Abstract
Astrocyte glycogen is a critical metabolic variable that affects hypothalamic control of glucostasis. Glucocorticoid hormones regulate peripheral glycogen, but their impact on hypothalamic glycogen is not known. A hypothalamic astrocyte primary culture model was used to investigate the premise that glucocorticoids impose sex-dimorphic independent and interactive control of glycogen metabolic enzyme protein expression and glycogen accumulation. The glucocorticoid receptor (GR) agonist dexamethasone (DEX) down-regulated glycogen synthase (GS), glycogen phosphorylase (GP)–brain type (GPbb), and GP–muscle type (GPmm) proteins in glucose-supplied male astrocytes, but enhanced these profiles in female. The catecholamine neurotransmitter norepinephrine (NE) did not alter these proteins, but amplified DEX inhibition of GS and GPbb in male or abolished GR stimulation of GPmm in female. In both sexes, DEX and NE individually increased glycogen content, but DEX attenuated the magnitude of noradrenergic stimulation. Glucoprivation suppressed GS, GPbb, and GPmm in male, but not female astrocytes, and elevated or diminished glycogen in these sexes, respectively. Glucose-deprived astrocytes exhibit GR-dependent induced glycogen accumulation in both sexes, and corresponding loss (male) or attenuation (female) of noradrenergic-dependent glycogen build-up. Current evidence for GR augmentation of hypothalamic astrocyte glycogen content in each sex, yet divergent effects on glycogen enzyme proteins infers that glucocorticoids may elicit opposite adjustments in glycogen turnover in each sex. Results document GR modulation of NE stimulation of glycogen accumulation in the presence (male and female) or absence (female) of glucose. Outcomes provide novel proof that astrocyte energy status influences the magnitude of GR and NE signal effects on glycogen mass.
1. Introduction
Adrenal glucocorticoid hormones act on classical/type II glucocorticoid receptors (GRs) expressed in discrete organs (e.g., the liver, adipose tissue, pancreas, and brain) to regulate systemic glucostasis through control of glucose production, uptake, and storage [1,2]. Glucose is stored as the complex polysaccharide glycogen in liver, skeletal muscle, and brain. Glucocorticoid regulation of hepatic glycogen metabolism, involving modulation of noradrenergic input, is well documented [3,4,5,6]. In the brain, this energy depot is maintained primarily within the astrocyte cell compartment [7,8,9]. Astrocyte glycogen affects neuro-metabolic stability as it undergoes continuous remodeling during metabolic homeostasis, and provides needed oxidizable energy substrate during states of increased neurological activity or neuro-glucopenia [10,11]. A sizable proportion of glucose taken up from the circulation into brain astrocytes passes through the glycogen shunt, wherein glucose is sequentially incorporated into and released from this polysaccharide prior to glycolytic catabolism and conversion to the trafficable metabolic fuel L-lactate [12,13]. Glycogen metabolism is monitored in the hypothalamus as an indicator of substrate fuel accumulation to shape neural outflow governing systemic glucose homeostasis [14]. While the hypothalamus is a principal target for glucose-sensitive glucocorticoid and noradrenergic signaling, insight into potential singular or interactive effects of these stimuli on local astrocyte glycogen amassment and turnover during glucose sufficiency or deprivation is presently lacking.
Glycogen metabolism is governed by opposing actions of glycogen synthase (GS) and glycogen phosphorylase (GP) enzymes, which correspondingly promote glycogen synthesis or glycogenolysis. Astrocytes express distinctive GP variants, e.g., GP–muscle (GPmm)- and GP–brain (GPbb)-type variants [15]. Evidence that GPmm and GPbb respectively mediate noradrenergic versus glucoprivic induction of astrocyte breakdown [16] implies that brain astrocyte glycogen breakdown may be subject to physiological stimulus-specific regulation. Intracerebroventricular administration of the GR agonist dexamethasone (DEX) affects male rat hypothalamic GS and GPmm protein levels in vivo [17], but it is unclear if GR ligands control hypothalamic astrocyte glycogen by direct action or through intermediary cell types. GR expression in cortical and cerebellar astrocytes is documented GR [18], but localization of GR mRNA and/or protein to hypothalamic astrocytes has not been confirmed. The catecholamine NE is a neurochemical link between hindbrain dorsal vagal complex metabolic-sensory A2 noradrenergic neurons and glucose-regulatory hypothalamic loci [19]. NE up-regulates hypothalamic total (i.e., isoform-nonspecific) tissue GP protein content during hypoglycemia [20]. NE imposes direct regulation of hypothalamic astrocyte function due to alpha1-adrenergic receptor (α1-AR), alpha2-AR (α2-AR), and beta1-AR (β1-AR) protein expression by these glia [21,22]. GR exert control of norepinephrine (NE)-initiated cortical astrocyte glycogenolysis [23], but it is unclear if glucocorticoids affect NE control of hypothalamic glycogen metabolism. Current research used a characterized astrocyte primary culture model [22,24,25] in conjunction with Western blot and uHPLC-electrospray ionization-mass spectrometry (LC-ESI-MS) methods [26] to examine the premise that GRs exert independent effects on and/or interact with NE to control glycogen metabolic enzyme protein expression and glycogen content in hypothalamic astrocytes. Hypothalamic–pituitary–adrenal glucocorticoid (HPA) axis outflow is stimulated by stress, including hypoglycemia, causing increased GR activation in the hypothalamus and other neural targets. Central aminergic autonomic transmission is also augmented in reaction to hypoglycemia, resulting in intensified noradrenergic input to the hypothalamus [27]. Effects of these glucose-sensitive regulatory cues on hypothalamic astrocyte glycogen metabolism during glucose deficiency have not been investigated. Recent studies show that glycogen mass varies significantly between male and female astrocytes, and that glucoprivation in vivo or in vitro causes sex-specific changes in volume of this metabolic fuel reserve [25,28,29]. These findings, in alignment with current U.S. National Institutes of Health policy emphasis on consideration of sex as a critical biological variable, provide strong justification for determination here of whether the direction and/or magnitude of GR regulation of hypothalamic astrocyte glycogen metabolism may vary according to sex depending upon glucose availability.
