Abstract
As described in Japanese essay (Hojoki), written around 1200, various disasters such as big fires, earthquakes, and famines have occurred in Japan. Asian countries have been suffering from the disasters; furthermore, natural disasters are increasing due to global warming. Because tropical-infectious diseases are often disaster-related infectious diseases (DRIDs), the strategies against the former kind of disease could be applicable to DRIDs. Meteorological analysis of the occurrence of DRIDs using a method of time series analysis is important. In situations of disasters, it is desirable if you can identify the pathogen and identify disease severity simultaneously. A dipstick DNA chromatography assay termed as Single-Tag Hybridization—Printed Array Strip (STH—PAS) system was developed based on the DNA sequences of various mosquito-borne diseases. The plasma levels of matricellular proteins including galectin-9 (Gal-9) and osteopontin (OPN) were found to reflect the disease severities in the dengue virus and other DIRDs. Because both proteins have been reported to be immune-check molecules, their inhibition might enhance the immune system against pathogens. We found that brefelamide derivatives could inhibit OPN and other inflammatory molecules synthesis. Very recently, different derivatives were found to inhibit PD-L1 transcription. Applications of these agents should be considered as multi-step strategies against DRIDs.
Keywords:
disaster; tropical; infectious diseases; global warmings; STH-PAS; galectin 9; osteopontin; brefelamide 1. Introduction
Hojoki is one of the oldest essays in Japan and describes a big fire (1177), a tornado followed by relocation of the capital (1180), a famine (1181-2) and an earthquake (1185). In the famine, 42,300 people died in Kyoto city, and the city was filled with corpses. The author Chomei Kamono, a son of priest of Shimogamo Shrine at Kyoto city, decided to live in a small hermitage in nature, after experiencing these various kinds of disasters. Other studies also indicate that we have been suffering from big disasters in Japan. Recently at the Great East Japan Earthquake and Tsunami (GEJET) of March 11, 2011, a magnitude 9.0 earthquake attacked Sendai and neighboring cities, and the number of missing people was about 20,000. This area was found to be attacked by a tsunami (Jogan) at July 13, 869, indicating a large-scale tsunami occurs with a 1000 year interval [1,2].
The impacts of natural disasters have been growing in the world. Leaning and Guha-Sapir described it as follows: “There were three times as many natural disasters from 2000 through 2009 as there were from 1980 through 1989”. The economic damage brought by natural disasters shows that the world suffers catastrophic disasters every several years [3]. Since 1990, natural disasters have affected about 217 million people per year worldwide. The immediate and longer-term effects of these disruptions on large populations constitute humanitarian crises.
In 2018, there were 315 natural disaster events recorded with 11,804 deaths, over 68 million people affected, and US$131.7 billion in economic losses across the world. Flooding affected the highest number of people, accounting for 50% of the total affected, followed by storms which accounted for 28%. In terms of economic losses, storms were the costliest type of disaster. In the United States, hurricanes Florence and Michael cost $14 billion and $16 billion respectively, while in Asia, China, India, Japan, and the Philippines faced extensive damage from multiple storms. The costliest storm was typhoon Jebi in Japan ($12.5 billion) (Figure 1) [4].

Figure 1.
The damages caused by Typhoon Jebi (2018, July) at Okayama prefecture, Japan. Many houses (a), roads (b) and a car (c) were flooded. A factory exploded due to a chemical reaction to water (d). Roads and railroad tracks were destroyed by flood (e,f).
After disaster, communicable diseases associated with natural disasters, such as water-related communicable diseases, diseases associated with crowding, and vector borne diseases, are known to occur [5]. In the flood of China, after controlling the other meteorological and socioeconomic factors, malaria, diarrhea, and hepatitis A virus (HAV) infection were significantly related to the flood, both from the spatial and temporal analyses [6]. Tetanus outbreaks were also reported after a tsunami in Indonesia [7]. Global warming is a major cause of these disasters. The signs of climate change are undeniable, and the inevitable impact for Earth and all its inhabitants is a serious concern. Because of the tight associations between climate change and human health, some of the health care challenges are posed by a changing climate [8]. Ice is melting, sea levels are rising, biodiversity is declining, precipitation has increased, atmospheric levels of carbon dioxide and greenhouse gases are alarmingly high, and extreme weather conditions are becoming increasingly common. Global warming, extreme weather, flooding, and other consequences of climate change on microbial communities in the ocean and soil have been proposed [9]. Using an empirically-informed, process-based mathematical model to study the feasibility of Aedes aegypti infestation into continental Europe. It was found that in a low carbon emission future (RCP2.6), minimal change to the current situation throughout the whole of the 21st century. However, in a high carbon future (RCP8.5), a large parts of southern Europe risks being invaded by A aegypti [10]. Furthermore, the links between climate variations and the emergence of climate-sensitive infectious diseases were described, such as the mosquito-borne diseases dengue, chikungunya, and Zika [11]. Our comparative study indicated that significant disasters in the Philippines were geophysical, hydrological, or meteorological, whereas the significant disasters in Indonesia were geophysical or hydrological. Leptospirosis, dengue, diarrhea, and cholera were recognized as common disaster-related infections in the Philippines, whereas diarrhea and respiratory infection were the major disaster-related infections in Indonesia. These results indicate that disaster-related infectious diseases (DRIDs) differ from place to place [12].
2. Studies of Disaster-Related Infectious Disease
2.1. Infectious Disease Itself Is a Disaster
In turn, infectious diseases could themselves be regarded as constituting natural disasters. Some microorganisms have been called natural weapons. For example, human immunodeficiency virus (HIV) infection as well as cigarette smoking are two major novel causes of mortality which both emerged in the twentieth century [13]. Such “weapons” could be a cause of disasters, if they are not treated appropriately. In South Africa, the number of deaths caused by acquired immune deficiency syndrome (AIDS) associated with tuberculosis (TB) in 1990 accounted for 47% of all deaths, and an increase in HIV infections was also noted during this year [2]. Increased HIV infections were seen in South Africa in 1990, indicating the invasion of HIV into TB-endemic area, which accounts for much of the rise of AIDS/TB in South Africa [14]. Moreover, extensively drug-resistant (XDR)-TB emerged and was reported for the first time in a society in KwaZulu-Natal in South Africa [15].
