
- Rayburg, S.; Thoms, M. A coupled hydraulic modeling approach to driving a water balance model for a complex floodplain wetland system. Hydrol. Res. 2009, 40, 364–379. [Google Scholar] [CrossRef]
- Rolls, R.J.; Wilson, G.G. Spatial and temporal patterns in fish assemblages following an artificial extended floodplain inundation event, Northern Murray-Darling Basin, Australia. Environ. Manag. 2010, 45, 822–833. [Google Scholar] [CrossRef]
- Bureau of Meteorology, 2015. Climate Data Online. Viewed 20 January 2019. Available online: https://www.bom.gov.au/climate/data/ (accessed on 20 January 2019).
- Rayburg, S.; Thoms, M.; Lenon, E. Unraveling the physical template of a terminal flood plain-wetland sediment storage system. In Sediment Dynamics and the Hydrology of Fluvial Systems, Proceedings of a Symposium held in Dundee, UK, July 2006; IAHS Publication: Wallingford, UK, 2006; Volume 306, pp. 304–313. [Google Scholar]
- King, D.P. Vegetation Communities of the Northern Floodplains Western New South Wales; Northern Floodplains Regional Planning Committee: Walgett, Australia, 2004. [Google Scholar]
- Condon, R.W. Angledool Station—New Angledool; Western Lands Commission: Sydney, Australia, 1975. [Google Scholar]
- Rayment, G.; Lyons, D. Soil Chemical Methods—Australasia; CSIRO Publishing: Collingwood, Australia, 2010; 495p. [Google Scholar]
- James, C.S.; Capon, S.J.; Quinn, G.P. Nurse plant effects of a dominant shrub (Duma florulenta) on understorey vegetation in a large, semiarid wetland in relation to flood frequency and drying. J. Veg. Sci. 2015, 26, 985–994. [Google Scholar] [CrossRef]
- James, C.S.; Capon, S.J.; White, M.G.; Rayburg, S.C.; Thoms, M.C. Spatial variability of the soil seed bank in a heterogeneous ephemeral wetland system in semi-arid Australia. Plant Ecol. 2007, 190, 205–217. [Google Scholar] [CrossRef]
- Fernando, D.R.; Lynch, J.P.; Reichman, S.M.; Clark, G.J.; Miller, R.E.; Doody, T.M. Inundation of a floodplain lake woodlands system: Nutritional profiling and benefit to mature Eucalyptus largiflorens (Black Box) trees. Wetl. Ecol. Manag. 2018, 26, 961–975. [Google Scholar] [CrossRef]
- Kobayashi, T.; Ralph, T.J.; Sharma, P.; Mitrovic, S.M. Influence of historical inundation frequency on soil microbes (Cyanobacteria, Proteobacteria, Actinobacteria) in semi-arid floodplain wetlands. Mar. Freshw. Res. 2020, 71, 617. [Google Scholar] [CrossRef]
