Water Table Dynamics Control Carbon Losses from the Destabilization of Soil Organic Matter in a Small, Lowland Agricultural Catchment

 ,
,
Abstract
1. Introduction
2. Material and Methods
2.1. Description of the Study Area and Sampling Sites
2.2. Analytical Methods
2.2.1. Soil CO2 Efflux Measurements
2.2.2. Physical Parameters
2.2.3. Soil Solution Measurements
2.3. Modeling the Control of Abiotic Parameters on the DIC/DOC Concentrations and FCO2
2.4. Modeling the Coupling between DIC, DOC and FCO2
3. Results
3.1. Temporal Dynamics of the Hydrological Variables and Soil Temperature
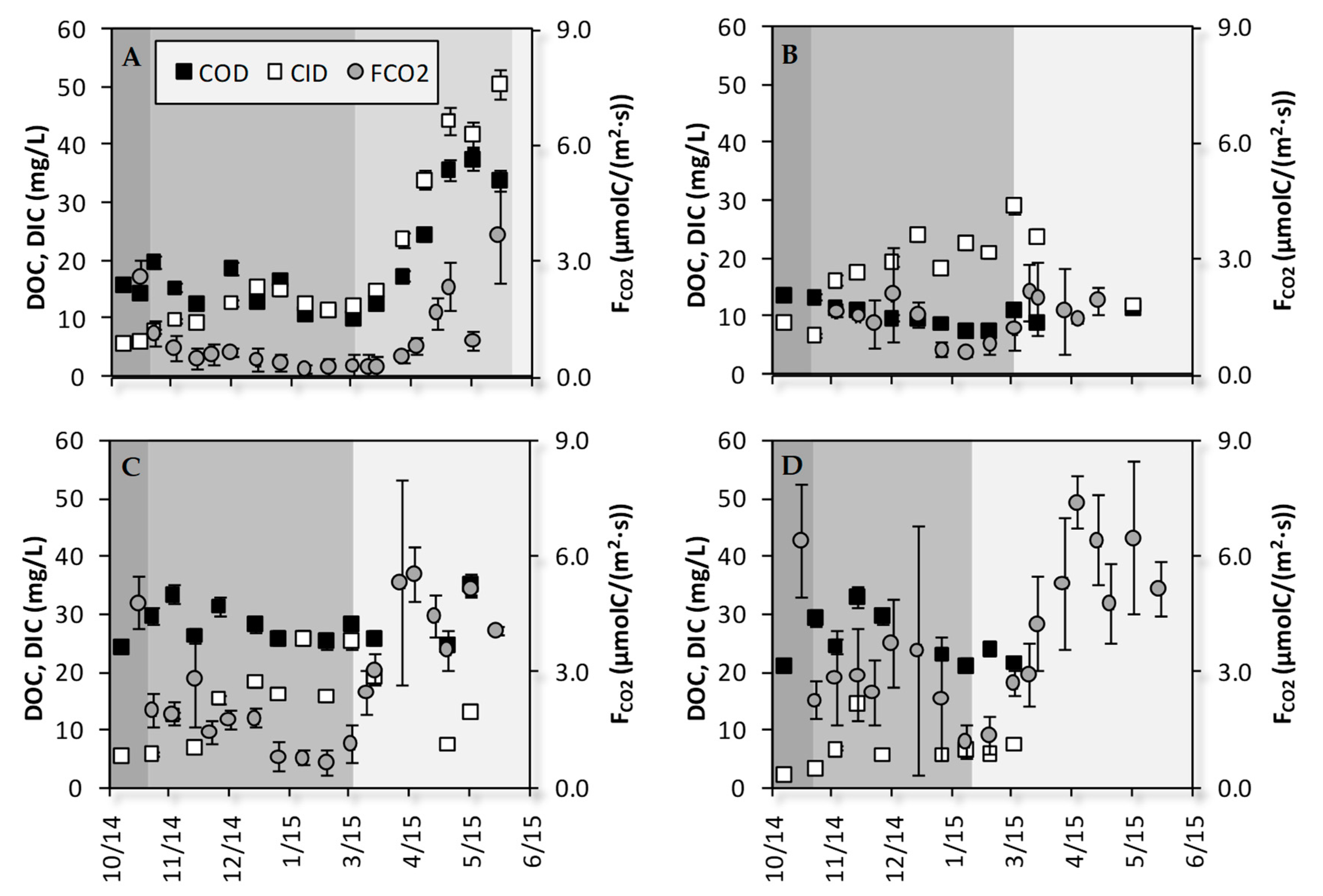
3.2. Temporal Dynamics of DIC, DOC Concentrations and FCO2
3.3. Correlations between DIC, DOC, FCO2, WFPS and Soil Temperature
4. Discussion
4.1. Does the DOC Concentration Control the Soil CO2 Efflux?
4.2. How Do the Water Table Dynamics Drive the Coupling between DIC, DOC and FCO2?
4.3. Even if FCO2 Decreases, OM Mineralization Can Continue
4.4. Reducing Periods: A Time for Soil Heterotrophic Respiration?
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Batjes, N.H. Total carbon and nitrogen in the soils of the world. Eur. J. Soil Sci. 1996, 47, 151–163. [Google Scholar] [CrossRef]
- Lehmann, J.; Kleber, M. The contentious nature of soil organic matter. Nature 2015, 528, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.W.I.; Torn, M.S.; Abiven, S.; Dittmar, T.; Guggenberger, G.; Janssens, I.A.; Kleber, M.; Kogel-Knabner, I.; Lehmann, J.; Manning, D.A.C.; et al. Persistence of soil organic matter as an ecosystem property. Nature 2011, 478, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Sollins, P.; Homann, P.; Caldwell, B.A. Stabilization and destabilization of soil organic matter: Mechanisms and controls. Geoderma 1996, 74, 65–105. [Google Scholar] [CrossRef]
- Mayes, M.A.; Lajtha, K.; Bailey, V. Advancing soil carbon cycle science. EOS 2016, 97. [Google Scholar] [CrossRef]
- Kindler, R.; Siemens, J.; Kaiser, K.; Walmsley, D.; Bernhofer, C.; Buchmann, N.; Cellier, P.; Eugster, W.; Gleixner, G.; Grünwald, T.; et al. Dissolved carbon leaching from soil is a crucial component of the net ecosystem carbon balance. Glob. Chang. Biol. 2011, 17, 1167–1185. [Google Scholar] [CrossRef]
- Raich, J.W.; Schlesinger, W.H. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate. Tellus B Chem. Phys. Meteorol. 1992, 44, 81–99. [Google Scholar] [CrossRef]
- Thompson, J.A.; Kolka, R.K. Soil carbon storage estimation in a forested watershed using quantitative soil-landscape modeling. Soil Sci. Soc. Am. J. 2005, 69, 1086–1093. [Google Scholar] [CrossRef]
- Wang, X.; Cammeraat, E.L.H.; Romeijn, P.; Kalbitz, K. Soil organic carbon redistribution by water erosion—The role of CO2 emissions for the carbon budget. PLoS ONE 2014, 9, e96299. [Google Scholar] [CrossRef]