HIV/tuberculosis co-infections has been one of the biggest issues in global health because HIV infected patients have 5–10 times more risk of developing tuberculosis than HIV uninfected individuals in spite of their treatment with anti-retroviral drugs [16]. We should be careful with regard to the occurrence of an AIDS/TB epidemic in Southeast Asia. We found that latent TB infection rate is quite high in the Philippines [17] and in Indonesia as well [18]. The rate of HIV infection has been rising alarmingly over the past seven years in the subpopulation of men who have sex with men (MSM) in the Philippines. In the year 2014 alone, there were 6011 newly diagnosed cases of HIV, representing 27% of the total number of cases identified in the Philippines (22,527) since the epidemic began in 1984 [19]. The Philippines has the fastest growing HIV epidemic in the world. According to UNAIDS, there was a 203% increase in the rate of new infections between 2010 and 2018, concentrated among men who have sex with men (MSM), drug users, and sex workers. MSM also have female partners, as do drug users. Rates among pregnant women are also rising [20].
The HIV invasion in a TB endemic country inevitably results in an increase of AIDS/TB as described above. The immunosuppressive state induced by HIV infection has been shown to hinder immune responses in tuberculosis infection affecting granuloma formation and fibrosis in HIV/tuberculosis co-infected patients. Hence, although these patients are at a higher risk of developing extra-pulmonary tuberculosis, lung damage in pulmonary tuberculosis is limited in them. Both these infections involve chronic immune activation, ultimately responsible for immunopathogenesis of the diseases. Immune activation has been shown to be the central mechanism responsible for disease progression in HIV infection. Similarly, chronic inflammation in pulmonary tuberculosis is responsible for granuloma formation and fibrosis [21].
Research on the molecular diagnosis of drug-resistant TB facilitated the development of the Xpert MTB/RIF and line probe assays, both of which efficiently identify rifampicin resistant (RR)- and multi-drug resistant (MDR)-TB [22]. The National TB program (NTP) in the Philippines introduced the GeneXpert MTB/RIF assay in San Lazaro Hospital (SLH), in Manila, Philippines, as recommended by the WHO. Among all the cases in 2015 that were presumptively screened for TB (n = 1052), 122 (11%) were found to have RR-TB. However, 27% (n = 32) of RR-TB patients who were identified using the Xpert MTB/RIF assay were not enrolled: 10% (n = 12) refused to revisit for enrollment after phone calls and home visits, 8% (n = 10) died within two weeks of diagnosis, 7% (n = 8) could not be located/relocated, and 2% (n = 2) were treated with the first-line drugs at the local health center, despite the diagnosis of RR-TB. There has been increase in the detection of HIV-positive MDR-TB patients from 2011 (n = 1) to 2015 (n = 14). A total of 37 cases were identified, and the identification rate gradually increased over time: 0.5% in 2011, 3% in 2012, 5% in 2013, 9% in 2014, and 15% in 2015. The HIV-positive rate among MDR-TB patients was increased 30-fold in 2015. The mortality rate was high among the AIDS/MDR-TB patients, and ranged from 36% to 67% (average, 46%). The cause of death was primarily severe sepsis or meningitis among the cases with fatal outcomes [23,24]. One of the reasons of high MDR-TB rates in the Philippines could be attributed to patient loss. Independent factors associated with loss to follow-up included patients’ higher self-rating of the severity of vomiting as an adverse drug reaction and alcohol abuse [25]. It is known that most prevalent genotype of all MTB (37 cases) belonged to Manila type at 2014 [26] and recent whole genome sequencing clarified that the majority (80.3%) belonged to lineage 1 Manila clade with the minority belonging to lineages 2 and 4 [27]. It is difficult to conclude if new strains have been emerging because the locations of two studies are not the same.
2.2. Infectious Diseases Caused by Disaster in Global Warming
In daily life, human health is guarded from microorganisms by many systems and devices, such as tap water, gas heating, washing, and refrigeration. Disasters destroy these protective systems, and the victims will be exposed to natural threats such as contaminated water, animals, and disease vectors immediately after the disaster. After a while, human-to-human communicable diseases occur due to the dense lives of the victims in affected camp. In the Republic of the Philippines, dengue fever (DF) and leptospirosis are recognized as common DRIDs, in addition to diarrhea and cholera. A clear relationship has been observed between an increase in DF cases and the southwest and northeast monsoons, particularly in Manila, in the Philippines. An earlier study addressed the reasons for DF epidemics in terms of rainfall [28]. We assumed that DF epidemics are correlated not only with rainfall but also relative humidity and temperature by using a method of time series analysis. DF during a week was related to rainfall over the prior 6–7 weeks (Figure 2a) [29]. This can be attributed to the life-cycle duration of mosquitoes and the requirement of an adequate number of cases for spread, which is in turn affected by population density [30]. Moreover, leptospirosis associated with floodwaters following monsoons in Manila and the peak occurrence of leptospirosis preceded that of DF by only one month (Figure 2b), though it occurs 2–3 months later than the peak occurrence of dengue in Thailand [29]. It should also be noted that Philippines has reported outbreaks of dengue, diphtheria, measles, and polio, which might be linked with dengue vaccine failure in which dengue-seronegative children vaccinated with Dengvaxia had higher chances of subsequent hospitalization for dengue than seropositive children [31]. Above situations indicate the effective management of these tropical infectious diseases could lead to the management of DRIDs as well.
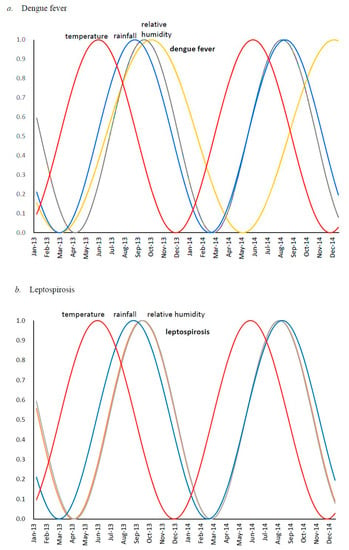
Figure 2.
Seasonal cycles obtained from spectral analysis based on maximum entropy method, for: (a) dengue fever (orange line), temperature (red line), rainfall (blue line) and relative humidity (grey line); (b) leptospirosis (orange line), temperature (red line), rainfall (blue line) and relative humidity (grey line) overlap [24].