- Kuzmina, Z.V.; Treshkin, S.E.; Shinkarenko, S.S. Effects of river control and climate changes on the dynamics of the terrestrial ecosystems of the Lower Volga region. Arid Ecosyst. 2018, 8, 231–244. [Google Scholar] [CrossRef]
- Capon, S.J. Plant community responses to wetting and drying in a large arid floodplain. River Res. Appl. 2003, 19, 509–520. [Google Scholar] [CrossRef]
- Sandi, S.G.; Saco, P.M.; Saintilan, N.; Wen, L.; Riccardi, G.; Kuczera, G.; Willgoose, G.; Rodríguez, J.F. Detecting inundation thresholds for dryland wetland vulnerability. Adv. Water Resour. 2019, 128, 168–182. [Google Scholar] [CrossRef]
- Thapa, R.; Thoms, M.; Parsons, M. An adaptive cycle hypothesis of semi-arid floodplain vegetation productivity in dry and wet resource states. Ecohydrology 2016, 9, 39–51. [Google Scholar] [CrossRef]
- Argiroff, W.A.; Zak, D.R.; Lanser, C.M.; Wiley, M.J. Microbial community functional potential and composition are shaped by hydrologic connectivity in riverine floodplain soils. Microb. Ecol. 2017, 73, 630–644. [Google Scholar] [CrossRef]
- Wohl, E.; Knox, R.L. A first-order approximation of floodplain soil organic carbon stocks in a river network: The South Platte River, Colorado, USA as a case study. Sci. Total Environ. 2022, 852, 158507. [Google Scholar] [CrossRef]
- Morra, B.; Brisbin, H.; Stringham, T.; Sullivan, B.W. Ecosystem carbon and nitrogen gains following 27 years of grazing management in a semiarid alluvial valley. J. Environ. Manag. 2023, 337, 117724. [Google Scholar] [CrossRef]
- Barbosa-Briones, E.; Cardona-Benavides, A.; Reyes-Hernández, H.; Muñoz-Robles, C. Ecohydrological function of vegetation patches in semi-arid shrublands of central Mexico. J. Arid Environ. 2019, 168, 36–45. [Google Scholar] [CrossRef]
- Davids, L.; Pryke, J.S.; Seymour, C.L. High species turnover of arthropod communities associated with distinct vegetation patches (“heuweltjies”) in an arid shrubland. J. Arid Environ. 2023, 213, 104967. [Google Scholar] [CrossRef]
- D’Odorico, P.; Okin, G.S.; Bestelmeyer, B.T. A synthetic review of feedbacks and drivers of shrub encroachment in arid grasslands. Ecohydrology 2012, 5, 520–530. [Google Scholar] [CrossRef]
- Stewart, J.; Parsons, A.J.; Wainwright, J.; Okin, G.S.; Bestelmeyer, B.T.; Fredrickson, L.; Schlesinger, W.H. Modeling emergent patterns of dynamic desert ecosystems. Ecol. Monogr. 2014, 84, 373–410. [Google Scholar] [CrossRef]