- Brewer, R. Fabric and Mineral Analysis of Soils; John Wiley & Sons: New York, NY, USA, 1964. [Google Scholar]
- Fahey, T.J.; Siccama, T.G.; Driscoll, C.T.; Likens, G.E.; Campbell, J.; Johnson, C.E.; Battles, J.J.; Aber, J.D.; Cole, J.J.; Fisk, M.C.; et al. The biogeochemistry of carbon at hubbard brook. Biogeochemistry 2005, 75, 109–176. [Google Scholar] [CrossRef]
- Neff, J.C.; Asner, G.P. Dissolved organic carbon in terrestrial ecosystems: Synthesis and a model. Ecosystems 2001, 4, 29–48. [Google Scholar] [CrossRef]
- Humbert, G.; Jaffrezic, A.; Fovet, O.; Gruau, G.; Durand, P. Dry-season length and runoff control annual variability in stream DOC dynamics in a small, shallow-groundwater-dominated agricultural watershed. Water Resour. Res. 2015, 51, 7860–7877. [Google Scholar] [CrossRef]
- Denis, M.; Jeanneau, L.; Petitjean, P.; Murzeau, A.; Liotaud, M.; Yonnet, L.; Gruau, G. New molecular evidence for surface and sub-surface soil erosion controls on the composition of stream DOM during storm events. Biogeosciences 2017, 14, 5039–5051. [Google Scholar] [CrossRef]
- Dungait, J.A.J.; Hopkins, D.W.; Gregory, A.S.; Whitmore, A.P. Soil organic matter turnover is governed by accessibility not recalcitrance. Glob. Chang. Biol. 2012, 18, 1781–1796. [Google Scholar] [CrossRef]
- Luo, Y.; Zhou, X. (Eds.) Soil Respiration and the Environment; Academic Press: London, UK, 2006; ISBN 978-0-12-088782-8. [Google Scholar]
- Yan, Z.; Bond-Lamberty, B.; Todd-Brown, K.E.; Bailey, V.L.; Li, S.; Liu, C.; Liu, C. A moisture function of soil heterotrophic respiration that incorporates microscale processes. Nat. Commun. 2018, 9, 2562. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Moncrieff, J.B. A model for soil CO2 production and transport 1: Model development. Agric. For. Meteorol. 1999, 95, 225–236. [Google Scholar] [CrossRef]
- Moyano, F.E.; Manzoni, S.; Chenu, C. Responses of soil heterotrophic respiration to moisture availability: An exploration of processes and models. Soil Biol. Biochem. 2013, 59, 72–85. [Google Scholar] [CrossRef]
- Langmuir, D. Aqueous Environmental Geochemistry; Prentice Hall: Upper Saddle River, NJ, USA, 1997. [Google Scholar]
- Stumm, W.; Morgan, J.J. Aquatic Chemistry: Chemical Equilibria and Rates in Natural Waters, 3rd ed.; Wiley: New York, NY, USA, 2012. [Google Scholar]
- Chapin, F.S.; Woodwell, G.M.; Randerson, J.T.; Rastetter, E.B.; Lovett, G.M.; Baldocchi, D.D.; Clark, D.A.; Harmon, M.E.; Schimel, D.S.; Valentini, R.; et al. reconciling carbon-cycle concepts, terminology, and methods. Ecosystems 2006, 9, 1041–1050. [Google Scholar] [CrossRef]