2.3. STH-PAS as Point-Of-Care Testing for Disaster-Related Infectious diseases
Dengue virus (DENV) is the cause of DF and is a mosquito-borne single positive-stranded RNA virus and is a family of Flaviviridae. Patients with DF are seen in most of the tropical and subtropical areas of the world, and their numbers has increased dramatically within the last 20 years. Current estimation indicates that nearly 390 million infections occur each year [32].
Symptoms begins 3~14 days after infection and leads to a broad spectrum of clinical symptoms and severity, including asymptomatic infection, DF, and fatal dengue hemorrhagic fever (DHF). DF/DHF is considered to be one of the most important reemerging infectious diseases. The characteristic symptom of dengue are sudden-onset fever, headache, muscle and joint pains and a rash. The diagnosis of dengue is typically made clinically, however, early disease can be difficult to differentiate from other infections such as chikungunya and Zika which are also Flaviviruses transmitted by mosquitoes.
Several laboratory diagnostic techniques have been used for the confirmation of dengue virus infection: viral isolation, viral antigen detection, viral genome detection, and antibody (Ab) detection.
IgM capture enzyme-linked immunosorbent assay (ELISA) and reverse transcriptase PCR (RT-PCR) are commonly used. NS-1 antigen detection tests have also recently become commercially available [33].
Detection of the DENV genome in serum samples by RT-PCR is efficient at the early stage of the disease, however, an expensive apparatus and reagents are necessary to conduct the assay, and the final diagnosis can be delayed because the specimen should be sent to the laboratory [2].
We developed a novel testing, dengue virus RNA detection system using the single tag hybridization chromatographic printed array strip (STH-PAS) method, which affords the visualization of the signals in a single reaction with great sensitivity and in a dramatically short time. Moreover, it does not require the preparation or staining of a gel and an expensive apparatus (Figure 3) [34]. The methods allow us to detect several genes in one strip so that we can also identify subtypes of DENVs.
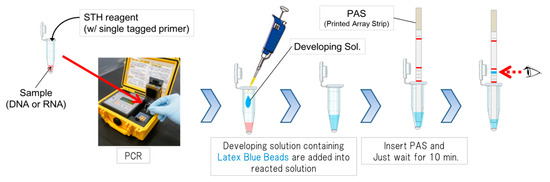
Figure 3.
A schematic illustration of single tag hybridization chromatic printed array strip (STH-PAS) system.
We evaluated the sensitivity and specificity of the kit as a clinical research using the samples from other infectious diseases as well as dengue infection. In the core technology “STH-PAS”, it is a feature that single-stranded tag DNA acting as marker is bound to PCR primer and amplified by PCR. The PCR amplicon has single-stranded tag, which is chromatographed on the printed array strip (PAS) and is trapped by complementary DNA printed in advance in line shape. That is, if the target gene is present in the sample, it is detected as blue line in the PAS. The keynote that the gene can be detected by such a simple visible result is cutting-edge and unique.
Sensitivities against serially diluted-DENV subtypes were detected by STH-PAS Dengue Subtype At Once kit using the primers which could detect all subtypes [34]. Virus RNA was obtained using QIAamp Viral RNA Mini Kit (QIAGEN, GmbH, Germany) according to manufacturer’s instructions. We added two µL of viral RNA extract to 8 µL of OneStep RT-PCR Master Mix (QIAGEN, GmbH, Germany) in combination with a tag-spacer sequence and biotin-labeled primer sequence (AVSS Co., Ltd., Japan).
The primer could detect all four serotypes of DENV (Figure 4). The detection limit of copy numbers is shown in Table 1. The results showed that we could detect all subtypes of DENV in one strip and with very high sensitivity. The extensive studies using large clinical samples are now ongoing.
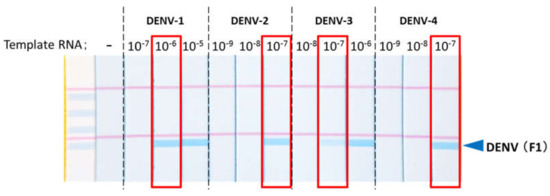
Figure 4.
The primer for dengue virus (DENV) could detect all DENV serotypes at low concentration. Each virus culture supernatant was detected by STH-PAS dengue virus universal detection kit.

Table 1.
Detection limit copy numbers of STH-PAS against different subtypes.
It should be noted that accurate and sensitive genetic diagnosis of multiple pathogens is possible using STH-PAS.
2.4. MCP Proteins as Severity Markers
When one suffers from infectious diseases, edema is the most common symptom of the affected organs. Matricellular proteins (MCPs) are important during wound repair, interact with cell surface receptors, (such as integrin) and are able to bind to growth factors. Based on this definition, several proteins have now been identified as MCPs [35], including galectins [36], and osteopontin (OPN) [37].