- Morris, L.R.; Monaco, T.A.; Blank, R.; Sheley, R.L. Long-term redevelopment of resource islands in shrublands of the great basin, USA. Ecosphere 2013, 4, 12. [Google Scholar] [CrossRef]
- Harman, C.J.; Lohse, K.A.; Troch, P.A.; Sivapalan, M. Spatial patterns of vegetation, soils, and microtopography from terrestrial laser scanning on two semiarid hillslopes of contrasting lithology. J. Geophys. Res. Biogeosci. 2014, 119, 163–180. [Google Scholar] [CrossRef]
- Boyd, D.S.; Davies, K.W. Differential seedling performance and environmental correlates in shrub canopy vs. interspace microsites. J. Arid Environ. 2012, 87, 50–57. [Google Scholar] [CrossRef]
- Chimal-Sánchez, E.; Montaño, N.O.; Camargo-Ricalde, S.L.; García-Sánchez, R.; Hernández Cuevas, L.V. Endemic Mimosa species, forming resource islands or not, and rainfall seasonality jointly influence arbuscular mycorrhizal fungi communities in a semiarid ecosystem of Mexico. Trees 2022, 36, 609–625. [Google Scholar] [CrossRef]
- Vogel, B.; Rostagno, C.M.; Molina, L.; Antilef, M.; La Manna, L. Cushion shrubs encroach subhumid rangelands and form fertility islands along a grazing gradient in Patagonia. Plant Soil 2022, 475, 623–643. [Google Scholar] [CrossRef]
- Minnick, T.J.; Alward, R.D. Plant-soil feedbacks and the partial recovery of soil spatial patterns on abandoned well pads in a sagebrush shrubland. Ecol. Appl. 2015, 25, 3–10. [Google Scholar] [CrossRef]
- García-Sánchez, R.; Camargo-Ricalde, S.L.; García-Moya, E.; Luna-Cavazos, M.; Romero-Manzanares, A.; Manuel Montaño, N. Prosopis laevigata and Mimosa biuncifera (Leguminosae), jointly influence plant diversity and soil fertility of a Mexican semiarid ecosystem. Rev. Biol. Trop./Int. J. Trop. Biol. Conserv. 2012, 60, 87–103. [Google Scholar] [CrossRef]
- Aguilera, L.E.; Gutiérrez, J.R.; Meserve, P.L. Variation in soil micro-organisms and nutrients underneath and outside the canopy of Adesmia bedwellii (Papilionaceae) shrubs in arid coastal Chile following drought and above average rainfall. J. Arid. Environ. 1999, 42, 61–70. [Google Scholar] [CrossRef]
- Hortal, S.; Bastida, F.; Armas, C.; Lozano, Y.M.; Moreno, J.L.; García, C.; Pugnaire, F.I. Soil microbial community under a nurse-plant species changes in composition, biomass and activity as the nurse grows. Soil Biol. Biochem. 2013, 64, 139–146. [Google Scholar] [CrossRef]
- Ludwig, J.A.; Wilcox, B.P.; Breshears, D.D.; Tongway, D.J.; Imeson, A.C. Vegetation patches and runoff-erosion as interacting ecohydrological processes in semiarid landscapes. Ecology 2005, 86, 288–297. [Google Scholar] [CrossRef]