- Lovley, D.R. Organic matter mineralization with the reduction of ferric iron: A review. Geomicrobiol. J. 1987, 5, 375–399. [Google Scholar] [CrossRef]
- Weber, K.A.; Achenbach, L.A.; Coates, J.D. Microorganisms pumping iron: Anaerobic microbial iron oxidation and reduction. Nat. Rev. Microbiol. 2006, 4, 752. [Google Scholar] [CrossRef]
- Grybos, M.; Davranche, M.; Gruau, G.; Petitjean, P.; Pédrot, M. Increasing pH drives organic matter solubilization from wetland soils under reducing conditions. Geoderma 2009, 154, 13–19. [Google Scholar] [CrossRef]
- Lambert, T.; Pierson-Wickmann, A.-C.; Gruau, G.; Jaffrezic, A.; Petitjean, P.; Thibault, J.-N.; Jeanneau, L. Hydrologically driven seasonal changes in the sources and production mechanisms of dissolved organic carbon in a small lowland catchment. Water Resour. Res. 2013, 49, 1–12. [Google Scholar] [CrossRef]
- Knorr, K.-H. DOC-dynamics in a small headwater catchment as driven by redox fluctuations and hydrological flow paths-are DOC exports mediated by iron reduction/oxidation cycles? Biogeosciences 2013, 10, 891–904. [Google Scholar] [CrossRef]
- Dupas, R.; Gruau, G.; Gu, S.; Humbert, G.; Jaffrézic, A.; Gascuel-Odoux, C. Groundwater control of biogeochemical processes causing phosphorus release from riparian wetlands. Water Res. 2015, 84, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Jeanneau, L.; Jaffrezic, A.; Pierson-Wickmann, A.-C.; Gruau, G.; Lambert, T.; Petitjean, P. Constraints on the sources and production mechanisms of dissolved organic matter in soils from molecular biomarkers. Vadose Zone J. 2014, 13. [Google Scholar] [CrossRef]
- Lambert, T.; Pierson-Wickmann, A.-C.; Gruau, G.; Jaffrezic, A.; Petitjean, P.; Thibault, J.N.; Jeanneau, L. DOC sources and DOC transport pathways in a small headwater catchment as revealed by carbon isotope fluctuation during storm events. Biogeosciences 2014, 11, 3043–3056. [Google Scholar] [CrossRef]
- Lambert, T.; Pierson-Wickmann, A.-C.; Gruau, G.; Thibault, J.-N.; Jaffrezic, A. Carbon isotopes as tracers of dissolved organic carbon sources and water pathways in headwater catchments. J. Hydrol. 2011, 402, 228–238. [Google Scholar] [CrossRef]
- Denis, M.; Jeanneau, L.; Pierson-Wickman, A.-C.; Humbert, G.; Petitjean, P.; Jaffrézic, A.; Gruau, G. A comparative study on the pore-size and filter type effect on the molecular composition of soil and stream dissolved organic matter. Org. Geochem. 2017, 110, 36–44. [Google Scholar] [CrossRef]
- Jeanneau, L.; Denis, M.; Pierson-Wickmann, A.-C.; Gruau, G.; Lambert, T.; Petitjean, P. Sources of dissolved organic matter during storm and inter-storm conditions in a lowland headwater catchment: Constraints from high-frequency molecular data. Biogeosciences 2015, 12, 4333–4343. [Google Scholar] [CrossRef]