Galectin-9 (Gal-9), a member of galectin-family of β-galactoside binding animal lectin, is expressed ubiquitously with abundance in the epithelium of gastrointestinal tract and immune cells. Gal-9 is annotated as an immune checkpoint molecule because of its immune suppressive function by binding to T-cell immunoglobulin and mucin domain 3 (Tim-3) and attenuating Th1 and Th17 biased inflammation [38]. Pharmaceutical companies are conducting research to neutralize this lectin with the aim of enhancing immunity against cancer and/or infection. However, Gal-9 binds several other target molecules including adhesion molecule CD44 [39], one of TNF receptor superfamily 4-1BB [40], integrins [41] and IgE [42], and exerts various functions depending on the target, which could induce inflammation in certain situations. The effects of Gal-9 in infectious diseases have been studied using recombinant protein and/or Gal-9 deficient mouse. As expected from the anti-inflammatory function, Gal-9 reduced Tim-3+ T cells, supposedly Th1(interferon-γ secreting cells), Th17 (IL-17 secreting cells) and CD8 (cytotoxic T lymphocyte) with concomitant induction of Treg in several reports. This regulatory function of Gal-9 limited bystander tissue damage and improved the lesion or survival in the models of coxsackievirus B3 induced myocarditis [43], herpes simplex virus type 1(HSV-1) induced keratitis [44] and polymicrobial sepsis [45]. Naturally these immune suppressive environments allowed pathogens to infect and persist more efficiently as was demonstrated in HSV-1 [46], hepatitis c virus (HCV) [47] and Klebsiella pneumoniae infection [48]. In HCV study, Gal-9 was demonstrated to be the causative agent of tissue inflammation and damage by inducing inflammatory cytokines from Kupffer cells [47]. Gal-9 was shown to bind Pseudomonas aeruginosa directly via its lipopolysaccharide, which opsonizes the germ to stimulate phagocytosis by neutrophils [49]. Gal-9 also act directly to infected cells, for example, Gal-9 induced cycline-dependent kinase inhibitor p21 via Tim-3 signaling and attenuated infection and propagation of the HIV [50]. Further, Gal-9 induced APOBEC3, a mutagenic enzyme of the HIV genome, and reduced the infectivity of the progeny virus [51]. In Mycobacterium tuberculosis infection, Gal-9 expressed on the surface of infected macrophages worked as a receptor for Tim-3 expressed by Th1. The interaction transduced signals to produce IL-1β which worked autocrine to the infected macrophage to kill ingested mycobacterium [52]. In a collaborative study with SLH, we have been clarifying the role of Gal-9 and OPN in DENV infection. During the critical phase, Gal-9 levels (GalPharma (GalP), Kagawa, Japan) were significantly higher in DENV-infected patients than in healthy persons or those with non-dengue febrile illness. The highest Gal-9 levels were observed in patients with dengue hemorrhagic fever (DHF): 2464 pg/mL; patients with DF: 1407 pg/mL. In the recovery phase, Gal-9 levels significantly declined from peak levels in both DF and DHF patients. We proposed that Gal-9 could serve as an important novel biomarker of acute DENV infection and reflect disease severity [33]. We have also clarified that Gal-9 levels could reflect the severity in malaria infection, which is also a representative DRID [53]. We have examined if another MCP, OPN, is involved in acute virus infections. OPN is a highly phosphorylated and glycosylated MCP and proteolytic cleavage of OPN by thrombin (between Arg168 and Ser169) generates a functional fragment of N-terminal OPN (thrombin-cleaved OPN—trOPN). In DENV infection, the levels of OPN (IBL, Gunma, Japan), trOPN (IBL, Gunma, Japan), d-dimer, thrombin-antithrombin complex (TAT) protein, and thrombomodulin were significantly elevated in the critical phase in both patients with DF and those with DHF, as compared with healthy individuals. During the recovery phase, OPN levels declined, whereas trOPN levels further increased dramatically in both DF and DHF patients. However, the OPN levels measured by a different enzyme-linked immunosorbent assay (ELISA) (R&D Systems (RDS), Mineapolis, USA) remained greater in DENV-infected subjects than they were in HCs during the recovery phase. A Spearman rank correlation coefficient revealed a significant correlation between IBL and R&D Systems assessments of the DENV-infected patients during the critical phase. However, no correlation was evident during the recovery phase. It is known that the IBL ELISA kit only measured the full-length (FL)-OPN, and specifically did not measure the cleaved form, though the epitope of the OPN protein measured by the RDS kit was not disclosed and the kit probably measure both forms, but could not differentiate between them [54]. It has not been clear how the cleaved form of OPN would be generated in vivo, however various forms of OPN were identified in human monocytic THP-1 cells stimulated by phorbol 12-myristate 13-acetate (PMA) in vitro. Antibodies O-17, 10A16 and 34E3, which recognize N-terminus, the C-half, and thrombin-cleaved site of OPN, respectively, all detected distinct bands on Western blots following PMA stimulation. Bands corresponding to 18 and 30 kD were detected by antibodies 34E3 and 10A16, indicating that OPN cleavage occurred by endogenous proteases in the PMA-stimulated THP-1 cells [55]. OPN was identified by immunohistochemistry in macrophages, lymphocytes, and the extracellular matrix of pathologic tissue sections of patients with tuberculosis. The association of OPN with granulomatous pathology, together with the known properties of the protein, suggest that OPN may participate in granuloma formation [56].
We also confirmed a high level of plasma OPN in subjects with MTB from the Philippines [26] and from Indonesia [57]. (FL-OPN is involved in the complex pathways of coagulation and fibrinolysis, where FL-OPN serve as targets for thrombin cleavage. During this process, OPN fragments are produced. Among those fragments, proteolytic cleavage of FL-OPN by thrombin (between Arg168 and Ser169) generates a functional fragment of N-terminal thrombin-cleaved OPN (trOPN), which contains a cryptic binding site for integrins α9β1 and α4β1 that enhances the attachment of trOPN to integrins. Furthermore, other OPN forms are detected in DENV infections [54] or PMA-stimulated THP-1 cells as described above using a different ELISA system, which include a mixture of FL-OPN, and cleaved form of OPN and termed as undefined OPN (Ud-OPN) [55].
Higher plasma concentrations of Ud-OPN, but not FL-OPN or trOPN, negatively correlate with TB-specific memory T-cell numbers represented by IFN-γ-secreting cell numbers of ESAT-6 stimulated peripheral blood lymphocytes and positively correlate with neutrophil numbers, indicating generation of cleaved form of OPN is involved in granuloma formation [58].
It is known that MCPs modulate cell function by interacting with cell-surface receptors, proteases, hormones, and other bio-effector molecules. Therefore there is a caveat in ELISA measurement [59]. It was implicated that OPN levels measured by RDS could reflect the immune activation status of cells and host [55,58].
Examination of published concentration of blood Gal-9 in various diseases clarified stark differences even in healthy controls, with values ranging from 0 to 26,560 pg/mL. These large variations were attributed to the choice of ELISA kit, as the RDS kit demonstrated very high levels compared with other products including the kit from GalP of which we have been using in all the past studies. We compared RDS and GalP ELISA using a same set of plasma from patients of acute liver failure and healthy controls and found the following; (1) RDS ELISA indicated about 50-fold higher median Gal-9 levels than that indicated by GalP ELISA, (2) both measurements demonstrated significantly higher Gal-9 levels in patient group, however (3) the correlation between the two measurements was not strong. We found that RDS ELISA reacted aberrantly higher to degraded Gal-9 than the intact one and that the enhanced signals were mistranslated into a high concentration of Gal-9. Gal-9 is a fragile protein and it is likely that human specimens of any kinds will contain degraded Gal-9, which would distort the quantification by RDS ELISA, hence the kit has little value for quantification of Gal-9 [59].