- Maestre, F.T.; Cortina, J. Spatial patterns of surface soil properties and vegetation in a Mediterranean semi-arid steppe. Plant Soil 2002, 241, 279–291. [Google Scholar] [CrossRef]
- Yao, Y.; Shao, M.; Fu, X.; Wang, X.; Wei, X. Effects of shrubs on soil nutrients and enzymatic activities over a 0–100 cm soil profile in the desert-loess transition zone. Catena 2019, 174, 362–370. [Google Scholar] [CrossRef]
- Hanan, E.J.; Ross, M.S. Across scale patterning of plant-soil-water interactions surrounding tree islands in Southern Everglades landscapes. Landsc. Ecol. 2010, 25, 463–476. [Google Scholar] [CrossRef]
- Maestre, F.T.; Eldridge, D.J.; Soliveres, S.; Kefi, S.; Delgado-Baquerizo, M.; Bowker, M.A.; Garcıa-Palacios, P.; Gaitan, J.; Gallardo, A.; Lazaro, R.; et al. Structure and functioning of dryland ecosystems in a changing world. Annu. Rev. Ecol. Evol. Syst. 2016, 47, 215–237. [Google Scholar] [CrossRef]
- Baartman, J.E.M.; Temme, A.J.A.M.; Saco, P.M. The effect of landform variation on vegetation patterning and related sediment dynamics. Earth Surf. Process. Landf. 2018, 43, 2121–2135. [Google Scholar] [CrossRef]
- Li, W.; Li, X.; Huang, Y.; Wang, P.; Zhang, C. Spatial patch structure and adaptive strategy for desert shrub of Reaumuria soongorica in arid ecosystem of the Heihe River Basin. J. Geogr. Sci. 2019, 29, 1507–1526. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetated (V) | Non-Vegetated (NV) | V vs. NV | |||
|---|---|---|---|---|---|
| Parameter | Mean | CV | Mean | CV | p |
| pH | 6.54 | 0.06 | 5.96 | 0.11 | <0.0001 |
| EC (µS/cm) | 170.4 | 0.66 | 63.0 | 0.68 | <0.0001 |
| OM (%) | 1.83 | 0.78 | 0.71 | 0.91 | <0.0001 |
| Clay (%) | 16.2 | 0.54 | 18.7 | 0.46 | <0.0001 |
| Silt (%) | 78.0 | 0.11 | 75.0 | 0.10 | <0.0001 |
| Sand (%) | 5.73 | 0.77 | 6.28 | 0.77 | 0.247 |
| Macronutrients | |||||
| Ca | 2974 | 0.88 | 1639 | 0.80 | <0.0001 |
| K | 2870 | 0.71 | 2561 | 0.71 | 0.033 |
| Mg | 2204 | 0.71 | 2099 | 0.80 | 0.027 |
| N | 1451.4 | 0.64 | 561.9 | 1.01 | <0.0001 |
| P | 254.4 | 0.32 | 202.4 | 0.33 | <0.0001 |
| S | 139.6 | 0.61 | 55.9 | 0.69 | <0.0001 |
| Micronutrients | |||||
| Cu | 9.31 | 0.34 | 9.47 | 0.41 | 0.835 |
| Fe | 16,960 | 0.48 | 18,045 | 0.51 | 0.328 |
| Mn | 390.5 | 0.51 | 376.9 | 0.63 | 0.153 |
| Ni | 12.0 | 0.51 | 12.3 | 0.60 | 0.925 |
| Zn | 25.7 | 0.59 | 25.0 | 0.65 | 0.170 |
| Other | |||||