2. Materials and Methods
2.1. Primary Astrocyte Cell Cultures
High-purity hypothalamic astrocyte primary cultures were prepared from adult (2–3 months of age) male or female rat brains, as previously described [22,24,25]. All animal protocols were performed in accordance with the NIH Guide for the Care and Use of Laboratory Animals, 8th Edition, under ULM Institutional Animal Care and Use Committee approval. Briefly, dissected whole-hypothalamus tissue blocks were enzymatically digested in 2.5 mL 2.5% trypsin (prod. no. 15090-046; Thermo Fisher Scientific, Waltham, MA, USA) and 22.5 mL Hanks’ balanced salt solution (HBSS; prod. no. H2387, Sigma-Aldrich, St. Louis, MO, USA), incubated (30 min) in a 37 °C water bath with shaking at 10 min intervals, then centrifuged (5 min) at 300× g. Tissue was dissociated by pipet into a single-cell suspension in astrocyte plating media (DMEM high-glucose media (prod. no. 12800-017; ThermoFisherSci., Waltham, MA, USA) supplemented with 10.0% heat-inactivated fetal bovine serum (FBS; GE Healthcare Bio-Sciences, Pittsburgh, PA, USA) and 1.0% penicillin/streptomycin (prod. no. 15140-122, ThermoFisherSci.). Dissociated cells were suspended in adjusted 20 mL media, then plated at an initial concentration of 50 μg/mL in poly D-lysine (prod. no. A-003-E, MilliporeSigma, Burlington, MA, USA)-coated T75 culture flasks. Approximately two weeks after plating, microglia were removed from cultures by aspiration after shaking at 180 rpm (30 min). Cells were re-suspended in fresh media (20 mL) and shaken first on an orbital platform at 240 rpm (6–8 h), then vigorously by hand (1 min) before aspiration to discard oligodendrocyte precursor cells, as described [22]. Confluent astrocytes were washed twice with 10 mM phosphate-buffered saline, pH 7.4, then incubated at 37 °C (7–10 min) with 5.0 mL trypsin-EDTA (prod. no. 25300-062, ThermoFisherSci.). Detached astrocytes were centrifuged at 180× g (5 min) and re-suspended in 40 mL fresh media. Purified astrocytes were then counted, plated, and grown for 12–14 days, with media changes every 2–3 days. Three days before experimentation, cells were plated at a density of 1 × 106 cells/100 mm2 in poly D-lysine–coated culture dishes. Routine Western blot and immunofluorescence cytochemical detection of the astrocyte marker protein glial fibrillary acidic protein (GFAP) showed that culture purity exceeded 95%, as reported [22,24,25].
2.2. Experimental Design
Astrocyte cultures achieving 70% confluency were incubated for 18 h in DMEM high-glucose media containing 5.0% charcoal-stripped FBS (prod. no. 12676029; ThermoFisherSci.). Astrocytes from each sex were next incubated for 8 h in 10 nM 17β-estradiol -supplemented HBSS media containing 5.5 mM (G5.5) or 0 mM (G0) glucose [30,31,32,33], as reported [25]. Glucose-supplied and glucose-deprived cultures were treated with NE (10 nM [22]; prod. no. A7257; Sigma-Aldrich), dexamethasone (100 nM [23]; prod. no. D4902; Sigma-Aldrich), or NE (10 nM) plus DEX (100 nM) dissolved in dimethyl sulfoxide (DMSO). DMSO was administered in an equal volume to all treatment groups to achieve a uniform media concentration of 0.01%. Controls were incubated with HBSS media containing DMSO alone. Astrocytes were detached from plates with 0.05% trypsin EDTA (prod. no. 25300-062; ThermoFisherSci.), washed in PBS, pelleted in lysis buffer (2.0% sodium dodecyl sulfate, 10.0% glycerol, 5% β-mercaptoethanol, 1 mM sodium orthovanadate (protease inhibitor), 60 mM Tris-HCl, pH 6.8), heat-denatured (10 min, 95 °C), or ultra-pure water for Western blot protein or LC-ESI-MS glycogen analyses, respectively.
2.3. Western Blot Analysis
Astrocyte cell pellets were heat-denatured (10 min, 95 °C), sonicated, centrifuged, and diluted with 2x Laemmli buffer. Sample protein concentrations were determined by NanoDrop spectrophotometry (prod. no. ND-ONE-W, ThermoFisherSci.). Pooled sample aliquots of equivalent protein mass were loaded for each treatment group for separation in Bio-Rad Stain Free 10% acrylamide gels (prod. no 161-0183, Bio-Rad, Hercules, CA, USA), as described [34,35]. After UV gel activation (1 min) in a Bio-Rad ChemiDoc™ Touch Imaging System, proteins were transblotted to 0.45-µm PVDF-Plus membranes (prod. no. 1212639; Data Support Co., Panorama City, CA, USA). Membrane buffer washes and antibody incubations were performed by Freedom Rocker™ Blotbot® automation (Next Advance, Inc., Troy, NY, USA). After blocking (2 h) with Tris-buffered saline (TBS), pH 7.4, containing 0.2 % Tween-20 (prod. no. 9005-64-5; VWR, Radnor, PA, USA) and 2% bovine serum albumin (BSA) (prod. no. 9048-46-8; VWR), membranes were incubated (24–48 h; 4 °C) with primary antisera raised in rabbit against GS (prod. no. 3893S; 1: 2000; Cell Signaling Technology, Danvers, MA, USA), GPbb (prod. no. NBP1-32799; 1:2000; Novus Biologicals, Littleton, CO, USA), or GPmm (prod. no. NBP2-16689; 1: 2000; Novus Biol.) [34]. Membranes were next incubated (1 h) with goat anti-rabbit horseradish peroxidase-conjugated secondary antibodies (prod. no. NEF812001EA; 1:4000; PerkinElmer, Waltham, MA, USA), followed by Supersignal WestFemto maximum sensitivity chemiluminescent substrate (prod. no. 34096; ThermoFisherSci.). Target protein optical density (O.D.) signals were quantified in the ChemiDoc™ Touch Imaging System described above, and normalized to total in-lane protein, e.g., all protein electrophoresed in the individual sample lane, using Bio-Rad proprietary stain-free imaging gel technology and Image Lab™ 6.0.0 software (http://www.bio-rad.com/en-us/applications-technologies/stain-free-imaging-technology?ID=NZ0G1815) (accessed on 29 September 2022), as previously reported [21,22,24,34,35]. This superior method for Western blot normalization markedly reduces data variability through improved measurement accuracy and precision [36,37]. Precision plus protein molecular weight dual color standards (prod. no. 161-0374, Bio-Rad) were included in each Western blot analysis in the current study.
2.4. Astrocyte LC-ESI-MS Glycogen Analysis
Astrocyte glycogen concentrations were measured as reported [22,24,25]. In brief, cell lysate supernatant aliquots (10 μL) were hydrolyzed by incubation (2 h) with 10 μL 0.5 mg/mL amyloglucosidase and 10 μL 0.1 M sodium acetate, pH 5.0, then sequentially heated to 100 °C (5 min), then cooled to room temperature. Glycogen was measured in a ThermoFisherSci. Vanquish UHPLC + System equipped with ThermoScientific™ Dionex™ Chromeleon™ 7 Chromatography Data System software [26]. Column and autosampler temperatures were maintained at 35 °C and 15 °C, respectively. The auto-sampler needle was washed (10 s) with 10% (v/v) methanol. Hydrolyzed and non-hydrolyzed samples were derivatized with 100 μL 0.5 M 1-phenyl-3-methyl-5-pyrazolone (PMP) reagent supplemented with 0.3 M NaOH, then acidified with 400 μL 0.75% formic acid and extracted with chloroform to remove excess PMP. Sample supernatant (400 μL) was vacuum-concentrated to remove organic solvents, frozen at −80 °C, and lyophilized. Lyophilization product was diluted to 1.0 mL with 10 mM ammonium acetate, bath-sonicated (30 s), and centrifuged. Supernatant aliquots (250 μL) were transferred to 350 μL inserts, which were placed in 2 mL Surestop vials in an autosampler tray. D-(+)-Glucose-PMP was resolved using a Shodex™ Asahipak™ NH2P-40 3E column with acetonitrile:10 mM ammonium acetate (75:25 v/v; 0.2 mL/min) as mobile phase. D-(+)-Glucose-PMP ion chromatograms were extracted from Total Ion Current (TIC) at m/z 510.2 to generate area-under-the curve data. Critical LC-ESI-MS parameters such as sheath gas pressure (SGP; 25 psig), auxiliary gas pressure (AGP; 4.6 psig), sweep gas pressure (SWGP; 0.5 psig), vaporizer temperature (VT; 150 °C), ion transfer tube temperature (ITT; 150 °C), source voltage (−2000 V), foreline pressure (1.76 Torr; auto-set by instrument- and variable), source gas (nitrogen; Genius NM32LA 110V, 10–6520; Peak Scientific, Inchinnan, UK), and mass peak area detection algorithm (ICIS/Genesis) were maintained at pre-determined optimum settings. Sample protein content was determined by NanoDrop spectrophotometry (prod. no. ND-ONE-W, ThermoFisherSci.). Sample glycogen concentrations were expressed as microgram per milligram protein.