- Morel, B.; Durand, P.; Jaffrezic, A.; Gruau, G.; Molenat, J. Sources of dissolved organic carbon during stormflow in a headwater agricultural catchment. Hydrol. Process. 2009, 23, 2888–2901. [Google Scholar] [CrossRef]
- Fovet, O. ORE AgrHys. Vadose Zone J. 2018. [Google Scholar] [CrossRef]
- Gaillardet, J. RI OZCAR. Vadose Zone J. 2018. [Google Scholar] [CrossRef]
- IUSS Working Group WRB World Reference base for Soil Resources; FAO: Rome, Italy, 2006.
- Clark, J.M.; Ashley, D.; Wagner, M.; Chapman, P.J.; Lane, S.N.; Evans, C.D.; Heathwaite, A.L. Increased temperature sensitivity of net DOC production from ombrotrophic peat due to water table draw-down. Glob. Chang. Biol. 2009, 15, 794–807. [Google Scholar] [CrossRef]
- Christ, M.J.; David, M.B. Temperature and moisture effects on the production of dissolved organic carbon in a Spodosol. Soil Biol. Biochem. 1996, 28, 1191–1199. [Google Scholar] [CrossRef]
- Kätterer, T.; Reichstein, M.; Andrén, O.; Lomander, A. Temperature dependence of organic matter decomposition: A critical review using literature data analyzed with different models. Biol. Fertil. Soils 1998, 27, 258–262. [Google Scholar] [CrossRef]
- Creed, I.F.; Webster, K.L.; Braun, G.L.; Bourbonnière, R.A.; Beall, F.D. Topographically regulated traps of dissolved organic carbon create hotspots of soil carbon dioxide efflux in forests. Biogeochemistry 2013, 112, 149–164. [Google Scholar] [CrossRef]
- Gu, S.; Gruau, G.; Dupas, R.; Rumpel, C.; Crème, A.; Fovet, O.; Gascuel-Odoux, C.; Jeanneau, L.; Humbert, G.; Petitjean, P. Release of dissolved phosphorus from riparian wetlands: Evidence for complex interactions among hydroclimate variability, topography and soil properties. Sci. Total Environ. 2017, 598, 421–431. [Google Scholar] [CrossRef]
- Deng, Q.; Hui, D.; Chu, G.; Han, X.; Zhang, Q. Rain-induced changes in soil CO2 flux and microbial community composition in a tropical forest of China. Sci. Rep. 2017, 7, 5539. [Google Scholar] [CrossRef]
- Chow, A.T.; Tanji, K.K.; Gao, S.; Dahlgren, R.A. Temperature, water content and wet–dry cycle effects on DOC production and carbon mineralization in agricultural peat soils. Soil Biol. Biochem. 2006, 38, 477–488. [Google Scholar] [CrossRef]
- Buysse, P.; Flechard, C.R.; Hamon, Y.; Viaud, V. Impacts of water regime and land-use on soil CO2 efflux in a small temperate agricultural catchment. Biogeochemistry 2016, 130, 267–288. [Google Scholar] [CrossRef]
- Meyer, N.; Welp, G.; Amelung, W. The temperature sensitivity (q10) of soil respiration: Controlling factors and spatial prediction at regional scale based on environmental soil classes. Glob. Biogeochem. Cycles 2018, 32, 306–323. [Google Scholar] [CrossRef]