| Al | 21,398 | 0.52 | 22,188 | 0.57 | 0.979 |
| Ba | 1088.9 | 3.96 | 894.2 | 3.93 | 0.174 |
| Co | 7.99 | 0.91 | 7.74 | 0.55 | 0.835 |
| Cr | 21.1 | 0.37 | 22.9 | 0.43 | 0.106 |
| Na | 127.5 | 1.15 | 146.0 | 1.17 | 0.541 |
| Pb | 6.06 | 0.45 | 6.36 | 0.48 | 0.243 |
| Sr | 34.6 | 0.78 | 22.9 | 0.74 | <0.0001 |
| Ti | 201.7 | 0.47 | 233.9 | 0.45 | <0.0001 |
| V | 33.6 | 0.47 | 37.1 | 0.52 | 0.083 |
| FF | IFF | IFR | RF | NFSL | NFIS | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Parameter | Mean | CV | Mean | CV | Mean | CV | Mean | CV | Mean | CV | Mean | CV |
| pH | 6.42 | 0.02 | 6.87 | 0.02 | 6.58 | 0.03 | 6.52 | 0.06 | 6.65 | 0.08 | 6.21 | 0.08 |
| EC (µS/cm) | 117.8 | 0.37 | 167.4 | 0.36 | 231.1 | 0.37 | 103.6 | 0.56 | 290.7 | 0.52 | 111.8 | 0.83 |
| OM% | 1.48 | 0.52 | 1.32 | 0.50 | 3.59 | 0.63 | 1.70 | 0.57 | 1.89 | 0.49 | 1.05 | 0.48 |
| Clay% | 14.0 | 0.17 | 18.2 | 0.49 | 11.3 | 0.39 | 17.0 | 0.68 | 13.5 | 0.47 | 23.5 | 0.41 |
| Silt% | 73.9 | 0.04 | 74.4 | 0.10 | 83.8 | 0.06 | 79.7 | 0.14 | 81.8 | 0.07 | 74.7 | 0.12 |
| Sand% | 12.15 | 0.29 | 7.48 | 0.32 | 4.97 | 0.33 | 3.25 | 0.42 | 4.76 | 1.00 | 1.79 | 1.01 |
| Macronutrients | ||||||||||||
| Ca | 2877.8 | 0.26 | 3343.4 | 0.18 | 5301.5 | 0.82 | 2363.7 | 0.95 | 2759.9 | 0.93 | 1196.4 | 0.56 |
| K | 3010.3 | 0.24 | 6097.9 | 0.16 | 4200.9 | 0.42 | 1166.7 | 0.63 | 1346.1 | 0.26 | 1396.0 | 0.23 |
| Mg | 3489.5 | 0.20 | 4285.6 | 0.18 | 2834.1 | 0.46 | 1052.2 | 0.37 | 893.3 | 0.27 | 671.8 | 0.30 |
| N | 1006.5 | 0.58 | 1311.2 | 0.43 | 2637.1 | 0.48 | 1176.3 | 0.52 | 1620.3 | 0.38 | 957.1 | 0.50 |
| P | 230.8 | 0.36 | 195.4 | 0.19 | 274.3 | 0.37 | 287.5 | 0.33 | 283.6 | 0.23 | 254.9 | 0.21 |
| S | 106.1 | 0.44 | 123.1 | 0.31 | 242.8 | 0.52 | 120.5 | 0.49 | 150.1 | 0.44 | 95.0 | 0.42 |
| Micronutrients | ||||||||||||
| Cu | 10.70 | 0.20 | 13.41 | 0.13 | 10.23 | 0.28 | 5.85 | 0.22 | 8.12 | 0.25 | 7.58 | 0.19 |
| Fe | 25,297.9 | 0.21 | 26,911.2 | 0.15 | 15,234.2 | 0.37 | 9664.2 | 0.50 | 13,637.3 | 0.33 | 11,015.3 | 0.22 |
| Mn | 530.2 | 0.27 | 609.1 | 0.15 | 175.8 | 0.41 | 215.3 | 0.64 | 357.9 | 0.42 | 454.8 | 0.25 |
| Ni | 18.15 | 0.19 | 19.43 | 0.13 | 12.11 | 0.38 | 5.80 | 0.46 | 9.05 | 0.27 | 7.36 | 0.38 |
| Zn | 41.3 | 0.21 | 44.8 | 0.20 | 27.3 | 0.37 | 11.6 | 0.46 | 15.0 | 0.26 | 14.3 | 0.25 |
| Other | ||||||||||||
| Al | 33,346.8 | 0.21 | 36,325.5 | 0.14 | 18,759.3 | 0.38 | 11,237.7 | 0.19 | 16,051.7 | 0.27 | 12,667.8 | 0.20 |