2.5. Attempts to Controls MCP Proteins
Gal-9 and Tim3 were proposed to behave as co-inhibitor receptors on Th1 cells [60] and OPN was found to control CD8 cells in cancer by binding to its receptor CD44 [61] as immune check point molecule as well. Inhibition of these molecules should lead to an increase the immune system against infectious pathogens or infected cells. To seek this possibility, new inhibitors derived from brefelamide, an aromatic amide isolated from Dictyostelium cellular slime molds that may inhibit OPN transcription in A549 cells at 1 µM concentration, were synthesized as compounds C, D, and E (Figure 5). Their inhibitory activity against OPN synthesis in PMA-stimulated THP-1 cells was confirmed using ELISA, a multicolor immune-fluorescent microscope, and Western blot. In the ELISA performed using IBLELISA kit, each compound showed significant inhibition in culture supernatants with half maximal inhibitory concentration (IC50) values of 1.6, 1.8, and 2.2 µM for C, D, and E, respectively. The decreases in OPN expression and synthesis were confirmed using immunofluorescence and Western blot studies using compound-treated cells or cell lysates. Luminex assay of the supernatants of PMA-treated THP-1 cells showed significant reduction in the synthesis of interleukin (IL)-1β, galectin-9, and tumor necrosis factor (TNF)-α. Elucidation of the detailed mechanisms of the biological activities of these compounds would be necessary; however, they may be used in clinical trials for DRID, inflammatory disorders, and cancer [62].
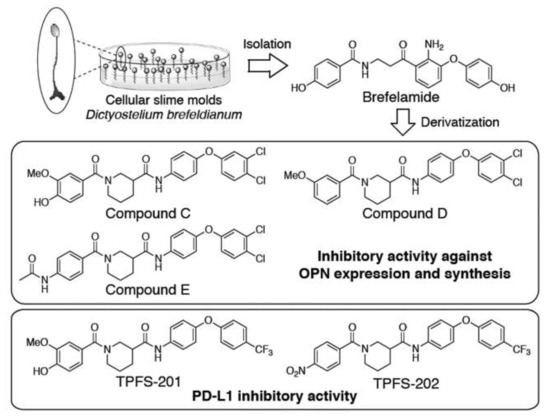
Figure 5.
Structures of compounds derived from slime mold. Compound C-E are from ref. [62] and TPFS-201 and 202 are from ref [63].
Very recently, we identified other two amide analogues of brefelamide, TPFS-201 and TPFS-202, as small molecular immune checkpoint inhibitors by downregulation of PD-L1 expression on tumor cells (Figure 5) [63]. The promoter activity, endogenous mRNA and surface protein expression of PD-L1 were suppressed in compound-treated cancer cells, which consequently restored T cell activity, as evidenced by diminished apoptosis and increased IL-2 promoter activity in Jurkat T cells co-cultured with compound-treated A549 cells. Compound-mediated PD-L1 inhibition was partially abolished by disruption of the putative TAZ/TEAD-binding motif in PD-L1 promoter. The inhibitory effect of TPFS-202 on PD-L1 was largely blunted in mouse cell lines, which is consistent with the previous finding that PD-L1 regulation by TAZ was not conserved in mouse cells due to distinct promoter sequences flanking TAZ/TEAD-binding motif. Together, our data indicate the potential utility of amide analogues of brefelamide as small molecule immune checkpoint inhibitors against DRID.
3. Conclusions
The research against tropical infectious diseases on improvement of molecular diagnosis and identification of severity biomarkers contributes to the management of DRID as well. New brefelamide derivatives may ameliorate the symptoms caused by exaggerated inflammation of DRID. International cooperation is vital to combat the diseases in the era of global warming.
Author Contributions
All author wrote the manuscript. G.B., H.K., A.S. and N.K. made the figures. T.H. edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Research Program on Emerging and Re-Emerging Infectious Diseases from the Japan Agency for Medical Research and Development (AMED JP18fk0108042h0002), and partially supported by the Japan Society for the Promotion of Science (JSPS) Grants-in-Aid for Scientific Research (KAKENHI), Grant Number JP17H01690.
Acknowledgments
We thank Keiji Mise of Sapporo Medical University for preparing Figure 2.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Shibahara, S. Revisiting the March 11, 2011 earthquake and tsunami: Resilience and restoration. Tohoku J. Exp. Med. 2012, 226, 1–2. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hattori, T.; Chagan-Yasutan, H.; Shiratori, B.; Egawa, S.; Izumi, T.; Kubo, T.; Nakajima, C.; Suzuki, Y.; Niki, T.; Alisjahbana, B.; et al. Development of point-of-care testing for disaster-related infectious diseases. Tohoku J. Exp. Med. 2016, 238, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Leaning, J.; Guha-Sapir, D. Natural disasters, armed conflict, and public health. N. Engl. J. Med. 2013, 369, 1836–1842. [Google Scholar] [CrossRef] [PubMed]
- Alexander, D. Natural Disasters; Routledge: Abingdon, UK, 2018. [Google Scholar]
- Watson, J.T.; Gayer, M.; Connolly, M.A. Epidemics after natural disasters. Emerg. Infect. Dis. 2007, 13, 1–5. [Google Scholar] [CrossRef]
- Gao, L.; Zhang, Y.; Ding, G.; Liu, Q.; Jiang, B. Identifying flood-related infectious diseases in Anhui Province, China: A spatial and temporal analysis. Am. J. Trop. Med. Hyg. 2016, 94, 741–749. [Google Scholar] [CrossRef]
- Pascapurnama, D.N.; Murakami, A.; Chagan-Yasutan, H.; Hattori, T.; Sasaki, H.; Egawa, S. Prevention of tetanus outbreak following natural disaster in indonesia: Lessons learned from previous disasters. Tohoku J. Exp. Med. 2016, 238, 219–227. [Google Scholar] [CrossRef]