2.5. Statistics
Mean normalized protein O.D. and glycogen values were evaluated between treatment groups within each sex by two-way analysis of variance and Student Newman Keuls post hoc test. Differences of p < 0.05 were considered significant. In each figure, statistical differences between specific pairs of treatment groups are denoted as follows: * p < 0.05; ** p < 0.01; *** p < 0.001.
3. Results
Figure 1 depicts patterns of GS protein expression in male (Figure 1A; F(7,16) = 23.98, p < 0.001; glucose status main effect: F(1,16) = 51.65, n = 3, p < 0.001; treatment main effect: F(3,16) = 35.99, p < 0.001; glucose status/treatment interaction: F(3,16) = 2.75, p = 0.077) and female (Figure 1B; F(7,16) = 22.41, n = 3, p < 0.001; glucose status main effect: F(1,16) = 19.23, p < 0.001; treatment main effect: F(3,16) = 44.57, p < 0.001; glucose status/treatment interaction: F(3,16) = 1.22, p = 0.335) hypothalamic astrocytes incubated with the GR agonist dexamethasone (DEX), the adrenergic receptor ligand norepinephrine (NE), or DEX plus NE. For each sex, treatment groups including astrocytes supplied with 5.5 mM glucose (G5.5), at left, are represented by gray bars, while glucose-deprived (G0) treatment groups, at right, are presented in white. Data show that DEX significantly decreased or increased astrocyte GS protein content in male (Figure 1A) or female (Figure 1B), respectively, compared to vehicle controls (G5.5-DEX (horizontal-striped gray bar) versus G5.5-V (solid gray bar)). NE administration to glucose-supplied cells did not modify GS expression in either sex (G5.5-NE (diagonal-striped gray bar) versus G5.5-V). Male astrocytes treated with DEX plus NE showed a significant decline in GS protein relative to V, DEX, and NE treatment groups (G5.5-DEX/NE (cross-hatched gray bar) versus G5.5-V, G5.5-DEX, or G5.5-NE). Incubation of female astrocytes with DEX plus NE elevated GS content above the control range (G5.5-DEX/NE versus G5.5-V) to levels associated with the GR treatment group (G5.5-DEX/NE versus G5.5-DEX).
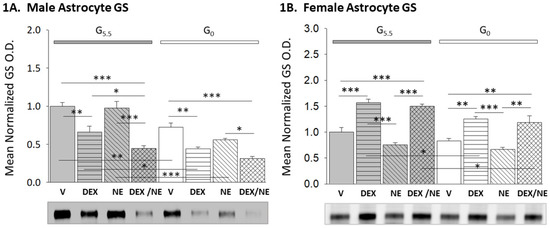
Figure 1.
Effects of the glucocorticoid receptor (GR) agonist dexamethasone (DEX) alone or in combination with the catecholamine transmitter norepinephrine (NE) on glycogen synthase (GS) protein expression in glucose-supplied or glucose-deprived male versus female hypothalamic primary astrocyte cultures. Hypothalamic astrocyte cultures derived from adult rats of each sex were pre-incubated in steroid-free media (18 h) prior to incubation (8 h) with HBSS media supplemented with 5.5 (G5.5) or 0 (G0) mM glucose containing DEX (10 nM), NE (100 nM), DEX (10 nM) plus NE (100 nM), or vehicle only (V; DMSO). Astrocyte lysate aliquots were analyzed by stain-free Western blot for GS protein content. Target protein optical density (O.D.) measures were obtained in a ChemiDoc™ Touch Imaging System, and normalized to total in-lane protein, e.g., all protein electrophoresed in the individual sample lane using Bio-Rad Image Lab™ 6.0.0 software. Data mean normalized GS protein O.D. measures ± S.E.M. for male ((1A); n = 3 individual experiments per treatment group) and female ((1B); n = 3 individual experiments per treatment group) G5.5-(gray bars) or G0-(white bars) exposed astrocytes divided into the following treatment groups: V (solid bars), DEX (horizontal-striped bars), NE (diagonal-striped bars), DEX/NE (cross-hatched bars). Mean normalized O.D. data were analyzed by two-way ANOVA and Student-Neuman-Keuls post hoc test. Statistical differences between discrete pairs of treatment groups are denoted as follows: * p < 0.05; ** p < 0.01; *** p < 0.001.
Glucose deprivation decreased GS levels in male astrocytes (G0-V (solid white bar) versus G5.5-V), but did not modify this protein profile in the female. DEX administration to G0 male and female cells caused diminution or augmentation of GS expression, respectively, compared to G0 controls (G0-DEX versus G0-V). In each sex, GS protein significantly lower in G0-DEX (horizontal-striped white bar) versus G5.5-DEX (horizontal-striped gray bar) groups. Male and female astrocytes incubated with NE alone exhibited no change in GS content, whereas DEX plus NE treatment suppressed (male) or increased (female) this protein profile compared to G0-V and G0-NE groups.
Data in Figure 2 describe effects of DEX or NE treatment alone versus combinatory NE and DEX administration on GPbb protein expression in male (Figure 2A; F(7,16) = 30.39, n = 3, p < 0.001; glucose status main effect: F(1,16) = 40.50, p < 0.001; treatment main effect: F(3,16) = 50.33, p < 0.001; glucose status/treatment interaction: F(3,16) = 7.07, p = 0.003) and female (Figure 1B; F(7,16) = 20.11, n = 4, p < 0.001; glucose status main effect: F(1,16) = 11.44, p = 0.002; treatment main effect: F(3,16) = 42.46, p < 0.001; glucose status/treatment interaction: F(3,16) = 0.64, p = 0.598) hypothalamic astrocytes. Results reveal opposite effects of DEX on GPbb protein levels in G5.5 male (decreased) and female (increased) astrocytes, but disclose no effect of NE alone on this protein profile in either sex. Male G5.5 astrocytes incubated with DEX plus NE exhibited significantly lower GPbb levels compared to G5.5-V, G5.5-DEX, and G5.5-NE groups, whereas in the female, DEX plus NE administration elevated GPbb expression relative to G5.5-V controls. Glucoprivation diminished GPbb protein in male, but not female astrocytes. DEX respectively decreased or stimulated GPbb expression in G0 male and female cultures. G0 astrocytes incubated with DEX plus NE showed reductions (male) or augmentation (female) of GPbb levels compared to G0-V and GO-NE groups.