| Ba | 106.2 | 0.26 | 134.1 | 0.15 | 87.9 | 0.31 | 44.1 | 0.31 | 6121.4 | 1.47 | 39.6 | 0.40 |
| Co | 11.02 | 0.20 | 13.36 | 0.12 | 5.36 | 0.43 | 6.31 | 2.47 | 6.14 | 0.30 | 5.75 | 0.21 |
| Cr | 26.9 | 0.26 | 28.5 | 0.13 | 17.7 | 0.32 | 14.8 | 0.39 | 20.7 | 0.40 | 18.3 | 0.29 |
| Na | 146.7 | 0.22 | 387.6 | 0.42 | 74.8 | 0.30 | 35.3 | 0.49 | 34.4 | 0.37 | 86.2 | 1.21 |
| Pb | 7.80 | 0.30 | 9.75 | 0.21 | 5.54 | 0.30 | 3.19 | 0.49 | 5.31 | 0.26 | 4.78 | 0.18 |
| Sr | 36.9 | 0.21 | 45.3 | 0.14 | 60.9 | 0.76 | 27.9 | 0.78 | 22.4 | 0.61 | 14.2 | 0.52 |
| Ti | 272.9 | 0.29 | 307.2 | 0.29 | 115.2 | 0.36 | 125.3 | 0.29 | 227.0 | 0.29 | 162.5 | 0.24 |
| V | 48.2 | 0.31 | 45.2 | 0.17 | 30.3 | 0.39 | 20.6 | 0.85 | 32.0 | 0.37 | 25.2 | 0.23 |
| FF | IFF | IFR | RF | NFSL | NFIS | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Parameter | Mean | CV | Mean | CV | Mean | CV | Mean | CV | Mean | CV | Mean | CV |
| pH | 6.47 | 0.04 | 6.84 | 0.02 | 6.15 | 0.03 | 5.36 | 0.06 | 5.50 | 0.09 | 5.42 | 0.05 |
| EC (µS/cm) | 79.3 | 0.24 | 83.6 | 0.27 | 99.9 | 0.44 | 30.3 | 0.62 | 68.2 | 0.73 | 16.5 | 0.35 |
| OM % | 0.73 | 0.51 | 0.52 | 0.53 | 1.30 | 0.98 | 0.59 | 0.44 | 0.67 | 0.54 | 0.46 | 0.35 |
| Clay % | 13.6 | 0.23 | 16.7 | 0.29 | 18.3 | 0.26 | 23.0 | 0.61 | 17.9 | 0.45 | 22.6 | 0.34 |
| Silt % | 72.9 | 0.05 | 74.0 | 0.07 | 76.7 | 0.06 | 73.1 | 0.17 | 78.6 | 0.08 | 75.0 | 0.10 |
| Sand % | 13.51 | 0.31 | 9.39 | 0.29 | 4.99 | 0.43 | 3.91 | 0.70 | 3.45 | 0.88 | 2.45 | 0.63 |
| Macronutrients | ||||||||||||
| Ca | 3191.6 | 0.23 | 3078.4 | 0.23 | 1860.7 | 0.54 | 483.1 | 0.48 | 587.8 | 0.49 | 636.8 | 0.51 |
| K | 3138.0 | 0.18 | 4896.8 | 0.20 | 3948.4 | 0.51 | 1051.0 | 0.62 | 1082.4 | 0.40 | 1249.4 | 0.39 |
| Mg | 3952.7 | 0.16 | 4171.9 | 0.20 | 2435.2 | 0.56 | 691.3 | 0.30 | 657.0 | 0.42 | 685.4 | 0.36 |
| N | 496.4 | 0.76 | 510.9 | 0.48 | 1161.9 | 0.87 | 344.2 | 0.79 | 613.0 | 0.52 | 245.2 | 0.69 |
| P | 203.9 | 0.37 | 147.9 | 0.23 | 192.9 | 0.37 | 239.0 | 0.23 | 200.4 | 0.28 | 230.1 | 0.23 |
| S | 71.1 | 0.34 | 66.7 | 0.28 | 95.2 | 0.66 | 36.8 | 0.48 | 40.6 | 0.40 | 25.1 | 0.33 |
| Micronutrients | ||||||||||||
| Cu | 12.33 | 0.18 | 13.54 | 0.16 | 10.55 | 0.42 | 5.48 | 0.27 | 6.79 | 0.30 | 8.10 | 0.26 |
| Fe | 29,524.8 | 0.20 | 27,225.8 | 0.17 | 15,672.6 | 0.47 | 9550.2 | 0.24 | 12,662.4 | 0.37 | 13,633.8 | 0.32 |
| Mn | 649.1 | 0.39 | 571.0 | 0.17 | 185.2 | 0.51 | 148.7 | 0.36 | 267.5 | 0.35 | 439.7 | 0.36 |
| Ni | 21.09 | 0.17 | 19.90 | 0.17 | 11.54 | 0.56 | 5.09 | 0.40 | 6.59 | 0.56 | 9.49 | 0.36 |