- Rossiello, M.R.; Szema, A. Health effects of climate change-induced wildfires and heatwaves. Cureus 2019, 11, 4771. [Google Scholar] [CrossRef]
- Hutchins, D.A.; Jansson, J.K.; Remais, J.V.; Rich, V.I.; Singh, B.K.; Trivedi, P. Climate change microbiology—Problems and perspectives. Nat. Rev. Microbiol. 2019, 17, 391–396. [Google Scholar] [CrossRef]
- Liu-Helmersson, J.; Rocklov, J.; Sewe, M.; Brannstrom, A. Climate change may enable Aedes aegypti infestation in major European cities by 2100. Environ. Res. 2019, 172, 693–699. [Google Scholar] [CrossRef]
- Filho, W.L.; Scheday, S.; Boenecke, J.; Gogoi, A.; Maharaj, A.; Korovou, S. Climate change, health and mosquito-borne diseases: Trends and Implications to the pacific Region. Int. J. Environ. Res. Public Health 2019, 16, 5114. [Google Scholar] [CrossRef]
- Usuzawa, M.; Telan, E.O.; Kawano, R.; Dizon, C.S.; Alisjahbana, B.; Ashino, Y.; Egawa, S.; Fukumoto, M.; Izumi, T.; Ono, Y.; et al. Awareness of disaster reduction frameworks and risk perception of natural disaster: A questionnaire survey among Philippine and Indonesian health care personnel and public health students. Tohoku J. Exp. Med. 2014, 233, 43–48. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Weiss, R.A.; McMichael, A.J. Social and environmental risk factors in the emergence of infectious diseases. Nat. Med. 2004, 10, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Mee, P.; Wagner, R.G.; Gomez-Olive, F.X.; Kabudula, C.; Kahn, K.; Madhavan, S.; Collinson, M.; Byass, P.; Tollman, S.M. Changing use of traditional healthcare amongst those dying of HIV related disease and TB in rural South Africa from 2003–2011: A retrospective cohort study. BMC Complement. Altern. Med. 2014, 14, 504. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, N.R.; Moll, A.; Sturm, A.W.; Pawinski, R.; Govender, T.; Lalloo, U.; Zeller, K.; Andrews, J.; Friedland, G. Extensively drug-resistant tuberculosis as a cause of death in patients co-infected with tuberculosis and HIV in a rural area of South Africa. Lancet 2006, 368, 1575–1580. [Google Scholar] [CrossRef]
- Lawn, S.D.; Myer, L.; Bekker, L.G.; Wood, R. Burden of tuberculosis in an antiretroviral treatment programme in sub-Saharan Africa: Impact on treatment outcomes and implications for tuberculosis control. AIDS 2006, 20, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Siddiqi, U.R.; Leano, P.S.; Chagan-Yasutan, H.; Shiratori, B.; Saitoh, H.; Ashino, Y.; Suzuki, Y.; Hattori, T.; Telan, E.F. Frequent detection of anti-tubercular-glycolipid-IgG and -IgA antibodies in healthcare workers with latent tuberculosis infection in the Philippines. Clin. Dev. Immunol. 2012, 2012, 610707. [Google Scholar] [CrossRef] [PubMed]
- Senoputra, M.A.; Shiratori, B.; Hasibuan, F.M.; Koesoemadinata, R.C.; Apriani, L.; Ashino, Y.; Ono, K.; Oda, T.; Matsumoto, M.; Suzuki, Y.; et al. Diagnostic value of antibody responses to multiple antigens from Mycobacterium tuberculosis in active and latent tuberculosis. Diagn. Microbiol. Infect. Dis. 2015, 83, 278–285. [Google Scholar] [CrossRef]
- Ross, A.G.; Ditangco, R.A.; Belimac, J.G.; Olveda, R.M.; Mercado, E.S.; Chau, T.N.; Crowe, S.M. The dire sexual health crisis among MSM in the Philippines: An exploding HIV epidemic in the absence of essential health services. Int. J. Infect. Dis. 2015, 37, 6–8. [Google Scholar] [CrossRef]
- Devi, S. Stigma, politics, and an epidemic: HIV in the Philippines. Lancet 2019, 394, 2139–2140. [Google Scholar] [CrossRef]
- Lawn, S.D.; Butera, S.T.; Folks, T.M. Contribution of immune activation to the pathogenesis and transmission of human immunodeficiency virus type 1 infection. Clin. Microbiol. Rev. 2001, 14, 753–777. [Google Scholar] [CrossRef]
- Aurin, T.H.; Munshi, S.K.; Kamal, S.M.; Rahman, M.M.; Hossain, M.S.; Marma, T.; Rahman, F.; Noor, R. Molecular approaches for detection of the multi-drug resistant tuberculosis (MDR-TB) in Bangladesh. PLoS ONE 2014, 9, e99810. [Google Scholar] [CrossRef] [PubMed]
- Hattori, T.; Kobayashi, N.; Nagai, H.; Chagan-Yasutan, H.; Telan, E.; Solante, M.B. Nationwide HIV-, MDR-TB survey in Japan and collaborative study in the Philippines. Int. J. Mycobacteriol. 2016, 5, 18–19. [Google Scholar] [CrossRef] [PubMed]
- Solante, M.B.; Chagan-Yasutan, H.; Hattori, T.; Leano, S.; Saludar, N.R.; Gartin, A.M.C.G.; Soolinger, D.V.; Telan, E. High rates of human immunodeficiency virus and drug resistance in tuberculosis patients in Manila, Philippines. Biotechnol. Res. J. 2017, 1, 157–162. [Google Scholar] [CrossRef]
- Tupasi, T.E.; Garfin, A.M.; Kurbatova, E.V.; Mangan, J.M.; Orillaza-Chi, R.; Naval, L.C.; Balane, G.I.; Basilio, R.; Golubkov, A.; Joson, E.S.; et al. Factors associated with loss to follow-up during treatment for multidrug-resistant tuberculosis, the philippines, 2012–2014. Emerg. Infect. Dis. 2016, 22, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Shiratori, B.; Leano, S.; Nakajima, C.; Chagan-Yasutan, H.; Niki, T.; Ashino, Y.; Suzuki, Y.; Telan, E.; Hattori, T. Elevated OPN, IP-10, and neutrophilia in loop-mediated isothermal amplification confirmed tuberculosis patients. Mediat. Inflamm. 2014, 2014, 513263. [Google Scholar] [CrossRef] [PubMed]