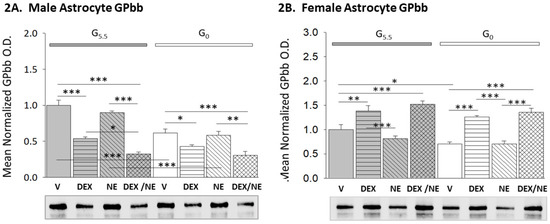
Figure 2.
Impact of glucose provision versus starvation on DEX- or NE-associated patterns of glycogen phosphorylase–brain type (GPbb) expression in male and female hypothalamic primary astrocyte cultures. Data illustrate mean normalized GPbb protein O.D. values ± S.E.M. for glucose-suppled (G5.5; gray bars) or glucose-deprived (G0; white bars) male ((2A); n = 3 independent experiments per treatment group) and female ((2B); n = 4 independent experiments per treatment group) hypothalamic astrocytes assigned to the following treatment groups: G5.5- or G0-V (solid bars), G5.5- or G0-DEX (horizontal-striped bars), G5.5- or G0-NE (diagonal-striped bars), G5.5- or G0-DEX/NE (cross-hatched bars). * p < 0.05; ** p < 0.01; *** p < 0.001.
Figure 3 depicts patterns of GPmm protein expression in male (Figure 3A; F(7,16) = 16.45, n = 3, p < 0.001; glucose status main effect: F(1,16) = 12.17, p = 0.003; treatment main effect: F(3,16) = 30.97, p < 0.001; glucose status/treatment interaction: F(3,16) = 3.36, p = 0.045) and female (Figure 3B; F(7,16) = 7.81, n = 3, p < 0.001; glucose status main effect: F(1,16) = 0.71, p = 0.413; treatment main effect: F(3,16) = 17.30, p < 0.001; glucose status/treatment interaction: F(3,16) =0.68, p = 0.578) hypothalamic astrocytes incubated with DEX, NE, or DEX plus NE. Data indicate that DEX suppressed or elevated GPmm protein in male versus female G5.5 astrocytes, in that order. In each sex, incubation with DEX alone or DEX plus NE elicited adjustments (decreased in male; increased in female) of comparable magnitude in GPmm expression. GPmm content was significantly lower in G0 versus G5.5 male, but not female astrocytes. G0 male astrocytes exhibited a reduction in GPmm protein content after incubation with DEX plus NE, but not DEX alone, whereas G0 female astrocytes showed increases in GPmm levels after incubation with either DEX or DEX plus NE.
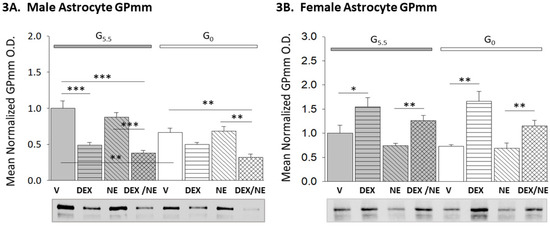
Figure 3.
Effects of DEX or NE alone or in combination on glycogen phosphorylase–muscle type (GPmm) expression in male versus female hypothalamic primary astrocytes. Bars depict mean normalized male ((3A); n = 3 independent experiments per treatment group) and female ((3B); n = 3 independent experiments per treatment group) hypothalamic astrocyte GPmm protein O.D. values ± S.E.M. after the following treatments: G5.5- or G0-V, G5.5- or G0-DEX, G5.5- or G0-NE, G5.5- or G0-DEX/NE. * p < 0.05; ** p < 0.01; *** p < 0.001.
The data in Figure 4 illustrate effects of DEX or NE alone or combinatory DEX plus NE treatment on male (Figure 4A; F(7,16) = 29.35, n = 3, p < 0.001; glucose status main effect: F(1,16) = 14.33, p = 0.002; treatment main effect: F(3,16) = 25.84, p < 0.001; glucose status/treatment interaction: F(3,16) = 37.86, p < 0.001) and female (Figure 4B; F(7,16) = 42.52, n = 3, p < 0.001; glucose status main effect: F(1,16) = 24.53, p < 0.001; treatment main effect: F(3,16) = 52.81, p < 0.001; glucose status/treatment interaction: F(3,16) = 38.22, p < 0.001) hypothalamic astrocyte glycogen content. As shown in Figure 4A and Figure 4B, respectively, glycogen levels in G5.5 male and female astrocytes were significantly increased by DEX treatment. In each sex, incubation with NE alone markedly elevated glycogen levels beyond measures associated with G5.5-V and G5.5-DEX groups. Administration of DEX plus NE to G5.5 male or female astrocytes also augmented glycogen mass, but the magnitude of this increase was less than that associated with DEX alone. Glucoprivation caused opposing adjustments in male (increased) versus female (decreased) astrocyte glycogen content. DEX treatment enhanced glycogen content of G0 male and female astrocytes. In each sex, glycogen levels were significantly higher in G0-DEX versus G5.5-DEX groups. Glucose withdrawal affected noradrenergic regulation of glycogen mass, as NE had no (male) or diminished (female) impact on GO astrocyte glycogen content. G0 male astrocyte glycogen content was unaffected by DEX plus NE treatment, whereas this combinatory therapy increased G0 female astrocyte glycogen mass beyond that elicited by NE alone.
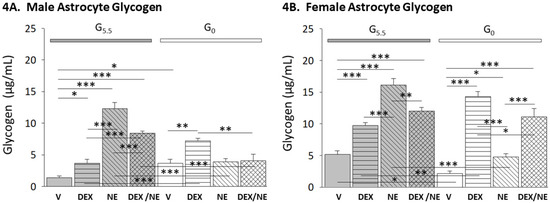
Figure 4.
Impact of glucose on DEX or NE regulation of male versus female hypothalamic astrocyte glycogen concentrations. Hypothalamic astrocyte lysate glycogen content was measured by ultra-high performance liquid chromatography-electrospray ionization mass spectrometry. Data portray mean astrocyte glycogen concentrations (μg/mg protein) ± S.E.M. for glucose-suppled (G5.5) or glucose-deprived (G0) male ((4A); n = 3 independent experiments per treatment group) and female ((4B); n = 3 independent experiments per treatment group) astrocytes treated as follows: G5.5- or G0-V, G5.5- or G0-DEX, G5.5- or G0-NE, G5.5- or G0-DEX/NE. * p < 0.05; ** p < 0.01; *** p < 0.001.
Figure 5 illustrates GR expression profiles in male (Figure 5A; F(7,16) = 17.04, n = 3, p < 0.001; glucose status main effect: F(1,16) = 38.47, p < 0.001; treatment main effect: F(3,16) = 25.59, p < 0.001; glucose status/treatment interaction: F(3,16) = 1.35, p = 0.295) and female (Figure 5B; F(7,16) = 5.14, n = 4, p = 0.001; glucose status main effect: F(1,16) = 6.70, p = 0.016; treatment main effect: F(3,16) = 5.44, p = 0.005; glucose status/treatment interaction: F(3,16) = 4.31, p = 0.014) hypothalamic astrocytes incubated with DEX, NE, or DEX plus NE. Data show that GR protein decreased in G5.5 male astrocytes after incubation with DEX, NE, or DEX plus NE, and that the magnitude of this reduction was greatest in response to the latter combinatory dosing. Meanwhile, GR expression in female G5.5 astrocytes was unaffected by any of these treatments. Glucoprivation did not modify GR expression in astrocytes of either sex. Treatment with either DEX or DEX plus NE resulted in diminished GR protein profiles in G0 male and female astrocytes.