| Zn | 44.1 | 0.15 | 43.7 | 0.22 | 25.1 | 0.45 | 11.8 | 0.92 | 12.0 | 0.25 | 13.4 | 0.31 |
| Other | ||||||||||||
| Al | 38,277.5 | 0.16 | 36,836.1 | 0.16 | 19,277.8 | 0.49 | 11,081.4 | 0.17 | 13,435.2 | 0.29 | 14,221.3 | 0.30 |
| Ba | 129.2 | 0.36 | 138.2 | 0.21 | 74.7 | 0.32 | 36.4 | 0.31 | 4942.0 | 1.49 | 44.6 | 0.42 |
| Co | 12.31 | 0.19 | 13.20 | 0.15 | 5.62 | 0.52 | 3.46 | 0.38 | 5.28 | 0.31 | 6.60 | 0.36 |
| Cr | 32.2 | 0.31 | 29.3 | 0.15 | 17.8 | 0.40 | 15.2 | 0.27 | 20.8 | 0.47 | 22.4 | 0.42 |
| Na | 226.5 | 0.23 | 444.8 | 0.33 | 104.9 | 1.06 | 34.0 | 0.70 | 26.9 | 0.48 | 38.8 | 0.43 |
| Pb | 9.03 | 0.21 | 9.90 | 0.27 | 5.26 | 0.49 | 3.54 | 0.26 | 4.76 | 0.41 | 5.66 | 0.27 |
| Sr | 42.88 | 0.22 | 43.27 | 0.18 | 23.82 | 0.47 | 7.97 | 0.31 | 8.49 | 0.46 | 10.71 | 0.39 |
| Ti | 310.4 | 0.28 | 349.4 | 0.36 | 164.4 | 0.39 | 172.9 | 0.20 | 198.9 | 0.35 | 207.4 | 0.28 |
| V | 59.4 | 0.43 | 45.6 | 0.22 | 32.9 | 0.50 | 21.6 | 0.37 | 30.1 | 0.39 | 32.7 | 0.36 |
| Parameter | FF V v FF NV | IFF V v IFF NV | IFR V v IFR NV | RF V vs. RF NV | NFSL V v NFSL NV | NFIS V v NFIS NV |
|---|---|---|---|---|---|---|
| pH | 0.771 | 0.045 (V) | <0.0001 (V) | <0.0001 (V) | <0.0001 (V) | <0.0001 (V) |
| EC (µS/cm) | <0.0001 (V) | <0.0001 (V) | <0.0001 (V) | <0.0001 (V) | <0.0001 (V) | <0.0001 (V) |
| OM% | <0.0001 (V) | <0.0001 (V) | <0.0001 (V) | <0.0001 (V) | <0.0001 (V) | <0.0001 (V) |
| Clay% | 0.647 | 0.653 | <0.0001 (NV) | <0.0001 (NV) | 0.001 (NV) | 0.822 |
| Silt% | 0.253 | 0.267 | <0.0001 (V) | <0.0001 (V) | 0.002 (V) | 0.742 |
| Sand% | 0.138 | 0.002 (NV) | 0.930 | 0.602 | 0.243 | 0.002 (NV) |
| Macronutrients | ||||||
| Ca | 0.042 (NV) | 0.086 | <0.0001 (V) | <0.0001 (V) | <0.0001 (V) | <0.0001 (V) |
| K | 0.333 | <0.0001 (V) | 0.519 | 0.313 | <0.0001 (V) | 0.024 (V) |
| Mg | 0.002 (NV) | 0.458 | 0.149 | <0.0001 (V) | <0.0001 (V) | 0.968 |
| N | <0.0001 (V) | <0.0001 (V) | <0.0001 (V) | <0.0001 (V) | <0.0001 (V) | <0.0001 (V) |
| P | 0.087 | <0.0001 (V) | <0.0001 (V) | 0.012 (V) | <0.0001 (V) | 0.024 (V) |
| S | <0.0001 (V) | <0.0001 (V) | <0.0001 (V) | <0.0001 (V) | <0.0001 (V) | <0.0001 (V) |
| Micronutrients | ||||||
| Cu | 0.001 (NV) | 0.600 | 0.904 | 0.226 | 0.001 (V) | 0.420 |
| Fe | 0.002 (NV) | 0.410 | 0.994 | 0.349 | 0.269 | 0.006 (NV) |
| Mn | 0.028 (NV) | 0.087 | 0.628 | 0.016 (V) | 0.005 (V) | 0.420 |
| Ni | <0.0001 (NV) | 0.358 | 0.784 | 0.315 | 0.001 (V) | 0.044 (NV) |