- Phelan, J.E.; Lim, D.R.; Mitarai, S.; de Sessions, P.F.; Tujan, M.A.A.; Reyes, L.T.; Medado, I.A.P.; Palparan, A.G.; Naim, A.N.M.; Jie, S.; et al. Mycobacterium tuberculosis whole genome sequencing provides insights into the Manila strain and drug-resistance mutations in the Philippines. Sci. Rep. 2019, 9, 9305. [Google Scholar] [CrossRef]
- Su, G.L. Correlation of climatic factors and dengue incidence in Metro Manila, Philippines. Ambio 2008, 37, 292–294. [Google Scholar] [CrossRef]
- Sumi, A.; Telan, E.F.; Chagan-Yasutan, H.; Piolo, M.B.; Hattori, T.; Kobayashi, N. Effect of temperature, relative humidity and rainfall on dengue fever and leptospirosis infections in Manila, the Philippines. Epidemiol. Infect. 2017, 145, 78–86. [Google Scholar] [CrossRef]
- Bravo, L.; Roque, V.G.; Brett, J.; Dizon, R.; L’Azou, M. Epidemiology of dengue disease in the Philippines (2000–2011): A systematic literature review. PLoS Negl. Trop. Dis. 2014, 8, 3027. [Google Scholar] [CrossRef]
- The Lancet Infectious, D. Infectious disease crisis in the Philippines. Lancet Infect. Dis. 2019, 19, 1265. [Google Scholar] [CrossRef]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Chagan-Yasutan, H.; Ndhlovu, L.C.; Lacuesta, T.L.; Kubo, T.; Leano, P.S.; Niki, T.; Oguma, S.; Morita, K.; Chew, G.M.; Barbour, J.D.; et al. Galectin-9 plasma levels reflect adverse hematological and immunological features in acute dengue virus infection. J. Clin. Virol. 2013, 58, 635–640. [Google Scholar] [CrossRef] [PubMed]
- Liles, V.R.; Pangilinan, L.S.; Daroy, M.L.G.; Dimamay, M.T.A.; Reyes, R.S.; Bulusan, M.K.; Dimamay, M.P.S.; Luna, P.A.S.; Mercado, A.; Bai, G.; et al. Evaluation of a rapid diagnostic test for detection of dengue infection using a single-tag hybridization chromatographic-printed array strip format. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 515–521. [Google Scholar] [CrossRef]
- Bornstein, P.; Sage, E.H. Matricellular proteins: Extracellular modulators of cell function. Curr. Opin. Cell Biol. 2002, 14, 608–616. [Google Scholar] [CrossRef]
- Elola, M.T.; Wolfenstein-Todel, C.; Troncoso, M.F.; Vasta, G.R.; Rabinovich, G.A. Galectins: Matricellular glycan-binding proteins linking cell adhesion, migration, and survival. Cell Mol. Life Sci. 2007, 64, 1679–1700. [Google Scholar] [CrossRef]
- Murphy-Ullrich, J.E.; Sage, E.H. Revisiting the matricellular concept. Matrix Biol. 2014, 37, 1–14. [Google Scholar] [CrossRef]
- Zhu, C.; Anderson, A.C.; Schubart, A.; Xiong, H.; Imitola, J.; Khoury, S.J.; Zheng, X.X.; Strom, T.B.; Kuchroo, V.K. The Tim-3 ligand galectin-9 negatively regulates T helper type 1 immunity. Nat. Immunol. 2005, 6, 1245–1252. [Google Scholar] [CrossRef]
- Katoh, S.; Ishii, N.; Nobumoto, A.; Takeshita, K.; Dai, S.Y.; Shinonaga, R.; Niki, T.; Nishi, N.; Tominaga, A.; Yamauchi, A.; et al. Galectin-9 inhibits CD44-hyaluronan interaction and suppresses a murine model of allergic asthma. Am. J. Respir. Crit. Care Med. 2007, 176, 27–35. [Google Scholar] [CrossRef]
- Madireddi, S.; Eun, S.Y.; Lee, S.W.; Nemcovicova, I.; Mehta, A.K.; Zajonc, D.M.; Nishi, N.; Niki, T.; Hirashima, M.; Croft, M. Galectin-9 controls the therapeutic activity of 4-1BB-targeting antibodies. J. Exp. Med. 2014, 211, 1433–1448. [Google Scholar] [CrossRef]
- Nobumoto, A.; Nagahara, K.; Oomizu, S.; Katoh, S.; Nishi, N.; Takeshita, K.; Niki, T.; Tominaga, A.; Yamauchi, A.; Hirashima, M. Galectin-9 suppresses tumor metastasis by blocking adhesion to endothelium and extracellular matrices. Glycobiology 2008, 18, 735–744. [Google Scholar] [CrossRef]
- Niki, T.; Tsutsui, S.; Hirose, S.; Aradono, S.; Sugimoto, Y.; Takeshita, K.; Nishi, N.; Hirashima, M. Galectin-9 is a high affinity IgE-binding lectin with anti-allergic effect by blocking IgE-antigen complex formation. J. Biol. Chem. 2009, 284, 32344–32352. [Google Scholar] [CrossRef]
- Lv, K.; Xu, W.; Wang, C.; Niki, T.; Hirashima, M.; Xiong, S. Galectin-9 administration ameliorates CVB3 induced myocarditis by promoting the proliferation of regulatory T cells and alternatively activated Th2 cells. Clin. Immunol. 2011, 140, 92–101. [Google Scholar] [CrossRef]
- PB, J.R.; Schreiber, T.H.; Rajasagi, N.K.; Suryawanshi, A.; Mulik, S.; Veiga-Parga, T.; Niki, T.; Hirashima, M.; Podack, E.R.; Rouse, B.T. TNFRSF25 agonistic antibody and galectin-9 combination therapy controls herpes simplex virus-induced immunoinflammatory lesions. J. Virol. 2012, 86, 10606–10620. [Google Scholar] [CrossRef]
- Kadowaki, T.; Morishita, A.; Niki, T.; Hara, J.; Sato, M.; Tani, J.; Miyoshi, H.; Yoneyama, H.; Masaki, T.; Hattori, T.; et al. Galectin-9 prolongs the survival of septic mice by expanding Tim-3-expressing natural killer T cells and PDCA-1+ CD11c+ macrophages. Crit. Care 2013, 17, 284. [Google Scholar] [CrossRef]