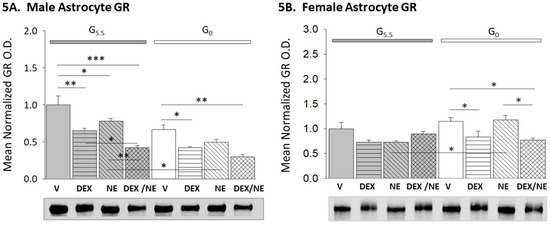
Figure 5.
Effects of glucose on DEX- or NE-associated patterns of male versus female hypothalamic astrocyte GR expression. Bars depict mean normalized male ((5A); n = 3 independent experiments per treatment group) and female ((5B); n = 4 independent experiments per treatment group) hypothalamic astrocyte GR protein O.D. values ± S.E.M. after the following treatments: G5.5- or G0-V, G5.5- or G0-DEX, G5.5- or G0-NE, G5.5- or G0-DEX/NE. * p < 0.05; ** p < 0.01; *** p < 0.001.
4. Discussion
The current research used primary hypothalamic astrocyte cultures from each sex to address the hypothesis that GR impose sex-dimorphic regulation of glycogen metabolism in these glial cells. The results show that the GR agonist DEX causes divergent, sex-contingent alterations in astrocyte GS, GPbb, and GPmm protein levels, yet increases glycogen mass in each sex. Outcomes also document GR and NE interaction as DEX blunts noradrenergic up-regulation of glycogen content in the presence (male and female) or absence (female) of glucose. Novel evidence that glucoprivation amplifies DEX-induced hypothalamic astrocyte glycogen accumulation in each sex, but abolishes (male) or attenuates (female) noradrenergic augmentation of glycogen supports the view that cellular metabolic status impacts GR and NE regulation of glycogen mass.
The present outcomes show that astrocyte GR activation up- (female) or down- (male) regulates GS and GP variant protein profiles, signifying that positive versus negative directionality of post-receptor signaling effects elicited by the current, well-characterized DEX dosage is sex-specific. Bi-directional GR regulation of these target proteins may reflect differences in volume of DEX input to male versus female astrocytes due to disparate GR expression levels and/or differential modulation of downstream post-receptor signaling, transcriptional, and/or post-translational events in male versus female astrocytes. The former possibility is supported by observations that at exposure levels implemented here, DEX down-regulates GR protein expression in male, but not female. Further effort will be required to examine for each sex whether graded DEX administration in vitro promotes dosage-proportionate changes in astrocyte GS, GPbb, and/or GPmm protein expression over all or part of a comprehensive, physiological-like dose range, and to establish, moreover, whether stimulatory versus inhibitory protein responses to DEX are concentration-dependent. Current outcomes reveal discrepant levels of measurable astrocyte glycogen between the two sexes in non-drug-treated control cultures, a divergence that may reflect, in part, augmented GS and GPbb protein expression in male versus female alongside comparatively higher GPmm content in the latter sex [38]. Notably, despite present observations of sex-specific DEX effects on glycogenic enzyme protein expression, DEX was found here to enhance glycogen levels in astrocytes of each sex, which reflects an evident positive effect on the ratio of glycogen synthesis versus breakdown. Current evidence for DEX-induced diminution of male astrocyte GS and GP isoform protein profiles does not disclose whether specific activities of these enzymes are modified by that treatment. GS is active in the non-phosphorylated state and is allosterically activated by glucose 6-phosphate, whereas GP is activated by phosphorylation or AMP allosteric effects; it remains unclear if or how glucocorticoids may regulate these post-transcriptional modifications in either sex. It would be informative to learn if DEX exerts sex-specific effects on expression of activated GS, GPbb, and GPmm relative to total astrocyte glycogenic enzyme protein profiles, as that information could shed light on glucocorticoid control of glycogen turnover aside from mass. The question of whether astrocyte glycogen in its entirety is a common substrate for GPbb- versus GPmm-mediated breakdown, or alternatively, if this fuel reserve is organized into spatially distinct pools that are disassembled by a single GP variant has not yet been addressed. In the present context, it would be useful to discover whether DEX elicits similar or disparate changes in GPbb versus GPmm enzyme activity, and how DEX-mediated adjustments in individual GP isoform specific activity may impact net glycogen accrual. However, it needs to be pointed out that efforts to resolve this critical issue are currently impeded by the lack of available antibody-based analytical tools for quantification of GP variant phosphorylation status. There exists a need to identify potential signaling pathways that may be involved in sex-specific DEX effects on hypothalamic astrocyte glycogen accumulation, including whether protein kinase A-glycogen phosphorylase kinase-GP dysfunction occur alone or simultaneous to glycogen synthase kinase-3β activation [39]. Recent studies show that stress amplifies cortical glycogen amassment by mechanisms involving GPbb inactivation [40]; further research is required to determine if GRs promote glycogen build-up in a brain region- and stressor-specific manner.
Current data show that glucoprivation amplifies DEX enhancement of hypothalamic astrocyte glycogen buildup. In each sex, DEX triggered a greater absolute gain in glycogen mass in glucose-deprived versus glucose-supplied astrocytes, despite causing relatively lower GS protein expression profiles in the former treatment group relative to the latter. These data infer that glucose withdrawal may directly amplify or, alternatively, may synergize with signaling mechanisms that underlie DEX stimulation of GS and/or repression of GP activity. It is plausible that detection of glucoprivic stress by astrocyte glucose sensors, namely GLUT2 and glucokinase [41,42,43], may likely exacerbate effects of GR activation on glycogen accretion in hypothalamic astrocytes. Our collective work [22,24,25] shows that glucose deficiency results in sex-specific divergent alterations in hypothalamic primary astrocyte culture glycogen accumulation in vitro. Similarly, insulin-induced hypoglycemia (IIH) causes opposing changes in hypothalamic glycogen content in vivo, as tissue levels are elevated or diminished in hypoglycemic male and female rats, respectively [28,29,38]. Hypoglycemic hypercorticosteronemia is a critical element of the integrated array of physiological counter-regulatory responses to IIH, but it should be noted that the magnitude of stress-induced adrenal glucocorticoid secretion as well as de novo brain synthesis of neurosteroid GR ligands can vary by sex [44]. Additional work is warranted to investigate whether hypoglycemic patterns of hypothalamic astrocyte GR activation impose contrary, e.g., exacerbating (male) versus attenuating (female) modulatory effects on glycogen responses to neuro-glucopenia in vivo due to differential stress-associated GR ligand signal volume. Alterations in brain tissue glucocorticoid concentrations in vivo due in part to fluctuations in adrenal glucocorticoid secretion associated with the circadian rhythm or physical, emotional, or metabolic stress will affect hormone signal volume detected by astrocytes. Ongoing work aims to establish and compare endogenous hypothalamic glucocorticoid concentrations between the two sexes under non-stress versus stress, including IIH conditions.