| Zn | 0.117 | 0.717 | 0.341 | 0.594 | <0.0001 (V) | 0.262 |
| Other | ||||||
| Al | 0.002 (NV) | 0.503 | 0.878 | 0.865 | 0.005 (V) | 0.165 |
| Ba | 0.026 (NV) | 0.550 | 0.028 (V) | 0.002 (V) | 0.001 (V) | 0.153 |
| Co | 0.013 (NV) | 0.704 | 0.704 | 0.146 | 0.020 (V) | 0.156 |
| Cr | 0.007 (NV) | 0.149 | 0.675 | 0.223 | 0.415 | 0.017 (NV) |
| Na | <0.0001 (NV) | 0.018 (NV) | 0.556 | 0.040 (V) | <0.0001 (V) | 0.910 |
| Pb | 0.011 (NV) | 0.156 | 0.594 | 0.109 | 0.366 | 0.003 (NV) |
| Sr | 0.007 (NV) | 0.259 | <0.0001 (V) | <0.0001 (V) | <0.0001 (V) | 0.040 (V) |
| Ti | 0.036 (NV) | 0.217 | <0.0001 (NV) | <0.0001 (NV) | 0.084 | <0.0001 (NV) |
| V | 0.044 (NV) | 0.929 | 0.710 | 0.006 (NV) | 0.370 | 0.001 (NV) |
| FF V | FF NV | IFF V | IFF NV | IFR V | IFR NV | RF V | RF NV | NFSL V | NFSL NV | NFIS V | NFIS NV | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FF V | ||||||||||||
| FF NV | 0.120 | |||||||||||
| IFF V | 0.454 | 0.419 | ||||||||||
| IFF NV | 0.443 | 0.270 | 0.177 | |||||||||
| IFR V | 0.631 | 0.800 | 0.751 | 0.824 | ||||||||
| IFR NV | 0.435 | 0.589 | 0.624 | 0.624 | 0.233 | |||||||
| RF V | 0.905 | 0.967 | 0.988 | 0.978 | 0.454 | 0.345 | ||||||
| RF NV | 0.950 | 0.976 | 0.998 | 0.988 | 0.741 | 0.441 | 0.368 | |||||
| NFSL V | 0.825 | 0.939 | 0.957 | 0.956 | 0.431 | 0.409 | 0.314 | 0.640 | ||||
| NFSL NV | 0.895 | 0.954 | 0.993 | 0.972 | 0.704 | 0.363 | 0.402 | 0.188 | 0.408 | |||
| NFIS V | 0.888 | 0.954 | 0.987 | 0.968 | 0.612 | 0.386 | 0.341 | 0.457 | 0.316 | 0.296 | ||
| NFIS NV | 0.883 | 0.933 | 0.991 | 0.960 | 0.753 | 0.449 | 0.607 | 0.304 | 0.573 | 0.144 | 0.341 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carroll-MacDonald, T.-A.; Rayburg, S.; Neave, M. Spatial Variability of Topsoil Properties on a Semi-Arid Floodplain. Soil Syst. 2023, 7, 42. https://doi.org/10.3390/soilsystems7020042
Carroll-MacDonald T-A, Rayburg S, Neave M. Spatial Variability of Topsoil Properties on a Semi-Arid Floodplain. Soil Systems. 2023; 7(2):42. https://doi.org/10.3390/soilsystems7020042
Chicago/Turabian StyleCarroll-MacDonald, Tiffany-Anne, Scott Rayburg, and Melissa Neave. 2023. "Spatial Variability of Topsoil Properties on a Semi-Arid Floodplain" Soil Systems 7, no. 2: 42. https://doi.org/10.3390/soilsystems7020042
APA StyleCarroll-MacDonald, T.-A., Rayburg, S., & Neave, M. (2023). Spatial Variability of Topsoil Properties on a Semi-Arid Floodplain. Soil Systems, 7(2), 42. https://doi.org/10.3390/soilsystems7020042