- Davidson, E.A.; Janssens, I.A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 2006, 440, 165. [Google Scholar] [CrossRef] [PubMed]
- Davidson, E.A.; Janssens, I.A.; Luo, Y. On the variability of respiration in terrestrial ecosystems: Moving beyond Q10. Glob. Chang. Biol. 2005, 12, 154–164. [Google Scholar] [CrossRef]
- Ruamps, L.S.; Nunan, N.; Pouteau, V.; Leloup, J.; Raynaud, X.; Roy, V.; Chenu, C. Regulation of soil organic C mineralisation at the pore scale. FEMS Microbiol. Ecol. 2013, 86, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Schimel, J.P. Effects of drying–rewetting frequency on soil carbon and nitrogen transformations. Soil Biol. Biochem. 2002, 34, 777–787. [Google Scholar] [CrossRef]
- Worrall, F.; Gibson, H.S.; Burt, T.P. Production vs. solubility in controlling runoff of DOC from peat soils—The use of an event analysis. J. Hydrol. 2008, 358, 84–95. [Google Scholar] [CrossRef]
- Kalbitz, K.; Solinger, S.; Park, J.-H.; Michalzik, B.; Matzner, E. Controls on the Dynamics of Dissolved Organic Matter in Soils: A Review. Soil Sci. 2000, 165, 277–304. [Google Scholar] [CrossRef]
- Estop-Aragonés, C.; Knorr, K.-H.; Blodau, C. Controls on in situ oxygen and dissolved inorganic carbon dynamics in peats of a temperate fen. J. Geophys. Res. Biogeosciences 2012, 117. [Google Scholar] [CrossRef]
- Tété, E.; Viaud, V.; Walter, C. Organic carbon and nitrogen mineralization in a poorly-drained mineral soil under transient waterlogged conditions: An incubation experiment. Eur. J. Soil Sci. 2015, 66, 427–437. [Google Scholar] [CrossRef]
- Thomsen, U.; Thamdrup, B.; Stahl, D.A.; Canfield, D.E. Pathways of organic carbon oxidation in a deep lacustrine sediment, Lake Michigan. Limnol. Oceanogr. 2004, 49, 2046–2057. [Google Scholar] [CrossRef]
- Roden, E.E.; Wetzel, R.G. Organic carbon oxidation and suppression of methane production by microbial Fe(III) oxide reduction in vegetated and unvegetated freshwater wetland sediments. Limnol. Oceanogr. 1996, 41, 1733–1748. [Google Scholar] [CrossRef]
- De-Campos, A.B.; Huang, C.; Johnston, C.T. Biogeochemistry of terrestrial soils as influenced by short-term flooding. Biogeochemistry 2012, 111, 239–252. [Google Scholar] [CrossRef]
- McNicol, G.; Silver, W.L. Separate effects of flooding and anaerobiosis on soil greenhouse gas emissions and redox sensitive biogeochemistry. J. Geophys. Res. Biogeosciences 2014, 119, 557–566. [Google Scholar] [CrossRef]
- Huang, W.; Hall, S.J. Elevated moisture stimulates carbon loss from mineral soils by releasing protected organic matter. Nat. Commun. 2017, 8, 1774. [Google Scholar] [CrossRef]