Our previous studies showed that noradrenergic stimulation of glycogen amassment in hypothalamic astrocyte cultures is dosage-dependent [22]. Prior reports that NE promotes glycogenolysis in cortical astrocytes [45,46] suggest that effects of this neurochemical on glycogen metabolism are likely distinctive between brain regions. Data here affirm significant augmentation of glycogen content in each sex after incubation with 10 nM NE, but reveal that this increase in mass occurs despite no change in GS, GPbb, or GPmm protein expression under current experimental conditions. Thus, at this specific exposure level, NE is evidently capable of regulating the activity of one or both glycogenic enzymes to favor net glycogen synthesis, without modifying net cellular levels of GS or either GP isoform. However, the prospect cannot be overlooked that for one or both enzymes, NE may impose here distinctive regulatory effects on transcriptional, translational, and post-translational metabolism to alter individual rates, but not ratio of these processes, resulting in no adjustment in protein content. A critical difference between prior and present work involving NE regulation of glycogenic enzyme protein expression and glycogen accumulation is that hormone treatments here were delivered to astrocytes incubated in media supplemented with 10 nM estradiol [24,25]. Hence, observed discrepancies in NE effects on glycogenic enzyme protein profiles expression may thus reflect, in part, a critical modulatory role for estradiol in noradrenergic regulation of hypothalamic glycogen metabolism. Outcomes here document sex-specific effects of glucose availability on noradrenergic control of hypothalamic glycogen as glucoprivation eradicated or blunted NE-induced augmentation of glycogen amassment in male and female astrocytes, respectively. Loss of this response in glucose-deprived male astrocytes may reflect, in part, diminished glycogen synthesis as these cells exhibit a reduction in GS protein content after NE dosing. Recent evidence that α1-AR, α2-AR, and β1-AR proteins are expressed at higher levels in glucose-supplied male versus female astrocyte cultures [47] infers that sex differences in effects of NE alone or in combination with DEX on specific target proteins may reflect, in part, dissimilar receptivity to NE. Sex-specific outcomes of NE plus DEX treatment when glucose is present may also involve divergent effects of NE on GR protein expression. Current data do not identify mechanism(s) responsible for glucoprivic-associated gain of noradrenergic suppression of GS profiles in male, but prior observations that glucose withdrawal up-regulates α2-AR and β1-AR protein profiles in that sex [38] support the possibility that augmented sensitivity to NE may be a factor.
Although DEX amplifies hypothalamic astrocyte glycogen content in the absence of NE, this steroid was also observed to paradoxically mitigate noradrenergic augmentation of this energy reserve. These findings align with evidence that DEX inhibits noradrenergic stimulation of cortical astrocyte GS activity and glycogen accumulation [14]. Here, treatment of male astrocytes with either DEX alone or DEX plus NE caused reductions in GS, GPbb, and GPmm protein profiles relative to vehicle controls, yet GS and GPmm levels were comparatively lower after DEX plus NE versus DEX incubation. The presence of significantly greater glycogen content in DEX plus NE- versus DEX-treated male astrocytes exposes apparent dissimilar effects of singular DEX versus combinatory DEX and NE treatments on glycogenic enzyme activity and regulation of glycogen amassment. However, female astrocytes incubated with DEX plus NE or DEX alone showed similar up-regulated patterns of GS and GPbb protein, lack of change in GPmm expression, and equivalent measures of glycogen mass. These dissimilar sex-contingent effects of GR and noradrenergic stimulus interaction on hypothalamic astrocyte glycogen metabolism may reflect, in part, differences in the ratio of GR and adrenergic receptor expression and relative magnitude of signal volume and/or disparate sensitivity of one or both post-receptor signal pathways to cross-regulation. Current outcomes show that glucoprivation abolishes augmentation of male hypothalamic astrocyte glycogen content by DEX plus NE treatment, but exacerbates treatment stimulation of glycogen mass in the female. Primarily similar DEX plus NE effects on GS, GPbb, and GPmm protein profiles in glucose-supplied versus glucose-deprived astrocytes infer that in the male, glucoprivic regulation of enzyme activity(-ies) may prevent glycogen buildup or alternatively render rates of glycogen synthesis and breakdown equal, but in the female may enhance glycogen amassment relative to disassembly.
The current research makes progress toward the identification of endocrine (glucocorticoid) and neurotransmitter (NE) cues that may govern hypothalamic astrocyte glycogen mass and turnover in vivo, and advances insight on how these signals may interact to affect such control. Hypothalamic glycogen status regulates neural outflow controlling glucose homeostasis [14]. The observations here of sex-dimorphic effects of these glucose-sensitive physiological signals on hypothalamic astrocyte glycogen metabolism justify further efforts to clarify astrocyte-dependent mechanisms that contribute to sex differences in counter-regulatory responses to hypoglycemia in vivo. Potential contributing factors include sex-dimorphic GR and/or AR protein expression patterns as well as sex-dimorphic post-receptor signal responses to discrete ligand concentrations. Further consideration should be given to the possibility of dissimilar astrocyte glucose sensor function between sexes, resulting in divergent cell sensitivity to DEX or NE during glucose withdrawal. The novel outcomes described here provide a firm foundation for future whole-animal model research aimed at investigation of the premise that hypothalamic astrocytes are a likely target for glucocorticoid and/or noradrenergic input to neural circuitries that govern counter-regulation, and that hormonal and/or neurochemical control of regulatory signals from astrocytes to neurons concerning glycogen metabolism and net mass may be sex-contingent during glucose abundance and/or scarcity. Sex-dimorphic changes in glycogen accumulation due to GR or AR stimulation are a likely indication of coincident adjustments in glucose entering the glycolytic pathway from the glycogen shunt. As lactate is a critical metabolic cue affecting hypothalamic nerve cell glucose-regulatory transmission, it is presumed that glucocorticoid and noradrenergic signaling may thus impose differential control of neural outflow governing glucose homeostasis in each sex by sex-specific regulation of lactate volume transferred from astrocytes to neurons.
In summary, the present project provides novel proof in support of GR involvement in hypothalamic astrocyte glycogen metabolism. GR activation elicits divergent, sex-specific changes in glycogenic enzyme protein profiles in hypothalamic astrocyte primary cultures, yet increases glycogen mass in each sex. Current data affirm noradrenergic stimulation of glycogen accumulation, but also reveal that this effect on hypothalamic astrocytes is attenuated by GR. Glucoprivic effects on glycogenic enzyme protein profiles and glycogen content vary according to sex, but this metabolic stress respectively amplifies or diminishes GR and NE stimulation of glycogen buildup in hypothalamic astrocytes of each sex. Further studies are warranted to characterize mechanisms that mediate effects of sex and glucose nutrient status on GR control of astrocyte glycogen metabolism, and to identify the molecular basis of GR and NE signal interaction.
Author Contributions
P.R.N., A.A., K.B.,P.W.S. and K.P.B. conceived of and designed the work. P.R.N., A.A., K.B. and K.P.B. acquired and analyzed data and drafted the work. P.R.N., A.A., P.W.S. and K.P.B. interpreted data and reviewed and revised the work for intellectual content. P.R.N., A.A., K.B., P.W.S. and K.P.B. approved the final version of the submitted article and are accountable for all aspects of the work. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by NIH grant DK 109382.
Institutional Review Board Statement
The studies performed here were approved by the University of Louisiana Monroe Institutional Animal Care and Use Committee, reference no. 19AUG-KPB-01, in accordance with the National Institutes of Health (NIH) Guide for Care and Use of Laboratory Animals, 8th Edition.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors have no conflict interest to declare.