- Sehrawat, S.; Suryawanshi, A.; Hirashima, M.; Rouse, B.T. Role of Tim-3/galectin-9 inhibitory interaction in viral-induced immunopathology: Shifting the balance toward regulators. J. Immunol. 2009, 182, 3191–3201. [Google Scholar] [CrossRef] [PubMed]
- Mengshol, J.A.; Golden-Mason, L.; Arikawa, T.; Smith, M.; Niki, T.; McWilliams, R.; Randall, J.A.; McMahan, R.; Zimmerman, M.A.; Rangachari, M.; et al. A crucial role for Kupffer cell-derived galectin-9 in regulation of T cell immunity in hepatitis C infection. PLoS ONE 2010, 5, 9504. [Google Scholar] [CrossRef]
- Wang, F.; Xu, J.; Liao, Y.; Wang, Y.; Liu, C.; Zhu, X.; Chen, Z.K.; Sun, Z. Tim-3 ligand galectin-9 reduces IL-17 level and accelerates Klebsiella pneumoniae infection. Cell Immunol. 2011, 269, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Vega-Carrascal, I.; Bergin, D.A.; McElvaney, O.J.; McCarthy, C.; Banville, N.; Pohl, K.; Hirashima, M.; Kuchroo, V.K.; Reeves, E.P.; McElvaney, N.G. Galectin-9 signaling through TIM-3 is involved in neutrophil-mediated Gram-negative bacterial killing: An effect abrogated within the cystic fibrosis lung. J. Immunol. 2014, 192, 2418–2431. [Google Scholar] [CrossRef]
- Elahi, S.; Niki, T.; Hirashima, M.; Horton, H. Galectin-9 binding to Tim-3 renders activated human CD4+ T cells less susceptible to HIV-1 infection. Blood 2012, 119, 4192–4204. [Google Scholar] [CrossRef]
- Abdel-Mohsen, M.; Chavez, L.; Tandon, R.; Chew, G.M.; Deng, X.; Danesh, A.; Keating, S.; Lanteri, M.; Samuels, M.L.; Hoh, R.; et al. Human Galectin-9 Is a Potent Mediator of HIV Transcription and Reactivation. PLoS Pathog. 2016, 12, e1005677. [Google Scholar] [CrossRef]
- Jayaraman, P.; Sada-Ovalle, I.; Beladi, S.; Anderson, A.C.; Dardalhon, V.; Hotta, C.; Kuchroo, V.K.; Behar, S.M. Tim3 binding to galectin-9 stimulates antimicrobial immunity. J. Exp. Med. 2010, 207, 2343–2354. [Google Scholar] [CrossRef] [PubMed]
- Dembele, B.P.; Chagan-Yasutan, H.; Niki, T.; Ashino, Y.; Tangpukdee, N.; Shinichi, E.; Krudsood, S.; Kano, S.; Hattori, T. Plasma levels of Galectin-9 reflect disease severity in malaria infection. Malar. J. 2016, 15, 403. [Google Scholar] [CrossRef] [PubMed]
- Chagan-Yasutan, H.; Lacuesta, T.L.; Ndhlovu, L.C.; Oguma, S.; Leano, P.S.; Telan, E.F.; Kubo, T.; Morita, K.; Uede, T.; Dimaano, E.M.; et al. Elevated levels of full-length and thrombin-cleaved osteopontin during acute dengue virus infection are associated with coagulation abnormalities. Thromb. Res. 2014, 134, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Bai, G.; Motoda, H.; Ozuru, R.; Chagan-Yasutan, H.; Hattori, T.; Matsuba, T. Synthesis of a cleaved form of osteopontin by THP-1 cells and Its alteration by phorbol 12-Myristate 13-Acetate and BCG infection. Int. J. Mol. Sci. 2018, 19, 418. [Google Scholar] [CrossRef]
- Nau, G.J.; Guilfoile, P.; Chupp, G.L.; Berman, J.S.; Kim, S.J.; Kornfeld, H.; Young, R.A. A chemoattractant cytokine associated with granulomas in tuberculosis and silicosis. Proc. Natl. Acad. Sci. USA 1997, 94, 6414–6419. [Google Scholar] [CrossRef] [PubMed]
- Hasibuan, F.M.; Shiratori, B.; Senoputra, M.A.; Chagan-Yasutan, H.; Koesoemadinata, R.C.; Apriani, L.; Takahashi, Y.; Niki, T.; Alisjahbana, B.; Hattori, T. Evaluation of matricellular proteins in systemic and local immune response to Mycobacterium tuberculosis infection. Microbiol. Immunol. 2015, 59, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Shiratori, B.; Zhao, J.; Okumura, M.; Chagan-Yasutan, H.; Yanai, H.; Mizuno, K.; Yoshiyama, T.; Idei, T.; Ashino, Y.; Nakajima, C.; et al. Immunological roles of elevated plasma levels of matricellular proteins in Japanese patients with pulmonary tuberculosis. Int. J. Mol. Sci. 2016, 18. [Google Scholar] [CrossRef]
- Niki, T.; Fujita, K.; Rosen, H.; Hirashima, M.; Masaki, T.; Hattori, T.; Hoshino, K. Plasma Galectin-9 Concentrations in Normal and Diseased Condition. Cell Physiol. Biochem. 2018, 50, 1856–1868. [Google Scholar] [CrossRef]
- Anderson, A.C.; Joller, N.; Kuchroo, V.K. Lag-3, Tim-3, and TIGIT: Co-inhibitory Receptors with Specialized Functions in Immune Regulation. Immunity 2016, 44, 989–1004. [Google Scholar] [CrossRef]
- Klement, J.D.; Paschall, A.V.; Redd, P.S.; Ibrahim, M.L.; Lu, C.; Yang, D.; Celis, E.; Abrams, S.I.; Ozato, K.; Liu, K. An osteopontin/CD44 immune checkpoint controls CD8+ T cell activation and tumor immune evasion. J. Clin. Investig. 2018, 128, 5549–5560. [Google Scholar] [CrossRef]
- Bai, G.; Matsuba, T.; Kikuchi, H.; Chagan-Yasutan, H.; Motoda, H.; Ozuru, R.; Yamada, O.; Oshima, Y.; Hattori, T. Inhibition of inflammatory-molecule synthesis in THP-1 cells stimulated with phorbol 12-myristate 13-acetate by brefelamide derivatives. Int. Immunopharmacol. 2019, 75, 105831. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yamada, O.; Kida, S.; Murase, S.; Hattori, T.; Oshima, Y.; Kikuchi, H. Downregulation of PD-L1 by amide analogues of brefelamide: Alternatives to antibody-based cancer immunotherapy. Exp. Ther. Med. Press 2020. [Google Scholar] [CrossRef]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).