- Keiluweit, M.; Wanzek, T.; Kleber, M.; Nico, P.; Fendorf, S. Anaerobic microsites have an unaccounted role in soil carbon stabilization. Nat. Commun. 2017, 8, 1771. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Kdown | Kup | Gdown | Gup | |
|---|---|---|---|---|
| Land-use | Riparian area | Cropland (maize) a | Grass strip | Grass strip |
| Soil type (WRB 2006†) | Haplic Albeluvisol | Cambisol | Haplic Albeluvisol | Haplic Albeluvisol |
| Clay-Silt-Sand (%) | 29–60–11 | 21–69–10 | 24–60–16 | 23–61–16 |
| Soil bulk density (g cm−3) | 1.22 ± 0.16 | 1.30 ± 0.07 | NA | 1.14 ± 0.04 |
| (mean ± s. d.) | ||||
| Drainage classb | G | WD | G | PD |
| Number of days where the water table was above a soil depth of 10 cmc | 181 | 80 | 39 | 11 |
| Corg (g kg−1) | 60.2 | 18.8 | 37.6 | 23.7 |
| Ntotal (g kg−1) | 4.66 | 1.74 | 3.38 | 2.3 |
| C:N ratio | 12.9 | 10.8 | 11.1 | 10.3 |
| pH of soil solutionsd | 6.9 ± 0.3 * | 7.0 ± 0.3 * | 6.6 ± 0.3 ** | 6.6 ± 0.4 ** |
| Fe total (g 100 g−1 soil) | 0.771 | 0.603 | 0.453 | 0.491 |
| R2 | Slope | RMSE | Q10 | a | b | N | |
|---|---|---|---|---|---|---|---|
| All points | 0.47 | 0.44 | 1.3 | 2.06 ± 0.47 | 0.0013 | 0.145 | 71 |
| Kdown | 0.79 | 0.97 | 0.76 | 5.36 ± 2.90 | 0.0013 | 0.142 | 19 |
| Kup | 0.63 | 0.55 | 0.31 | 1.87 ± 0.35 | 0.0002 | 0.037 | 14 |
| Gdown | 0.73 | 0.65 | 1.48 | 2.32 ± 0.97 | 0.0013 | 0.151 | 19 |
| Gup | 0.72 | 0.68 | 1.01 | 3.14 ± 1.04 | 0.0009 | 0.122 | 19 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeanneau, L.; Buysse, P.; Denis, M.; Gruau, G.; Petitjean, P.; Jaffrézic, A.; Flechard, C.; Viaud, V. Water Table Dynamics Control Carbon Losses from the Destabilization of Soil Organic Matter in a Small, Lowland Agricultural Catchment. Soil Syst. 2020, 4, 2. https://doi.org/10.3390/soilsystems4010002
Jeanneau L, Buysse P, Denis M, Gruau G, Petitjean P, Jaffrézic A, Flechard C, Viaud V. Water Table Dynamics Control Carbon Losses from the Destabilization of Soil Organic Matter in a Small, Lowland Agricultural Catchment. Soil Systems. 2020; 4(1):2. https://doi.org/10.3390/soilsystems4010002
Chicago/Turabian StyleJeanneau, Laurent, Pauline Buysse, Marie Denis, Gérard Gruau, Patrice Petitjean, Anne Jaffrézic, Chris Flechard, and Valérie Viaud. 2020. "Water Table Dynamics Control Carbon Losses from the Destabilization of Soil Organic Matter in a Small, Lowland Agricultural Catchment" Soil Systems 4, no. 1: 2. https://doi.org/10.3390/soilsystems4010002
APA StyleJeanneau, L., Buysse, P., Denis, M., Gruau, G., Petitjean, P., Jaffrézic, A., Flechard, C., & Viaud, V. (2020). Water Table Dynamics Control Carbon Losses from the Destabilization of Soil Organic Matter in a Small, Lowland Agricultural Catchment. Soil Systems, 4(1), 2. https://doi.org/10.3390/soilsystems4010002