Abbreviations
| DEX | dexamethasone |
| GPbb | glycogen phosphorylase–brain type |
| GPmm | glycogen phosphorylase–muscle type |
| GS | glycogen synthase |
| IIH | insulin-induced hypoglycemia |
| LC-ESI-MS | uHPLC-electrospray ionization-mass spectrometry |
| NE | norepinephrine |
References
- So, A.Y.L.; Bernal, T.U.; Pillsbury, M.L.; Yamamoto, K.R.; Feldman, B.J. Glucocorticoid regulation of the circadian clock modulates glucose homeostasis. Proc. Natl. Acad. Sci. USA 2009, 106, 17582–17587. [Google Scholar] [CrossRef] [PubMed]
- Kuo, T.; McQueen, A.; Chen, T.C.; Wang, J.C. Regulation of Glucose Homeostasis by Glucocorticoids. Glucocorticoid Signal. 2015, 872, 99–126. [Google Scholar]
- Stalmans, W.; Laloux, M. Glucocorticoids and hepatic glycogen metabolism. Monogr. Endocrinol. 1979, 12, 517–533. [Google Scholar] [CrossRef] [PubMed]
- Green, G.A.; Chenoweth, M.; Dunn, A. Adrenal glucocorticoid permissive regulation of muscle glycogenolysis: Action on protein phosphatase(s) and its inhibitor(s). Proc. Natl. Acad. Sci. USA 1980, 77, 5711–5715. [Google Scholar] [CrossRef]
- Margolis, R.N.; Curnow, R.T. The role of insulin and glucocorticoids in the regulation of hepatic glycogen metabolism: Effect of fasting, refeeding, and adrenalectomy. Endocrinology 1983, 113, 2113–2119. [Google Scholar] [CrossRef]
- Viskupic, E.; Németh, S. Are endogenous glucocorticoids involved in short-term regulation of hepatic glycogen metabolism? Endocrinol. Exp. 1985, 19, 283–289. [Google Scholar]
- Brown, A.M. Brain glycogen re-awakened. J. Neurochem. 2004, 89, 537–552. [Google Scholar] [CrossRef]
- Obel, L.F.; Müller, M.S.; Walls, A.B.; Sickmann, M.; Bak, L.K.; Waagepetersen, S.; Schousboe, A. Brain glycogen-new perspectives on its metabolic function and regulation at the subcellular level. Front. Neuroenergetics 2012, 4, 3. [Google Scholar] [CrossRef]
- Bak, L.K.; Walls, A.B.; Schousboe, A.; Waagepetersen, H.S. Astrocytic glycogen metabolism in the healthy and diseased brain. J. Biol. Chem. 2018, 293, 7108–7116. [Google Scholar] [CrossRef]
- Walls, A.B.; Heimbürger, C.M.; Bouman, S.D.; Schousboe, A.; Waagepetersen, H.S. Robust glycogen shunt activity in astrocytes: Effects of glutamatergic and adrenergic agents. Neuroscience 2009, 158, 284–292. [Google Scholar] [CrossRef]
- Stobart, J.L.; Anderson, C.M. Multifunctional role of astrocytes as gatekeepers of neuronal energy supply. Cell. Neurosci. 2013, 7, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Schousboe, A.; Sickmann, H.M.; Walls, A.B.; Bak, L.K.; Waagepetersen, H.S. Functional importance of the astrocytic glycogen-shunt and glycolysis for maintenance of an intact intra/extracellular glutamate gradient. Neurotox. Res. 2010, 18, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Fink, K.; Velebit, J.; Vardjan, N.; Zorec, R.; Kreft, M. Noradrenaline-induced l-lactate production requires d-glucose entry and transit through the glycogen shunt in single-cultured rat astrocytes. J. Neurosci. Res. 2021, 99, 1084–1098. [Google Scholar] [CrossRef] [PubMed]
- Bheemanapally, K.; Alhamyani, A.R.; Ibrahim, M.M.H.; Briski, K.P. Ventromedial hypothalamic nucleus glycogen phosphorylase regulation of metabolic-sensory neuron AMPK and neurotransmitter protein expression: Role of L-lactate. Amer. J. Physiol. Regul. Integr. Comp. Physiol. 2021, 320, R791–R799. [Google Scholar] [CrossRef] [PubMed]
- Nadeau, O.W.; Fontes, J.D.; Carlson, G.M. The regulation of glycogenolysis in the brain. J. Biol. Chem. 2018, 293, 7099–7107. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.S.; Pedersen, S.; Walls, A.B.; Waagepetersen, H.S.; Bak, L.K. Isoform-selective regulation of glycogen phosphorylase by energy deprivation and phosphorylation in astrocytes. Glia 2014, 63, 154–162. [Google Scholar] [CrossRef]
- Alhamyani, A.; Mahmood, A.S.M.H.; Alshamrani, A.; Ibrahim, M.M.H.; Briski, K.P. Central type II glucocorticoid receptor regulation of ventromedial hypothalamic nucleus glycogen metabolic enzyme and glucoregulatory neurotransmitter marker protein expression in the male rat. J. Endocrinol. Diabetes 2021, 8, 148. [Google Scholar] [CrossRef]
- Vielkind, U.; Walencewicz, A.; Levine, M.; Churchill Bohn, M. Type II glucocorticoid receptors are expressed in oligodendrocytes and astrocytes. J. Neurosci. Res. 1990, 27, 360–373. [Google Scholar] [CrossRef]
- Briski, K.P.; Ibrahim, M.M.H.; Mahmood, A.S.M.H.; Alshamrani, A.A. Norepinephrine regulation of ventromedial hypothalamic nucleus astrocyte glycogen metabolism. Int. J. Mol. Sci. 2021, 22, 759. [Google Scholar] [CrossRef]
- Tamrakar, P.; Shrestha, P.K.; Briski, K.P. Dorsomedial hindbrain catecholamine regulation of hypothalamic astrocyte glycogen metabolic enzyme protein expression: Impact of estradiol. Neuroscience 2015, 292, 34–45. [Google Scholar] [CrossRef]
- Ibrahim, M.M.H.; Alhamami, H.N.; Briski, K.P. Norepinephrine regulation of ventromedial hypothalamic nucleus metabolic transmitter biomarker and astrocyte enzyme and receptor expression: Impact of 5’-AMP-activated protein kinase. Brain Res. 2019, 1711, 48–57. [Google Scholar] [CrossRef]
- Ibrahim, M.M.H.; Bheemanapally, K.; Sylvester, P.W.; Briski, K.P. Norepinephrine regulation of adrenergic receptor expression, 5’ AMP-activated protein kinase activity, and glycogen metabolism and mass in male versus female hypothalamic primary astrocyte cultures. ASN Neuro 2020, 12, 1759091420974134. [Google Scholar] [CrossRef] [PubMed]
- Allaman, I.; Pellerin, L.; Magistretti, P.J. Glucocorticoids modulate neurotransmitter-induced glycogen metabolism in cultured cortical astrocytes. J. Neurochem. 2004, 88, 900–908. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.M.H.; Bheemanapally, K.; Sylvester, P.W.; Briski, K.P. Sex-specific estrogen regulation of hypothalamic astrocyte estrogen receptor expression and glycogen metabolism in rats. Mol. Cell. Endocrinol. 2020, 504, 110703. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.M.H.; Bheemanapally, K.; Sylvester, P.W.; Briski, K.P. Sex differences in glucoprivic regulation of glycogen metabolism in hypothalamic primary astrocyte cultures: Role of estrogen receptor signaling. Mol. Cell. Endocrinol. 2020, 518, 111000. [Google Scholar] [CrossRef]
- Bheemanapally, K.; Ibrahim, M.M.H.; Briski, K.P. Combinatory high-resolution microdissection/ultra-performance liquid chromatographic-mass spectrometry approach for small tissue volume analysis of rat brain glycogen. J. Pharmaceut. Biomed. Anal. 2020, 178, 112884. [Google Scholar] [CrossRef]
- Pacak, K.; Palkovits, M. Stressor specificity of central neuroendocrine responses: Implications for stress-related disorders. Endocr. Rev. 2001, 22, 502–548. [Google Scholar] [CrossRef]
- Ali, M.H.; Napit, P.R.; Mahmood, A.S.M.H.; Bheemanapally, K.; Alhamami, H.N.; Uddin, M.M.; Mandal, K.S.; Ibrahim, M.M.H.; Briski, K.P. Hindbrain estrogen receptor regulation of ventromedial hypothalamic glycogen metabolism and glucoregulatory transmitter expression in the hypoglycemic male rat. Neuroscience 2019, 409, 253–260. [Google Scholar] [CrossRef]
- Napit, P.R.; Ali, M.H.; Shakya, M.; Mandal, S.K.; Bheemanapally, K.; Mahmood, A.S.M.H.; Ibrahim, M.M.H.; Briski, K.P. Hindbrain estrogen receptor regulation of counter-regulatory hormone secretion and ventromedial hypothalamic nucleus glycogen content and glucoregulatory transmitter signaling in hypoglycemic female rats. Neuroscience 2019, 411, 211–221. [Google Scholar] [CrossRef]
- Swanson, R.A.; Choi, D.W. Glial Glycogen Stores Affect Neuronal Survival during Glucose Deprivation in Vitro. J. Cereb. Blood Flow Metab. 1993, 13, 162–169. [Google Scholar] [CrossRef]
- Papadopoulos, M.C.; Koumenis, I.L.; Yuan, T.Y.; Giffard, R.G. Increasing Vulnerability of Astrocytes to Oxidative Injury with Age despite Constant Antioxidant Defenses. Neuroscience 1998, 82, 915–925. [Google Scholar] [CrossRef]
- Ouyang, Y.B.; Xu, L.J.; Sun, Y.J.; Giffard, R.G. Overexpression of inducible heat shock protein 70 and its mutants in astrocytes is associated with maintenance of mitochondrial physiology during glucose deprivation stress. Cell Stress Chaperones 2006, 11, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Fonseca, K.; Massieu, L.; García de la Cadena, S.; Guzmán, C.; Camacho-Arroyo, I. Neuroprotective role of estradiol against neuronal death induced by glucose deprivation in cultured rat hippocampal neurons. Neuroendocrinology 2012, 96, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, A.S.M.H.; Napit, P.R.; Ali, M.H.; Briski, K.P. Estrogen receptor involvement in noradrenergic regulation of ventromedial hypothalamic nucleus glucoregulatory neurotransmitter and stimulus-specific glycogen phosphorylase enzyme isoform expression. ASN Neuro 2020, 12, 1759091420910933. [Google Scholar] [CrossRef]
- Briski, K.P.; Ali, M.H.; Napit, P.R.; Mahmood, A.S.M.H.; Alhamyani, A.R.; Alshamrani, A.A. Sex differences in ventromedial hypothalamic nucleus glucoregulatory transmitter biomarker protein during recurring insulin-induced hypoglycemia. Brain Struct. Funct. 2021, 226, 1053–1065. [Google Scholar] [CrossRef]
- Gilda, J.E.; Gomes, A.V. Western blotting using in-gel protein labeling as a normalization control: Stain-free technology. Methods Mol. Biol. 2015, 1295, 381–391. [Google Scholar] [CrossRef]
- Moritz, C.P. Tubulin or Not Tubulin: Heading Toward Total Protein Staining as Loading Control in Western Blots. Proteomics 2017, 17, 1600189. [Google Scholar] [CrossRef]
- Ibrahim, M.M.H.; Uddin, M.M.; Bheemanapally, K.; Briski, K.P. Sex-dimorphic aromatase regulation of ventromedial hypothalamic nucleus glycogen content in euglycemic and insulin-induced hypoglycemic rats. Neurosci. Lett. 2020, 737, 135284. [Google Scholar] [CrossRef]
- Cai, Y.; Guo, H.; Fan, Z.; Zhang, X.; Wu, D.; Tang, W.; Gu, T.; Wang, S.; Yin, A.; Tao, L.; et al. Glycogenolysis Is Crucial for Astrocytic Glycogen Accumulation and Brain Damage after Reperfusion in Ischemic Stroke. iScience 2020, 23, 101136. [Google Scholar] [CrossRef]
- Zhu, Y.; Fan, Z.; Zhao, Q.; Li, J.; Cai, G.; Wang, R.; Liang, Y.; Lu, N.; Kang, J.; Luo, D.; et al. Brain-Type Glycogen Phosphorylase Is Crucial for Astrocytic Glycogen Accumulation in Chronic Social Defeat Stress-Induced Depression in Mice. Front. Mol. Neurosci. 2022, 14, 819440. [Google Scholar] [CrossRef]
- Mueckler, M.; Thorens, B. The SLC2 (GLUT) family of membrane transporters. Mol. Asp. Med. 2013, 34, 121–138. [Google Scholar] [CrossRef] [PubMed]
- Marty, N.; Dallaporta, M.; Foretz, M.; Emery, M.; Tarussio, D.; Bady, I.; Binnert, C.; Beermann, F.; Thorens, B. Regulation of glucagon secretion by glucose transporter type 2 (GLUT2) and astrocyte-dependent glucose sensors. J. Clin. Investig. 2005, 115, 3543–3553. [Google Scholar] [CrossRef] [PubMed]
- Matschinsky, F.M.; Wilson, D.F. The central role of glucokinase in glucose homeostasis; a perspective 50 years after demonstrating the presence of the enzyme in islets of Langerhans. Front. Physiol. 2019, 10, 148. [Google Scholar] [CrossRef]
- Sze, Y.; Gill, A.C.; Brunton, P.J. Sex-dependent changes in neuroactive steroid concentrations in the rat brain following acute swim stress. J. Neuroendocrinol. 2018, 30, e12644. [Google Scholar] [CrossRef]
- Gibbs, M.E. Role of Glycogenolysis in memory and learning: Regulation by noradrenaline, Serotonin and ATP. Front. Integr. Neurosci. 2016, 9, 70. [Google Scholar] [CrossRef] [PubMed]
- Beard, E.; Lengacher, S.; Dias, S.; Magistretti, P.J.; Finsterwald, C. Astrocytes as key regulators of brain energy metabolism: New therapeutic perspectives. Front. Physiol. 2022, 12, 825816. [Google Scholar] [CrossRef] [PubMed]
- Alhamyani, A.R.; Napit, P.R.; Bheemanapally, K.; Ibrahim, M.M.H.; Sylvester, P.W.; Briski, K.P. Glycogen phosphorylase isoform regulation of elucose and energy sensor expression in male versus female hypothalamic astrocyte primary cultures. Mol. Cell. Endocrinol. 2022, 553, 111698. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).